
Whole-Genome Analysis of Porcine Circovirus Type 2 in Russia


Abstract
:1. Introduction
2. Results and Discussion
2.1. PCV2 Is Ubiquitous in Russia
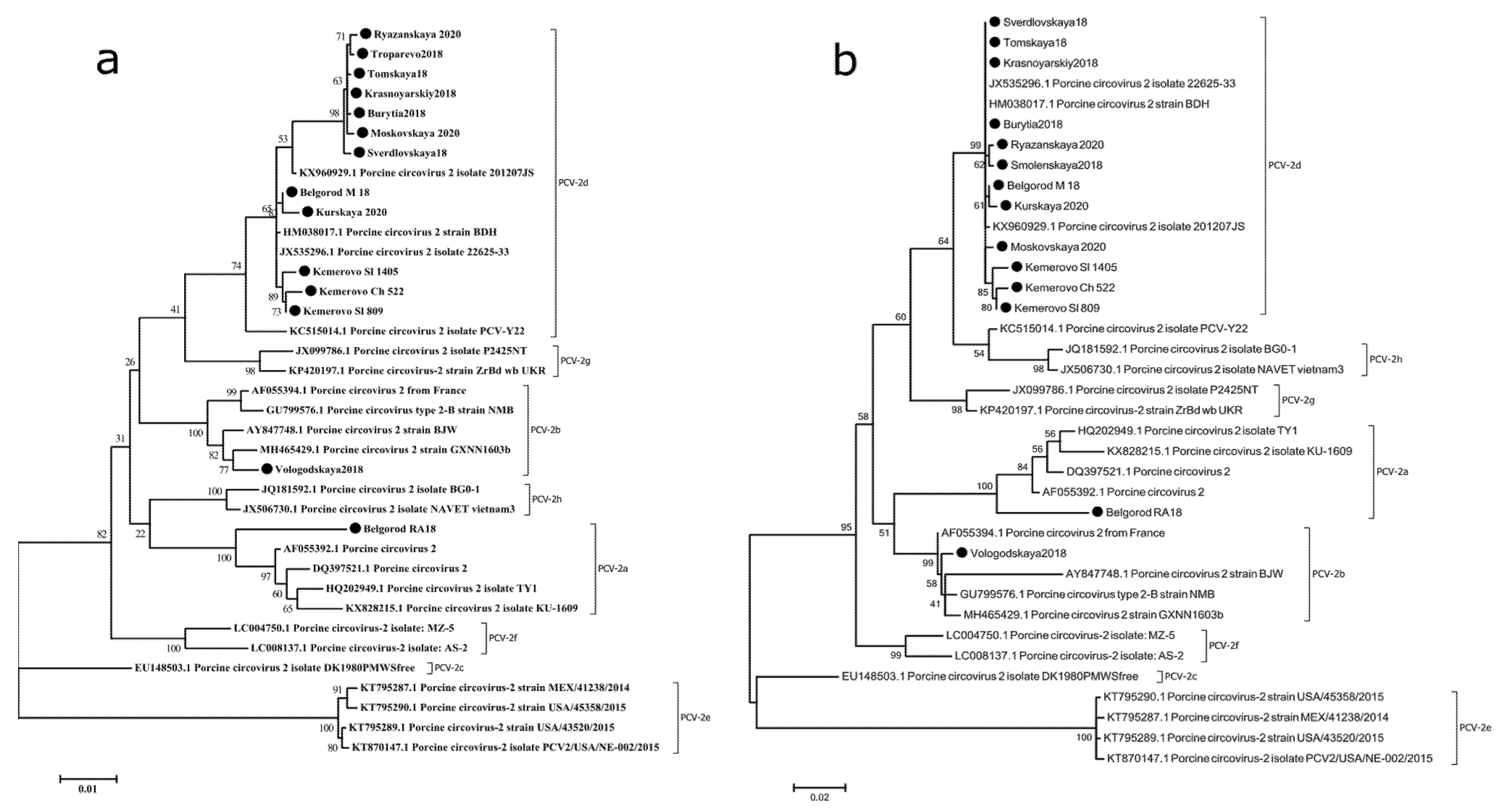
2.2. PCV-2d Is the Predominant Genotype in Russia
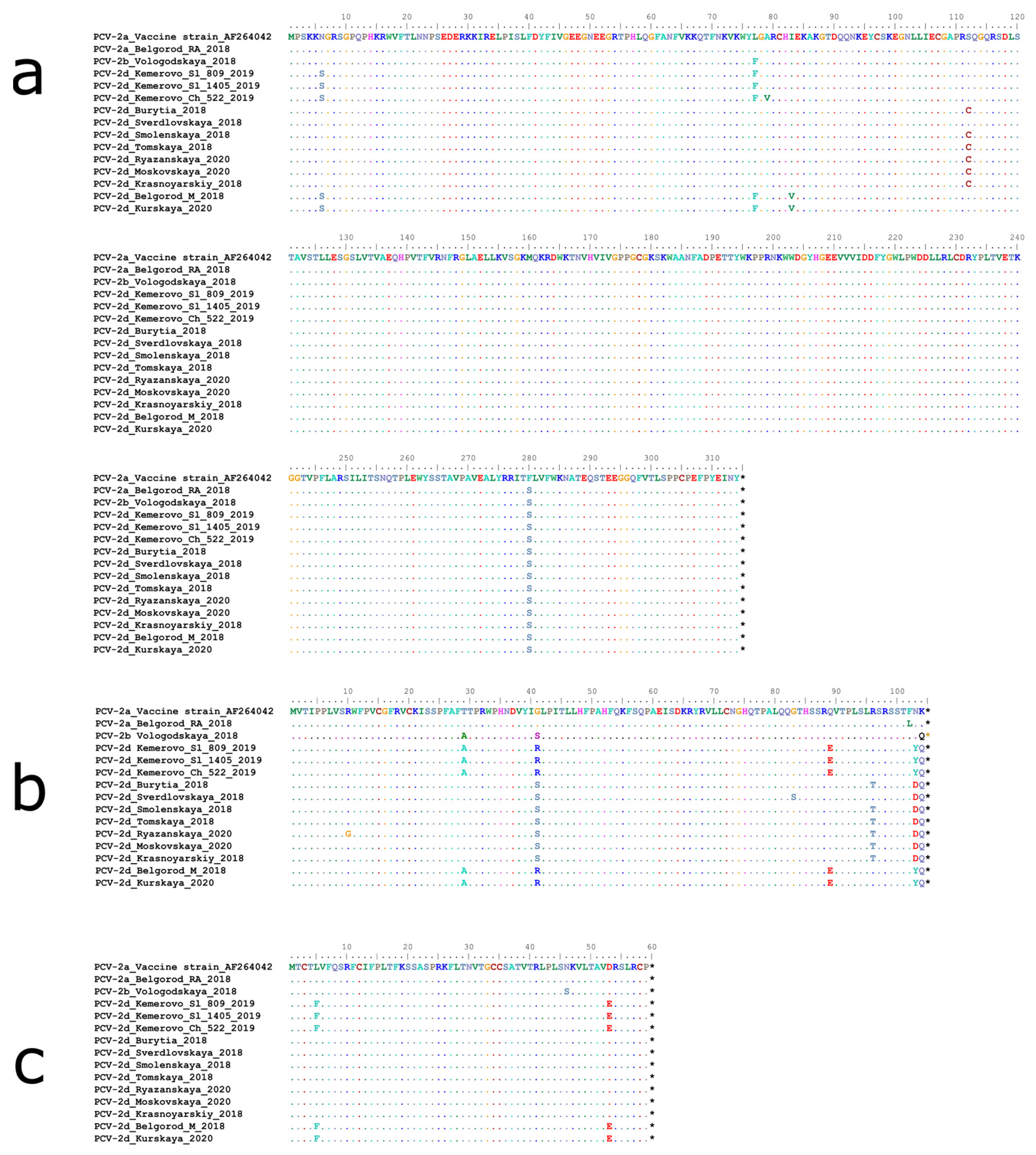
2.3. The Antibody Recognition Domains Sequences in ORF-2 Gene Are Affected to a Significant Extent in Russian PCV-2 Strains
2.4. Belgorod RA18 (PCV2a) Has a Unique Deletion in the Origin of Replication
2.5. PCV-2d Strains Can Be Classified into 3 Lines Indistinguishable on the Basis of ORF2 Sequences
3. Materials and Methods
3.1. Farms
3.2. Sample Collection, DNA Extraction
3.3. Whole-Genome Sequencing
3.4. Genome Alignment and Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saporiti, V.; Franzo, G.; Sibila, M.; Segalés, J. Porcine Circovirus 3 (PCV-3) as a Causal Agent of Disease in Swine and a Proposal of PCV-3 Associated Disease Case Definition. Transbound. Emerg. Dis. 2021, 68, 2936–2948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hu, W.; Li, J.; Liu, T.; Zhou, J.; Opriessnig, T.; Xiao, C. Novel Circovirus Species Identified in Farmed Pigs Designated as Porcine Circovirus 4, Hunan Province, China. Transbound. Emerg. Dis. 2020, 67, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J. Porcine Circovirus Type 2 (PCV2) Infections: Clinical Signs, Pathology and Laboratory Diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J. Porcine Circovirus: A Historical Perspective. Vet. Pathol. 2014, 51, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Segalés, J. Porcine Circovirus 2 Genotypes, Immunity and Vaccines: Multiple Genotypes but One Single Serotype. Pathogens 2020, 9, 1049. [Google Scholar] [CrossRef]
- Lv, Q.-Z.; Guo, K.-K.; Zhang, Y.-M. Current Understanding of Genomic DNA of Porcine Circovirus Type 2. Virus Genes 2014, 49, 1–10. [Google Scholar] [CrossRef]
- Cheung, A.K. Transcriptional Analysis of Porcine Circovirus Type 2. Virology 2003, 305, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Nawagitgul, P.; Morozov, I.; Bolin, S.R.; Harms, P.A.; Sorden, S.D.; Paul, P.S. Open Reading Frame 2 of Porcine Circovirus Type 2 Encodes a Major Capsid Protein. J. Gen. Virol. 2000, 81, 2281–2287. [Google Scholar] [CrossRef]
- Liu, J.; Chen, I.; Du, Q.; Chua, H.; Kwang, J. The ORF3 Protein of Porcine Circovirus Type 2 Is Involved in Viral Pathogenesis in Vivo. J. Virol. 2006, 80, 5065–5073. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Cao, J.; Zhou, N.; Jin, Y.; Wu, J.; Zhou, J. Identification and Functional Analysis of the Novel ORF4 Protein Encoded by Porcine Circovirus Type 2. J. Virol. 2013, 87, 1420–1429. [Google Scholar] [CrossRef] [Green Version]
- Franzo, G.; Segalés, J. Porcine Circovirus 2 (PCV-2) Genotype Update and Proposal of a New Genotyping Methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortey, M.; Pileri, E.; Sibila, M.; Pujols, J.; Balasch, M.; Plana, J.; Segalés, J. Genotypic Shift of Porcine Circovirus Type 2 from PCV-2a to PCV-2b in Spain from 1985 to 2008. Vet. J. 2011, 187, 363–368. [Google Scholar] [CrossRef]
- Franzo, G.; Cortey, M.; Segalés, J.; Hughes, J.; Drigo, M. Phylodynamic Analysis of Porcine Circovirus Type 2 Reveals Global Waves of Emerging Genotypes and the Circulation of Recombinant Forms. Mol. Phylogenet. Evol. 2016, 100, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.-T.; Harmon, K.M.; Halbur, P.G.; Opriessnig, T. PCV2d-2 Is the Predominant Type of PCV2 DNA in Pig Samples Collected in the U.S. during 2014–2016. Vet. Microbiol. 2016, 197, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Noll, L.; Lu, N.; Porter, E.; Stoy, C.; Zheng, W.; Liu, X.; Peddireddi, L.; Niederwerder, M.; Bai, J. Genetic Diversity and Prevalence of Porcine Circovirus Type 3 (PCV3) and Type 2 (PCV2) in the Midwest of the USA during 2016–2018. Transbound. Emerg. Dis. 2020, 67, 1284–1294. [Google Scholar] [CrossRef]
- Wei, C.; Lin, Z.; Dai, A.; Chen, H.; Ma, Y.; Li, N.; Wu, Y.; Yang, X.; Luo, M.; Liu, J. Emergence of a Novel Recombinant Porcine Circovirus Type 2 in China: PCV2c and PCV2d Recombinant. Transbound. Emerg. Dis. 2019, 66, 2496–2506. [Google Scholar] [CrossRef]
- Xiao, C.-T.; Halbur, P.G.; Opriessnig, T. Complete Genome Sequence of a Novel Porcine Circovirus Type 2b Variant Present in Cases of Vaccine Failures in the United States. J. Virol. 2012, 86, 12469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.W.; Park, C.; Kang, I.; Choi, K.; Jeong, J.; Park, S.-J.; Chae, C. Genetic and Antigenic Characterization of a Newly Emerging Porcine Circovirus Type 2b Mutant First Isolated in Cases of Vaccine Failure in Korea. Arch. Virol. 2014, 159, 3107–3111. [Google Scholar] [CrossRef]
- Franzo, G.; Tucciarone, C.M.; Cecchinato, M.; Drigo, M. Porcine Circovirus Type 2 (PCV2) Evolution before and after the Vaccination Introduction: A Large Scale Epidemiological Study. Sci. Rep. 2016, 6, 39458. [Google Scholar] [CrossRef]
- Kwon, T.; Lee, D.-U.; Yoo, S.J.; Je, S.H.; Shin, J.Y.; Lyoo, Y.S. Genotypic Diversity of Porcine Circovirus Type 2 (PCV2) and Genotype Shift to PCV2d in Korean Pig Population. Virus Res. 2017, 228, 24–29. [Google Scholar] [CrossRef]
- Bulgakov, A.D.; Grebennikova, T.V.; Iuzhakov, A.G.; Aliper, T.I.; Nepoklonov, E.A. Molecular-genetic analysis of the genomes of porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 circulating in the area of Russian Federation. Mol. Genet. Mikrobiol. Virusol. 2014, 4, 29–33. [Google Scholar]
- Saha, D.; Lefebvre, D.J.; Ooms, K.; Huang, L.; Delputte, P.L.; Van Doorsselaere, J.; Nauwynck, H.J. Single Amino Acid Mutations in the Capsid Switch the Neutralization Phenotype of Porcine Circovirus 2. J. Genet. Virol. 2012, 93, 1548–1555. [Google Scholar] [CrossRef]
- Bandrick, M.; Gutiérrez, A.H.; Desai, P.; Rincon, G.; Martin, W.D.; Terry, F.E.; De Groot, A.S.; Foss, D.L. T Cell Epitope Content Comparison (EpiCC) Analysis Demonstrates a Bivalent PCV2 Vaccine Has Greater T Cell Epitope Overlap with Field Strains than Monovalent PCV2 Vaccines. Vet. Immunol. Immunopathol. 2020, 223, 110034. [Google Scholar] [CrossRef] [PubMed]
- Trible, B.R.; Kerrigan, M.; Crossland, N.; Potter, M.; Faaberg, K.; Hesse, R.; Rowland, R.R.R. Antibody Recognition of Porcine Circovirus Type 2 Capsid Protein Epitopes after Vaccination, Infection, and Disease. Clin. Vaccine Immunol. 2011, 18, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, A.K. Identification of an Octanucleotide Motif Sequence Essential for Viral Protein, DNA, and Progeny Virus Biosynthesis at the Origin of DNA Replication of Porcine Circovirus Type 2. Virology 2004, 324, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olvera, A.; Cortey, M.; Segalés, J. Molecular Evolution of Porcine Circovirus Type 2 Genomes: Phylogeny and Clonality. Virology 2007, 357, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Pair | Primer Location (Numbers Correspond to Position within the PCV2 Genome) | Primer Sequence (5′-3′) | Amplicon Size, nt | Ta, °C |
|---|---|---|---|---|
| PCV2_1F | 229 | GGTTCGCTAATTTTGTGAAGA | 596 | 56 |
| PCV2_1R | 824 | GGTCTGATTGCTGGTAATCA | ||
| PCV2_2F | 686 | TGTTATTGATGACTTTTATG | 641 | 48 |
| PCV2_2R | 1326 | TATGTAAACTACTCCTCCC | ||
| PCV2_3F | 1073 | TTAAATTCTCTAAATTGTAC | 470 | 46 |
| PCV2_3R | 1542 | GTGGACATGATGAGATT | ||
| PCV2_4F | 1443 | TAACCTTTCTTATTCTGTA | 683 | 46 |
| PCV2_4R | 359 | TAAGTTGCCTTCTTTACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raev, S.; Yuzhakov, A.; Aliper, T. Whole-Genome Analysis of Porcine Circovirus Type 2 in Russia. Pathogens 2021, 10, 1631. https://doi.org/10.3390/pathogens10121631
Raev S, Yuzhakov A, Aliper T. Whole-Genome Analysis of Porcine Circovirus Type 2 in Russia. Pathogens. 2021; 10(12):1631. https://doi.org/10.3390/pathogens10121631
Chicago/Turabian StyleRaev, Sergei, Anton Yuzhakov, and Taras Aliper. 2021. "Whole-Genome Analysis of Porcine Circovirus Type 2 in Russia" Pathogens 10, no. 12: 1631. https://doi.org/10.3390/pathogens10121631
APA StyleRaev, S., Yuzhakov, A., & Aliper, T. (2021). Whole-Genome Analysis of Porcine Circovirus Type 2 in Russia. Pathogens, 10(12), 1631. https://doi.org/10.3390/pathogens10121631