
Vaccination as a Strategy to Prevent Bluetongue Virus Vertical Transmission


Abstract
:1. Introduction
2. BTV Viral Particle
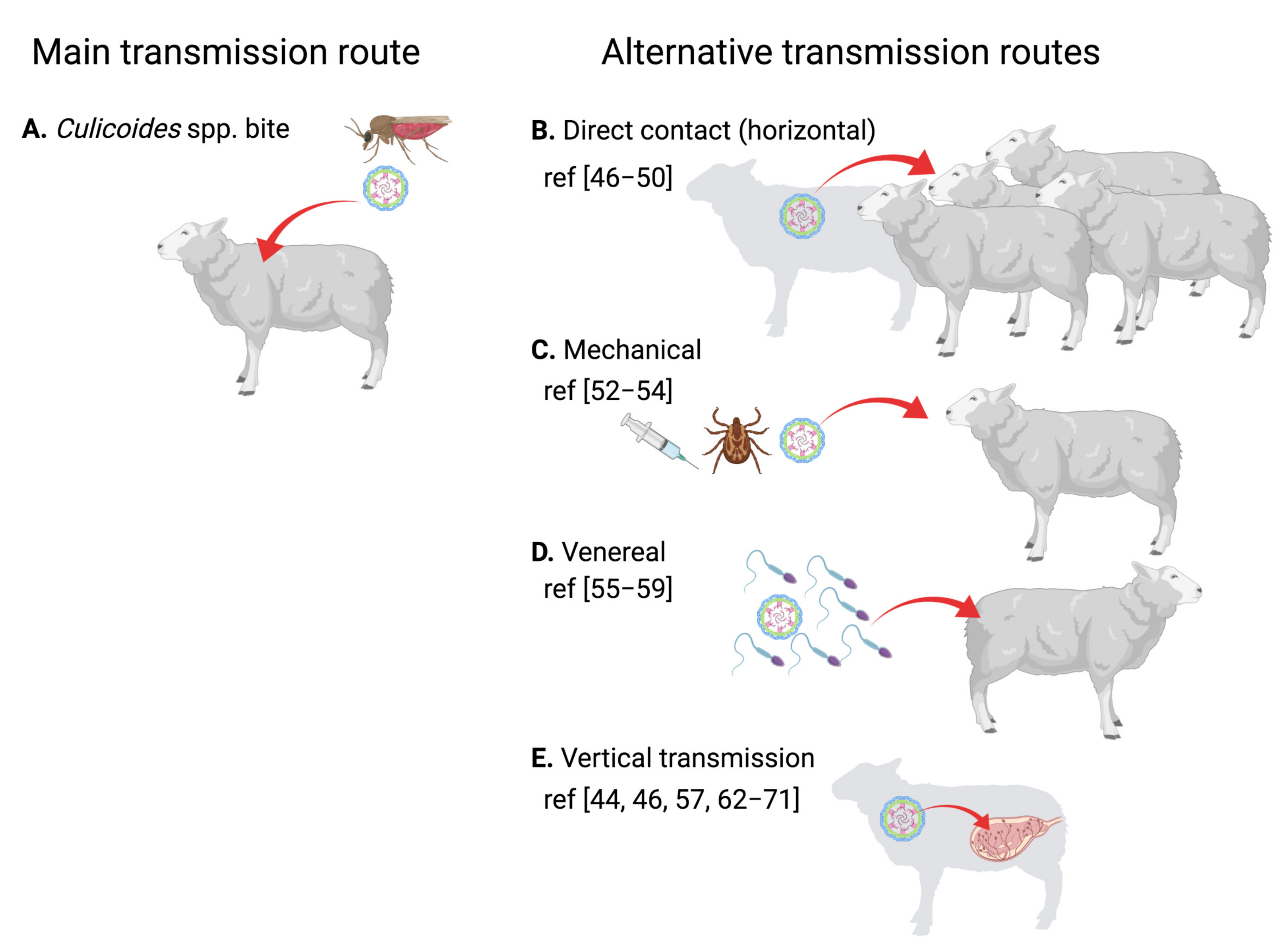
3. BTV Is Mainly an Arbovirus, but It Can Be Transmitted through Other Routes

4. Impact of BTV Vertical Transmission
5. Vaccination as a Strategy to Prevent BTV Vertical Transmission
6. BTV Vaccines: Live Attenuated, Inactivated or Recombinant Vaccines?

{kind=link}
{kind=link}
| Vaccine Type | Protection | Risk of BTV Vertical Transmission | DIVA 1 | |
|---|---|---|---|---|
| Classical | Live attenuated | Yes (serotype specific) | Possible | No |
| Inactivated | Yes (serotype specific) | No | No | |
| Alternative | Recombinant protein | |||
| BTV proteins [110,111,112,113] | Yes | No | Yes | |
| BTV VLP 2 [114] | Yes | No | Yes | |
| Live reverse genetics | ||||
| DISC 3 [120] | Yes | Unlikely; Needs to be tested | Yes 5 | |
| DISA 4 [121,123] | Yes | Needs to be tested | Yes 5 | |
| Viral recombinant vectors | ||||
| Poxvirus [128,129,130] | Yes 6 (potential for multiserotype) | No | Yes | |
| Adenovirus [130,131,132] | Yes 6 (potential for multiserotype) | No | Yes | |
| Rift Valley Fever Virus [133,134] | Yes 6 (bivalent BTV and RVFV) | No | Yes | |
| Herpesvirus [135,136] | Yes 6 (not tested in natural host) | No | Yes | |
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rushton, J.; Lyons, N. Economic impact of Bluetongue: A review of the effects on production. Vet. Ital. 2015, 51, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.; Backx, A.; Meroc, E.; Gerbier, G.; Staubach, C.; Hendrickx, G.; van der Spek, A.; Mintiens, K. Field observations during the bluetongue serotype 8 epidemic in 2006. I. Detection of first outbreaks and clinical signs in sheep and cattle in Belgium, France and the Netherlands. Prev. Vet. Med. 2008, 87, 21–30. [Google Scholar] [CrossRef]
- Katsoulos, P.D.; Giadinis, N.D.; Chaintoutis, S.C.; Dovas, C.I.; Kiossis, E.; Tsousis, G.; Psychas, V.; Vlemmas, I.; Papadopoulos, T.; Papadopoulos, O.; et al. Epidemiological characteristics and clinicopathological features of bluetongue in sheep and cattle, during the 2014 BTV serotype 4 incursion in Greece. Trop. Animal Health Prod. 2016, 48, 469–477. [Google Scholar] [CrossRef]
- Howerth, E.W. Cytokine release and endothelial dysfunction: A perfect storm in orbivirus pathogenesis. Vet Ital. 2015, 51, 275–281. [Google Scholar]
- Williamson, S.; Woodger, N.; Darpel, K. Differential diagnosis of bluetongue in cattle and sheep. In Pract. 2008, 30, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Rojas, J.M.; Rodríguez-Martín, D.; Martín, V.; Sevilla, N. Diagnosing bluetongue virus in domestic ruminants: Current perspectives. Vet. Med. 2019, 10, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Zientara, S.; Sanchez-Vizcaino, J.M. Control of bluetongue in Europe. Vet. Microbiol. 2013, 165, 33–37. [Google Scholar] [CrossRef]
- Alkhamis, M.A.; Aguilar-Vega, C.; Fountain-Jones, N.M.; Lin, K.; Perez, A.M.; Sánchez-Vizcaíno, J.M. Global emergence and evolutionary dynamics of bluetongue virus. Sci. Rep. 2020, 10, 21677. [Google Scholar] [CrossRef]
- Bumbarov, V.; Golender, N.; Jenckel, M.; Wernike, K.; Beer, M.; Khinich, E.; Zalesky, O.; Erster, O. Characterization of bluetongue virus serotype 28. Transbound. Emerg. Dis. 2020, 67, 171–182. [Google Scholar] [CrossRef]
- Sun, E.C.; Huang, L.P.; Xu, Q.Y.; Wang, H.X.; Xue, X.M.; Lu, P.; Li, W.J.; Liu, W.; Bu, Z.G.; Wu, D.L. Emergence of a Novel Bluetongue Virus Serotype, China 2014. Transbound. Emerg. Dis. 2016, 63, 585–589. [Google Scholar] [CrossRef]
- Schulz, C.; Breard, E.; Sailleau, C.; Jenckel, M.; Viarouge, C.; Vitour, D.; Palmarini, M.; Gallois, M.; Hoper, D.; Hoffmann, B.; et al. Bluetongue virus serotype 27: Detection and characterization of two novel variants in Corsica, France. J. Gen. Virol. 2016, 97, 2073–2083. [Google Scholar] [CrossRef]
- Fay, P.C.; Mohd Jaafar, F.; Batten, C.; Attoui, H.; Saunders, K.; Lomonossoff, G.P.; Reid, E.; Horton, D.; Maan, S.; Haig, D.; et al. Serological Cross-Reactions between Expressed VP2 Proteins from Different Bluetongue Virus Serotypes. Viruses 2021, 13, 1455. [Google Scholar] [CrossRef]
- Chaignat, V.; Worwa, G.; Scherrer, N.; Hilbe, M.; Ehrensperger, F.; Batten, C.; Cortyen, M.; Hofmann, M.; Thuer, B. Toggenburg Orbivirus, a new bluetongue virus: Initial detection, first observations in field and experimental infection of goats and sheep. Vet. Microbiol. 2009, 138, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maan, S.; Maan, N.S.; Nomikou, K.; Batten, C.; Antony, F.; Belaganahalli, M.N.; Samy, A.M.; Reda, A.A.; Al-Rashid, S.A.; El Batel, M.; et al. Novel bluetongue virus serotype from Kuwait. Emerg. Infect. Dis. 2011, 17, 886–889. [Google Scholar] [CrossRef]
- Zientara, S.; Sailleau, C.; Viarouge, C.; Höper, D.; Beer, M.; Jenckel, M.; Hoffmann, B.; Romey, A.; Bakkali-Kassimi, L.; Fablet, A.; et al. Novel bluetongue virus in goats, Corsica, France, 2014. Emerg. Infect. Dis. 2014, 20, 2123–2125. [Google Scholar] [CrossRef] [PubMed]
- Ries, C.; Domes, U.; Janowetz, B.; Böttcher, J.; Burkhardt, K.; Miller, T.; Beer, M.; Hoffmann, B. Isolation and Cultivation of a New Isolate of BTV-25 and Presumptive Evidence for a Potential Persistent Infection in Healthy Goats. Viruses 2020, 12, 983. [Google Scholar] [CrossRef]
- OIE. Bluetongue (Infection with Bluetongue Virus). In OIE Terrestrial Manual; OIE: Paris, France, 2014. [Google Scholar]
- Roy, P. Bluetongue virus structure and assembly. Curr. Opin. Virol. 2017, 24, 115–123. [Google Scholar] [CrossRef]
- Stewart, M.; Hardy, A.; Barry, G.; Pinto, R.M.; Caporale, M.; Melzi, E.; Hughes, J.; Taggart, A.; Janowicz, A.; Varela, M.; et al. Characterization of a second open reading frame in genome segment 10 of bluetongue virus. J. Gen. Virol. 2015, 96, 3280–3293. [Google Scholar] [CrossRef]
- Zhang, X.; Patel, A.; Celma, C.C.; Yu, X.; Roy, P.; Zhou, Z.H. Atomic model of a nonenveloped virus reveals pH sensors for a coordinated process of cell entry. Nat. Struct. Mol. Biol. 2016, 23, 74–80. [Google Scholar] [CrossRef]
- Wu, W.; Celma, C.C.; Kerviel, A.; Roy, P. Mapping the pH Sensors Critical for Host Cell Entry by a Complex Nonenveloped Virus. J. Virol. 2019, 93, e01897–e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Shivakoti, S.; Ding, K.; Cui, Y.; Roy, P.; Zhou, Z.H. In situ structures of RNA-dependent RNA polymerase inside bluetongue virus before and after uncoating. Proc. Natl. Acad. Sci. USA 2019, 116, 16535–16540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nason, E.L.; Rothagel, R.; Mukherjee, S.K.; Kar, A.K.; Forzan, M.; Prasad, B.V.V.; Roy, P. Interactions between the inner and outer capsids of bluetongue virus. J. Virol. 2004, 78, 8059–8067. [Google Scholar] [CrossRef] [Green Version]
- Sung, P.Y.; Vaughan, R.; Rahman, S.K.; Yi, G.; Kerviel, A.; Kao, C.C.; Roy, P. The Interaction of Bluetongue Virus VP6 and Genomic RNA Is Essential for Genome Packaging. J. Virol. 2019, 93, e02023–e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, M.; Celma, C.P.; Roy, P. Bluetongue virus non-structural protein 1 is a positive regulator of viral protein synthesis. Virol. J. 2012, 9, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerviel, A.; Ge, P.; Lai, M.; Jih, J.; Boyce, M.; Zhang, X.; Zhou, Z.H.; Roy, P. Atomic structure of the translation regulatory protein NS1 of bluetongue virus. Nat. Microbiol. 2019, 4, 837–845. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.K.; Kerviel, A.; Mohl, B.-P.; He, Y.; Zhou, Z.H.; Roy, P. A Calcium Sensor Discovered in Bluetongue Virus Nonstructural Protein 2 Is Critical for Virus Replication. J. Virol. 2020, 94, e01099–e20. [Google Scholar] [CrossRef]
- Kar, A.K.; Bhattacharya, B.; Roy, P. Bluetongue virus RNA binding protein NS2 is a modulator of viral replication and assembly. BMC Mol. Biol. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirblich, C.; Bhattacharya, B.; Roy, P. Nonstructural protein 3 of bluetongue virus assists virus release by recruiting ESCRT-I protein Tsg101. J. Virol. 2006, 80, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Celma, C.C.; Roy, P. A viral nonstructural protein regulates bluetongue virus trafficking and release. J. Virol. 2009, 83, 6806–6816. [Google Scholar] [CrossRef] [Green Version]
- Labadie, T.; Jegouic, S.; Roy, P. Bluetongue Virus Nonstructural Protein 3 Orchestrates Virus Maturation and Drives Non-Lytic Egress via Two Polybasic Motifs. Viruses 2019, 11, 1107. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Harty, R.N. The NS3 protein of bluetongue virus exhibits viroporin-like properties. J. Biol. Chem. 2004, 279, 43092–43097. [Google Scholar] [CrossRef] [Green Version]
- Beaton, A.R.; Rodriguez, J.; Reddy, Y.K.; Roy, P. The membrane trafficking protein calpactin forms a complex with bluetongue virus protein NS3 and mediates virus release. Proc. Natl. Acad. Sci. USA 2002, 99, 13154–13159. [Google Scholar] [CrossRef] [Green Version]
- Rojas, J.M.; Avia, M.; Martín, V.; Sevilla, N. Inhibition of the IFN Response by Bluetongue Virus: The Story So Far. Front. Microbiol. 2021, 12, 692069. [Google Scholar] [CrossRef]
- Pourcelot, M.; Amaral Moraes, R.; Fablet, A.; Bréard, E.; Sailleau, C.; Viarouge, C.; Postic, L.; Zientara, S.; Caignard, G.; Vitour, D. The VP3 Protein of Bluetongue Virus Associates with the MAVS Complex and Interferes with the RIG-I-Signaling Pathway. Viruses 2021, 13, 230. [Google Scholar] [CrossRef]
- Avia, M.; Rojas, J.M.; Miorin, L.; Pascual, E.; Van Rijn, P.A.; Martín, V.; García-Sastre, A.; Sevilla, N. Virus-induced autophagic degradation of STAT2 as a mechanism for interferon signaling blockade. EMBO Rep. 2019, 20, e48766. [Google Scholar] [CrossRef] [PubMed]
- Ratinier, M.; Shaw, A.E.; Barry, G.; Gu, Q.; Di Gialleonardo, L.; Janowicz, A.; Varela, M.; Randall, R.E.; Caporale, M.; Palmarini, M. Bluetongue Virus NS4 Protein Is an Interferon Antagonist and a Determinant of Virus Virulence. J. Virol. 2016, 90, 5427–5439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauveau, E.; Doceul, V.; Lara, E.; Breard, E.; Sailleau, C.; Vidalain, P.O.; Meurs, E.F.; Dabo, S.; Schwartz-Cornil, I.; Zientara, S.; et al. NS3 of bluetongue virus interferes with the induction of type I interferon. J. Virol. 2013, 87, 8241–8246. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lu, D.; Yang, H.; Li, Z.; Zhu, P.; Xie, J.; Liao, D.; Zheng, Y.; Li, H. Bluetongue virus non-structural protein 3 (NS3) and NS4 coordinatively antagonize type Ⅰ interferon signaling by targeting STAT1. Vet. Microbiol. 2021, 254, 108986. [Google Scholar] [CrossRef] [PubMed]
- Pourcelot, M.; Zemirli, N.; Da Costa, L.S.; Loyant, R.; Garcin, D.; Vitour, D.; Munitic, I.; Vazquez, A.; Arnoult, D. The Golgi apparatus acts as a platform for TBK1 activation after viral RNA sensing. BMC Biol. 2016, 14, 69. [Google Scholar] [CrossRef] [PubMed]
- Baylis, M.; O’Connell, L.; Mellor, P.S. Rates of bluetongue virus transmission between Culicoides sonorensis and sheep. Med. Vet. Entomol. 2008, 22, 228–237. [Google Scholar] [CrossRef]
- Alexander, K.A.; MacLachlan, N.J.; Kat, P.W.; House, C.; O’Brien, S.J.; Lerche, N.W.; Sawyer, M.; Frank, L.G.; Holekamp, K.; Smale, L.; et al. Evidence of natural bluetongue virus infection among African carnivores. Am. J. Trop. Med. Hyg. 1994, 51, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Jauniaux, T.P.; De Clercq, K.E.; Cassart, D.E.; Kennedy, S.; Vandenbussche, F.E.; Vandemeulebroucke, E.L.; Vanbinst, T.M.; Verheyden, B.I.; Goris, N.E.; Coignoul, F.L. Bluetongue in Eurasian lynx. Emerg. Infect. Dis. 2008, 14, 1496–1498. [Google Scholar] [CrossRef]
- Rasmussen, L.D.; Savini, G.; Lorusso, A.; Bellacicco, A.; Palmarini, M.; Caporale, M.; Rasmussen, T.B.; Belsham, G.J.; Bøtner, A. Transplacental transmission of field and rescued strains of BTV-2 and BTV-8 in experimentally infected sheep. Vet. Res. 2013, 44, 75. [Google Scholar] [CrossRef] [Green Version]
- van der Sluijs, M.; Timmermans, M.; Moulin, V.; Noordegraaf, C.V.; Vrijenhoek, M.; Debyser, I.; de Smit, A.J.; Moormann, R. Transplacental transmission of Bluetongue virus serotype 8 in ewes in early and mid gestation. Vet. Microbiol. 2011, 149, 113–125. [Google Scholar] [CrossRef]
- van der Sluijs, M.T.; Schroer-Joosten, D.P.; Fid-Fourkour, A.; Vrijenhoek, M.P.; Debyser, I.; Moulin, V.; Moormann, R.J.; de Smit, A.J. Transplacental transmission of Bluetongue virus serotype 1 and serotype 8 in sheep: Virological and pathological findings. PLoS ONE 2013, 8, e81429. [Google Scholar] [CrossRef]
- Mayo, C.E.; Crossley, B.M.; Hietala, S.K.; Gardner, I.A.; Breitmeyer, R.E.; Maclachlan, N.J. Colostral transmission of bluetongue virus nucleic acid among newborn dairy calves in California. Transbound. Emerg. Dis. 2010, 57, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Menzies, F.D.; McCullough, S.J.; McKeown, I.M.; Forster, J.L.; Jess, S.; Batten, C.; Murchie, A.K.; Gloster, J.; Fallows, J.G.; Pelgrim, W.; et al. Evidence for transplacental and contact transmission of bluetongue virus in cattle. Vet. Rec. 2008, 163, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Batten, C.A.; Henstock, M.R.; Steedman, H.M.; Waddington, S.; Edwards, L.; Oura, C.A. Bluetongue virus serotype 26: Infection kinetics, pathogenesis and possible contact transmission in goats. Vet. Microbiol. 2013, 162, 62–67. [Google Scholar] [CrossRef]
- Bréard, E.; Schulz, C.; Sailleau, C.; Bernelin-Cottet, C.; Viarouge, C.; Vitour, D.; Guillaume, B.; Caignard, G.; Gorlier, A.; Attoui, H.; et al. Bluetongue virus serotype 27: Experimental infection of goats, sheep and cattle with three BTV-27 variants reveal atypical characteristics and likely direct contact transmission BTV-27 between goats. Transbound. Emerg. Dis. 2018, 65, e251–e263. [Google Scholar] [CrossRef]
- Barratt-Boyes, S.M.; MacLachlan, N.J. Dynamics of viral spread in bluetongue virus infected calves. Vet. Microbiol. 1994, 40, 361–371. [Google Scholar] [CrossRef]
- Darpel, K.E.; Barber, J.; Hope, A.; Wilson, A.J.; Gubbins, S.; Henstock, M.; Frost, L.; Batten, C.; Veronesi, E.; Moffat, K.; et al. Using shared needles for subcutaneous inoculation can transmit bluetongue virus mechanically between ruminant hosts. Sci. Rep. 2016, 6, 20627. [Google Scholar] [CrossRef] [Green Version]
- Stott, J.L.; Osburn, B.I.; Alexander, L. Ornithodoros coriaceus (pajaroello tick) as a vector of bluetongue virus. Am. J. Vet. Res. 1985, 46, 1197–1199. [Google Scholar]
- Bouwknegt, C.; van Rijn, P.A.; Schipper, J.J.; Hölzel, D.; Boonstra, J.; Nijhof, A.M.; van Rooij, E.M.; Jongejan, F. Potential role of ticks as vectors of bluetongue virus. Exp. Appl. Acarol. 2010, 52, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Vanbinst, T.; Vandenbussche, F.; Dernelle, E.; De Clercq, K. A duplex real-time RT-PCR for the detection of bluetongue virus in bovine semen. J. Virol. Methods 2010, 169, 162–168. [Google Scholar] [CrossRef]
- Leemans, J.; Raes, M.; Vanbinst, T.; De Clercq, K.; Saegerman, C.; Kirschvink, N. Viral RNA load in semen from bluetongue serotype 8-infected rams: Relationship with sperm quality. Vet. J. 2012, 192, 304–310. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, K.; Vandaele, L.; Vanbinst, T.; Riou, M.; Deblauwe, I.; Wesselingh, W.; Pinard, A.; Van Eetvelde, M.; Boulesteix, O.; Leemans, B.; et al. Transmission of Bluetongue Virus Serotype 8 by Artificial Insemination with Frozen-Thawed Semen from Naturally Infected Bulls. Viruses 2021, 13, 652. [Google Scholar] [CrossRef]
- Bowen, R.A.; Howard, T.H.; Pickett, B.W. Seminal shedding of bluetongue virus in experimentally infected bulls. Prog. Clin. Biol. Res. 1985, 178, 91–96. [Google Scholar]
- Thomas, F.C.; Singh, E.L.; Hare, W.C. Embryo transfer as a means of controlling viral infections. VI. Bluetongue virus-free calves from infectious semen. Theriogenology 1985, 24, 345–350. [Google Scholar] [CrossRef]
- Pascall, D.J.; Nomikou, K.; Bréard, E.; Zientara, S.; Filipe, A.D.S.; Hoffmann, B.; Jacquot, M.; Singer, J.B.; De Clercq, K.; Bøtner, A.; et al. “Frozen evolution” of an RNA virus suggests accidental release as a potential cause of arbovirus re-emergence. PLoS Biol. 2020, 18, e3000673. [Google Scholar] [CrossRef]
- Schultz, G.; Delay, P.D. Losses in newborn lambs associated with bluetongue vaccination of pregnancy ewes. J. Am. Vet. Med. Assoc. 1955, 127, 224–226. [Google Scholar]
- Gibbs, E.P.; Lawman, M.J.; Herniman, K.O.A. Preliminary observations on transplacental infection of bluetongue virus in sheep-a possible overwintering mechanism. Res. Vet. Sci. 1979, 27, 118–120. [Google Scholar] [CrossRef]
- van der Sluijs, M.T.; de Smit, A.J.; Moormann, R.J. Vector independent transmission of the vector-borne bluetongue virus. Crit. Rev. Microbiol. 2016, 42, 57–64. [Google Scholar] [CrossRef]
- Richardson, C.; Taylor, W.P.; Terlecki, S.; Gibbs, E.P. Observations on transplacental infection with bluetongue virus in sheep. Am. J. Vet. Res. 1985, 46, 1912–1922. [Google Scholar]
- Savini, G.; Lorusso, A.; Paladini, C.; Migliaccio, P.; Di Gennaro, A.; Di Provvido, A.; Scacchia, M.; Monaco, F. Bluetongue serotype 2 and 9 modified live vaccine viruses as causative agents of abortion in livestock: A retrospective analysis in Italy. Transbound. Emerg. Dis. 2014, 61, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Stott, J.L.; Lauerman, L.H.; Luedke, A.J. Bluetongue virus in pregnant elk and their calves. Am. J. Vet. Res. 1982, 43, 423–428. [Google Scholar] [PubMed]
- De Clercq, K.; De Leeuw, I.; Verheyden, B.; Vandemeulebroucke, E.; Vanbinst, T.; Herr, C.; Méroc, E.; Bertels, G.; Steurbaut, N.; Miry, C.; et al. Transplacental infection and apparently immunotolerance induced by a wild-type bluetongue virus serotype 8 natural infection. Transbound. Emerg. Dis. 2008, 55, 352–359. [Google Scholar] [CrossRef]
- Darpel, K.E.; Batten, C.A.; Veronesi, E.; Williamson, S.; Anderson, P.; Dennison, M.; Clifford, S.; Smith, C.; Philips, L.; Bidewell, C.; et al. Transplacental transmission of bluetongue virus 8 in cattle, UK. Emerg. Infect. Dis. 2009, 15, 2025–2028. [Google Scholar] [CrossRef] [PubMed]
- Vercauteren, G.; Miry, C.; Vandenbussche, F.; Ducatelle, R.; Van der Heyden, S.; Vandemeulebroucke, E.; De Leeuw, I.; Deprez, P.; Chiers, K.; De Clercq, K. Bluetongue virus serotype 8-associated congenital hydranencephaly in calves. Transbound. Emerg. Dis. 2008, 55, 293–298. [Google Scholar] [CrossRef]
- Santman-Berends, I.M.; van Wuijckhuise, L.; Vellema, P.; van Rijn, P.A. Vertical transmission of bluetongue virus serotype 8 virus in Dutch dairy herds in 2007. Vet. Microbiol. 2010, 141, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Santman-Berends, I.M.G.A.; Hage, J.J.; van Rijn, P.A.; Stegeman, J.A.; van Schaik, G. Bluetongue virus serotype 8 (BTV-8) infection reduces fertility of Dutch dairy cattle and is vertically transmitted to offspring. Theriogenology 2010, 74, 1377–1384. [Google Scholar] [CrossRef]
- Akita, G.Y.; Ianconescu, M.; MacLachlan, N.J.; Osburn, B.I. Bluetongue disease in dogs associated with contaminated vaccine. Vet. Rec. 1994, 134, 283–284. [Google Scholar] [CrossRef]
- Brown, C.C.; Rhyan, J.C.; Grubman, M.J.; Wilbur, L.A. Distribution of bluetongue virus in tissues of experimentally infected pregnant dogs as determined by in situ hybridization. Vet. Pathol. 1996, 33, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Dubovi, E.J.; Hawkins, M.; Griffin, R.A., Jr.; Johnson, D.J.; Ostlund, E.N. Isolation of Bluetongue virus from canine abortions. J. Vet. Diagn. Investig. 2013, 25, 490–492. [Google Scholar] [CrossRef]
- Evermann, J.F. Letter to the Editor, regarding Bluetongue virus and canine abortions. J. Vet. Diagn. Investig. 2013, 25, 670. [Google Scholar] [CrossRef]
- Courtejoie, N.; Bournez, L.; Zanella, G.; Durand, B. Quantifying bluetongue vertical transmission in French cattle from surveillance data. Vet. Res. 2019, 50, 34. [Google Scholar] [CrossRef] [Green Version]
- Barnard, B.J.; Pienaar, J.G. Bluetongue virus as a cause of hydranencephaly in cattle. Onderstepoort J. Vet. Res. 1976, 43, 155–157. [Google Scholar]
- MacLachlan, N.J.; Osburn, B.I. Bluetongue virus-induced hydranencephaly in cattle. Vet. Pathol. 1983, 20, 563–573. [Google Scholar] [CrossRef] [Green Version]
- MacLachlan, N.J.; Osburn, B.I.; Ghalib, H.W.; Stott, J.L. Bluetongue virus-induced encephalopathy in fetal cattle. Vet. Pathol. 1985, 22, 415–417. [Google Scholar] [CrossRef]
- Carr, M.A.; De Mattos, C.C.; De Mattos, C.A.; Osburn, B.I. Association of bluetongue virus gene segment 5 with neuroinvasiveness. J. Virol. 1994, 68, 1255–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maclachlan, N.J.; Osburn, B.I. Teratogenic bluetongue and related orbivirus infections in pregnant ruminant livestock: Timing and pathogen genetics are critical. Curr. Opin. Virol. 2017, 27, 31–35. [Google Scholar] [CrossRef]
- Silverstein, A.M.; Uhr, J.W.; Kraner, K.L.; Lukes, R.J. Fetal response to antigenic stimulus. II. Antibody production by the fetal lamb. J. Exp. Med. 1963, 117, 799–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, R.D.; Dunne, H.W.; Heist, C.E. Ontogeny of the bovine immune response. Infect. Immun. 1973, 7, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Waldvogel, A.S.; Anderson, G.A.; Phillips, D.L.; Osburn, B.I. Association of virulent and avirulent strains of bluetongue virus serotype 11 with premature births of late-term bovine fetuses. J. Comp. Pathol. 1992, 106, 333–340. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Hawkes, R.A. A comparison of laboratory and ‘wild’ strains of bluetongue virus-is there any difference and does it matter? Vet. Ital. 2004, 40, 448–455. [Google Scholar]
- Osburn, B.I.; Johnson, R.T.; Silverstein, A.M.; Prendergast, R.A.; Jochim, M.M.; Levy, S.E. Experimental viral-induced congenital encephalopathies. II. The pathogenesis of bluetongue vaccine virus infection in fetal lambs. Lab. Investig. J. Tech. Methods Pathol. 1971, 25, 206–210. [Google Scholar]
- Osburn, B.I.; Silverstein, A.M.; Prendergast, R.A.; Johnson, R.T.; Parshall, C.J., Jr. Experimental viral-induced congenital encephalopathies. I. Pathology of hydranencephaly and porencephaly caused by bluetongue vaccine virus. Lab. Investig. J. Tech. Methods Pathol. 1971, 25, 197–205. [Google Scholar]
- Nusinovici, S.; Madouasse, A.; Fourichon, C. Quantification of the increase in the frequency of early calving associated with late exposure to bluetongue virus serotype 8 in dairy cows: Implications for syndromic surveillance. Vet. Res. 2016, 47, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marceau, A.; Madouasse, A.; Lehébel, A.; van Schaik, G.; Veldhuis, A.; Van der Stede, Y.; Fourichon, C. Can routinely recorded reproductive events be used as indicators of disease emergence in dairy cattle? An evaluation of 5 indicators during the emergence of bluetongue virus in France in 2007 and 2008. J. Dairy Sci. 2014, 97, 6135–6150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prendergast, A.J.; Klenerman, P.; Goulder, P.J. The impact of differential antiviral immunity in children and adults. Nat. Rev. Immunol. 2012, 12, 636–648. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, K.R.; DeMaula, C.D.; Mullens, B.A.; MacLachlan, N.J. Duration of viraemia infectious to Culicoides sonorensis in bluetongue virus-infected cattle and sheep. Vet. Microbiol. 2002, 88, 115–125. [Google Scholar] [CrossRef]
- Melzi, E.; Caporale, M.; Rocchi, M.; Martin, V.; Gamino, V.; di Provvido, A.; Marruchella, G.; Entrican, G.; Sevilla, N.; Palmarini, M. Follicular dendritic cell disruption as a novel mechanism of virus-induced immunosuppression. Proc. Natl. Acad. Sci. USA 2016, 113, E6238–E6247. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martín, D.; Louloudes-Lázaro, A.; Avia, M.; Martín, V.; Rojas, J.M.; Sevilla, N. The Interplay between Bluetongue Virus Infections and Adaptive Immunity. Viruses 2021, 13, 1511. [Google Scholar] [CrossRef]
- Ellis, J.A.; Luedke, A.J.; Davis, W.C.; Wechsler, S.J.; Mecham, J.O.; Pratt, D.L.; Elliott, J.D. T Lymphocyte Subset Alterations Following Bluetongue Virus Infection in Sheep and Cattle. Vet. Immunol. Immunopathol. 1990, 24, 49–67. [Google Scholar] [CrossRef]
- Nomikou, K.; Hughes, J.; Wash, R.; Kellam, P.; Breard, E.; Zientara, S.; Palmarini, M.; Biek, R.; Mertens, P. Widespread Reassortment Shapes the Evolution and Epidemiology of Bluetongue Virus following European Invasion. PLoS Pathog. 2015, 11, e1005056. [Google Scholar] [CrossRef]
- Maan, S.; Maan, N.S.; Ross-smith, N.; Batten, C.A.; Shaw, A.E.; Anthony, S.J.; Samuel, A.R.; Darpel, K.E.; Veronesi, E.; Oura, C.A.L.; et al. Sequence analysis of bluetongue virus serotype 8 from the Netherlands 2006 and comparison to other European strains. Virology 2008, 377, 308–318. [Google Scholar] [CrossRef] [Green Version]
- MacLennan, C.A.; Saul, A. Vaccines against poverty. Proc. Natl. Acad. Sci. USA 2014, 111, 12307–12312. [Google Scholar] [CrossRef] [Green Version]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef]
- Evans-Gilbert, T. Vertically transmitted chikungunya, Zika and dengue virus infections: The pathogenesis from mother to fetus and the implications of co-infections and vaccine development. Int. J. Pediatrics Adolesc. Med. 2020, 7, 107–111. [Google Scholar] [CrossRef]
- World Health Organization. Dengue vaccine: WHO position paper, September 2018–Recommendations. Vaccine 2019, 37, 4848–4849. [Google Scholar] [CrossRef]
- Shan, C.; Xie, X.; Luo, H.; Muruato, A.E.; Liu, Y.; Wakamiya, M.; La, J.H.; Chung, J.M.; Weaver, S.C.; Wang, T.; et al. Maternal vaccination and protective immunity against Zika virus vertical transmission. Nat. Commun. 2019, 10, 5677. [Google Scholar] [CrossRef] [Green Version]
- Barrett, A.D.T. Current status of Zika vaccine development: Zika vaccines advance into clinical evaluation. NPJ Vaccines 2018, 3, 24. [Google Scholar] [CrossRef]
- Martinelle, L.; Dal Pozzo, F.; Thiry, E.; De Clercq, K.; Saegerman, C. Reliable and Standardized Animal Models to Study the Pathogenesis of Bluetongue and Schmallenberg Viruses in Ruminant Natural Host Species with Special Emphasis on Placental Crossing. Viruses 2019, 11, 753. [Google Scholar] [CrossRef] [Green Version]
- Saminathan, M.; Singh, K.P.; Vineetha, S.; Maity, M.; Biswas, S.K.; Manjunathareddy, G.B.; Chauhan, H.C.; Milton, A.A.P.; Ramakrishnan, M.A.; Maan, S.; et al. Virological, immunological and pathological findings of transplacentally transmitted bluetongue virus serotype 1 in IFNAR1-blocked mice during early and mid gestation. Sci. Rep. 2020, 10, 2164. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Pinilla, E.; Rodriguez-Calvo, T.; Anguita, J.; Sevilla, N.; Ortego, J. Establishment of a bluetongue virus infection model in mice that are deficient in the alpha/beta interferon receptor. PLoS ONE 2009, 4, e5171. [Google Scholar] [CrossRef] [Green Version]
- Martinelle, L.; Dal Pozzo, F.; Sarradin, P.; De Leeuw, I.; De Clercq, K.; Thys, C.; Thiry, E.; Saegerman, C. Pulmonary artery haemorrhage in newborn calves following bluetongue virus serotype 8 experimental infections of pregnant heifers. Vet. Microbiol. 2013, 167, 250–259. [Google Scholar] [CrossRef]
- Miyazawa, T.; Yoshikawa, R.; Golder, M.; Okada, M.; Stewart, H.; Palmarini, M. Isolation of an infectious endogenous retrovirus in a proportion of live attenuated vaccines for pets. J. Virol. 2010, 84, 3690–3694. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Li, Y.; Zhang, Y.; Zhang, Z.; Meng, F.; Cui, Z.; Chang, S.; Zhao, P. Newcastle disease virus-attenuated vaccine LaSota played a key role in the pathogenicity of contaminated exogenous virus. Vet. Res. 2018, 49, 80. [Google Scholar] [CrossRef] [Green Version]
- van Rijn, P.A. Prospects of Next-Generation Vaccines for Bluetongue. Front. Vet. Sci. 2019, 6, 407. [Google Scholar] [CrossRef] [Green Version]
- Urakawa, T.; French, T.J.; Adachi, Y.; Fukusho, A.; LeBlois, H.; Flamand, M.; Mertens, P.; Roy, P. Synthesis of recombinant baculoviruses expressing the outer capsid protein VP2 of five BTV serotypes and the induction of neutralizing antibodies to homologous and heterologous BTV serotypes. Virus Res. 1994, 31, 149–161. [Google Scholar] [CrossRef]
- Inumaru, S.; Roy, P. Production and characterization of the neutralization antigen VP2 of bluetongue virus serotype 10 using a baculovirus expression vector. Virology 1987, 157, 472–479. [Google Scholar] [CrossRef]
- Fay, P.C.; Attoui, H.; Batten, C.; Mohd Jaafar, F.; Lomonossoff, G.P.; Daly, J.M.; Mertens, P.P.C. Bluetongue virus outer-capsid protein VP2 expressed in Nicotiana benthamiana raises neutralising antibodies and a protective immune response in IFNAR −/− mice. Vaccine: X 2019, 2, 100026. [Google Scholar] [CrossRef] [PubMed]
- Athmaram, T.N.; Bali, G.; Kahng, G.G.; Dwarakanath, S. Heterologous expression of Bluetongue VP2 viral protein fragment in Pichia pastoris. Virus Genes 2007, 35, 265–271. [Google Scholar] [CrossRef] [PubMed]
- French, T.J.; Marshall, J.J.; Roy, P. Assembly of double-shelled, viruslike particles of bluetongue virus by the simultaneous expression of four structural proteins. J. Virol. 1990, 64, 5695–5700. [Google Scholar] [CrossRef] [Green Version]
- Thuenemann, E.C.; Meyers, A.E.; Verwey, J.; Rybicki, E.P.; Lomonossoff, G.P. A method for rapid production of heteromultimeric protein complexes in plants: Assembly of protective bluetongue virus-like particles. Plant. Biotechnol. J. 2013, 11, 839–846. [Google Scholar] [CrossRef]
- Roy, P. Genetically engineered structure-based vaccine for bluetongue disease. Vet. Ital. 2004, 40, 594–600. [Google Scholar] [PubMed]
- Anderson, J.; Hägglund, S.; Bréard, E.; Riou, M.; Zohari, S.; Comtet, L.; Olofson, A.S.; Gélineau, R.; Martin, G.; Elvander, M.; et al. Strong protection induced by an experimental DIVA subunit vaccine against bluetongue virus serotype 8 in cattle. Vaccine 2014, 32, 6614–6621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, M.; Celma, C.C.P.; Roy, P. Development of Reverse Genetics Systems for Bluetongue Virus: Recovery of Infectious Virus from Synthetic RNA Transcripts. J. Virol. 2008, 82, 8339–8348. [Google Scholar] [CrossRef] [Green Version]
- van Gennip, R.G.; van de Water, S.G.; Maris-Veldhuis, M.; van Rijn, P.A. Bluetongue viruses based on modified-live vaccine serotype 6 with exchanged outer shell proteins confer full protection in sheep against virulent BTV8. PLoS ONE 2012, 7, e44619. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, E.; Celma, C.C.; Boyce, M.; Viarouge, C.; Sailleau, C.; Dubois, E.; Bréard, E.; Thiéry, R.; Zientara, S.; Roy, P. Generation of replication-defective virus-based vaccines that confer full protection in sheep against virulent bluetongue virus challenge. J. Virol. 2011, 85, 10213–10221. [Google Scholar] [CrossRef] [Green Version]
- Feenstra, F.; van Gennip, R.G.P.; Maris-Veldhuis, M.; Verheij, E.; van Rijn, P.A. Bluetongue virus without NS3/NS3a expression is not virulent and protects against virulent bluetongue virus challenge. J. Gen. Virol. 2014, 95, 2019–2029. [Google Scholar] [CrossRef]
- van Rijn, P.A.; Maris-Veldhuis, M.A.; van Gennip, R.G.P. The Bluetongue Disabled Infectious Single Animal (DISA) Vaccine Platform Based on Deletion NS3/NS3a Protein Is Safe and Protective in Cattle and Enables DIVA. Viruses 2021, 13, 857. [Google Scholar] [CrossRef]
- van Rijn, P.A.; Daus, F.J.; Maris-Veldhuis, M.A.; Feenstra, F.; van Gennip, R.G.P. Bluetongue Disabled Infectious Single Animal (DISA) vaccine: Studies on the optimal route and dose in sheep. Vaccine. 2017, 35, 231–237. [Google Scholar] [CrossRef]
- Feenstra, F.; Drolet, B.S.; Boonstra, J.; van Rijn, P.A. Non-structural protein NS3/NS3a is required for propagation of bluetongue virus in Culicoides sonorensis. Parasites Vectors 2015, 8, 476. [Google Scholar] [CrossRef] [Green Version]
- Rojas, J.M.; Sevilla, N.; Martín, V. A New Look at Vaccine Strategies Against PPRV Focused on Adenoviral Candidates. Front. Vet. Sci. 2021, 8, 1005. [Google Scholar] [CrossRef]
- García-Arriaza, J.; Esteban, M. Enhancing poxvirus vectors vaccine immunogenicity. Hum. Vaccines Immunother. 2014, 10, 2235–2244. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Calvo-Pinilla, E.; Moreno, S.; Nogales, A.; Ortego, J.; Marín-López, A. Viral Vector Vaccines against Bluetongue Virus. Microorganisms 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Pinilla, E.; Navasa, N.; Anguita, J.; Ortego, J. Multiserotype protection elicited by a combinatorial prime-boost vaccination strategy against bluetongue virus. PLoS ONE 2012, 7, e34735. [Google Scholar] [CrossRef]
- Lobato, Z.I.P.; Coupar, B.E.H.; Gray, C.P.; Lunt, R.; Andrew, M.E. Antibody responses and protective immunity to recombinant vaccinia virus-expressed bluetongue virus antigens. Vet. Immunol. Immunopathol. 1997, 59, 293–309. [Google Scholar] [CrossRef]
- Utrilla-Trigo, S.; Jiménez-Cabello, L.; Alonso-Ravelo, R.; Calvo-Pinilla, E.; Marín-López, A.; Moreno, S.; Lorenzo, G.; Benavides, J.; Gilbert, S.; Nogales, A.; et al. Heterologous Combination of ChAdOx1 and MVA Vectors Expressing Protein NS1 as Vaccination Strategy to Induce Durable and Cross-Protective CD8+ T Cell Immunity to Bluetongue Virus. Vaccines 2020, 8, 346. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Pascual, E.; Avia, M.; Pena, L.; Valcarcel, F.; Sevilla, N. Protective Efficacy in Sheep of Adenovirus-Vectored Vaccines against Bluetongue Virus Is Associated with Specific T Cell Responses. PLoS ONE 2015, 10, e0143273. [Google Scholar] [CrossRef]
- Rojas, J.M.; Barba-Moreno, D.; Avia, M.; Sevilla, N.; Martín, V. Vaccination With Recombinant Adenoviruses Expressing the Bluetongue Virus Subunits VP7 and VP2 Provides Protection Against Heterologous Virus Challenge. Front. Vet. Sci. 2021, 8, 158. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Pinilla, E.; Marín-López, A.; Moreno, S.; Lorenzo, G.; Utrilla-Trigo, S.; Jiménez-Cabello, L.; Benavides, J.; Nogales, A.; Blasco, R.; Brun, A.; et al. A protective bivalent vaccine against Rift Valley fever and bluetongue. NPJ Vaccines 2020, 5, 70. [Google Scholar] [CrossRef]
- Moreno, S.; Calvo-Pinilla, E.; Devignot, S.; Weber, F.; Ortego, J.; Brun, A. Recombinant Rift Valley fever viruses encoding bluetongue virus (BTV) antigens: Immunity and efficacy studies upon a BTV-4 challenge. PLoS Negl. Trop. Dis. 2020, 14, e0008942. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V.; Capocefalo, A.; Calvo-Pinilla, E.; Redaelli, M.; Mucignat-Caretta, C.; Mertens, P.; Ortego, J.; Donofrio, G. Immunization of knock-out α/β interferon receptor mice against lethal bluetongue infection with a BoHV-4-based vector expressing BTV-8 VP2 antigen. Vaccine 2011, 29, 3074–3082. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Eschbaumer, M.; Said, A.; Hoffmann, B.; Beer, M.; Osterrieder, N. An equine herpesvirus type 1 (EHV-1) expressing VP2 and VP5 of serotype 8 bluetongue virus (BTV-8) induces protection in a murine infection model. PLoS ONE 2012, 7, e34425. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas, J.M.; Martín, V.; Sevilla, N. Vaccination as a Strategy to Prevent Bluetongue Virus Vertical Transmission. Pathogens 2021, 10, 1528. https://doi.org/10.3390/pathogens10111528
Rojas JM, Martín V, Sevilla N. Vaccination as a Strategy to Prevent Bluetongue Virus Vertical Transmission. Pathogens. 2021; 10(11):1528. https://doi.org/10.3390/pathogens10111528
Chicago/Turabian StyleRojas, José M., Verónica Martín, and Noemí Sevilla. 2021. "Vaccination as a Strategy to Prevent Bluetongue Virus Vertical Transmission" Pathogens 10, no. 11: 1528. https://doi.org/10.3390/pathogens10111528
APA StyleRojas, J. M., Martín, V., & Sevilla, N. (2021). Vaccination as a Strategy to Prevent Bluetongue Virus Vertical Transmission. Pathogens, 10(11), 1528. https://doi.org/10.3390/pathogens10111528