
Molecular Detection of Zoonotic and Non-Zoonotic Pathogens from Wild Boars and Their Ticks in the Corsican Wetlands


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Tick Identification
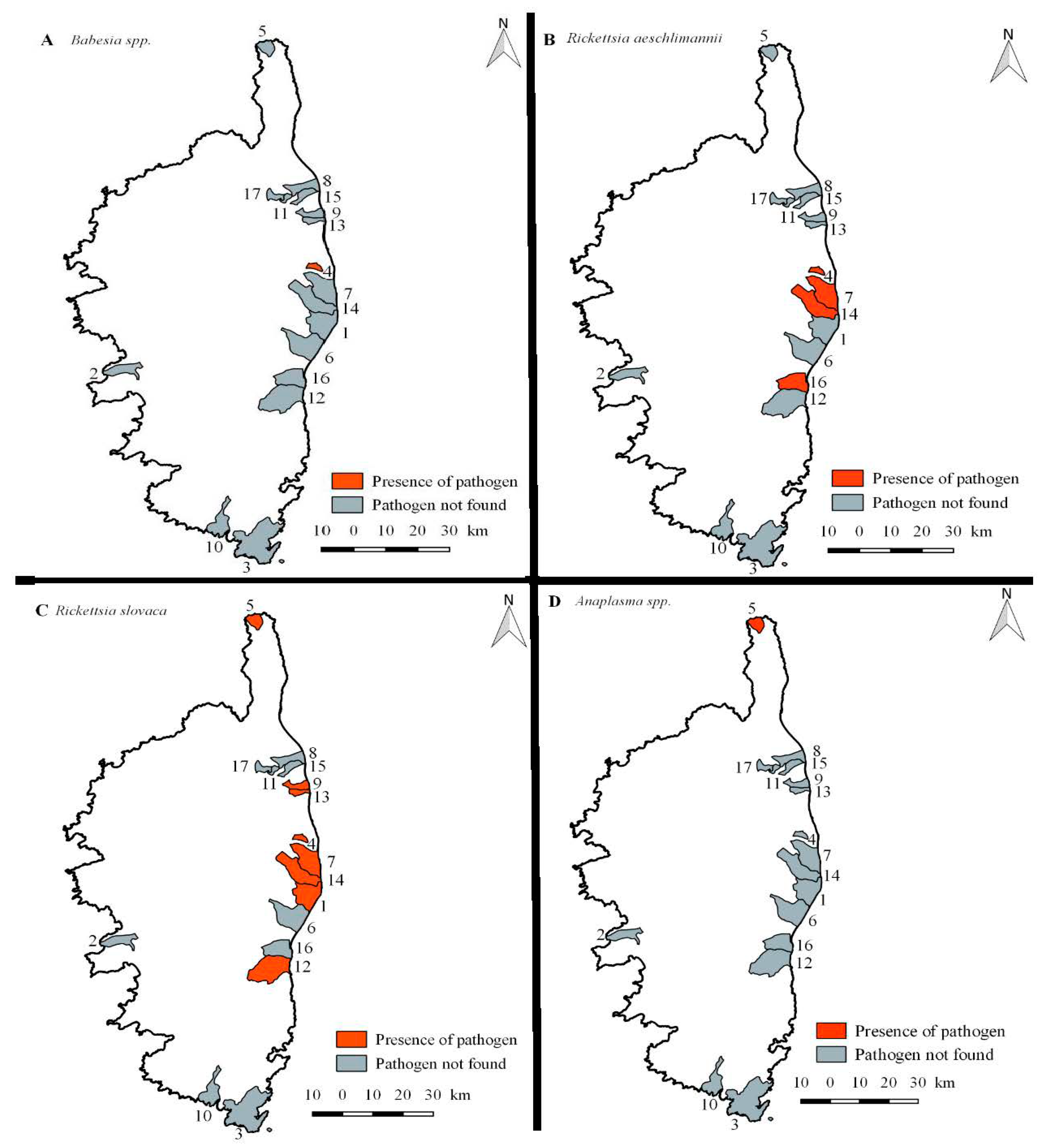
2.2. Detection of Tick-Borne Pathogen DNA in Wild Boar Samples
2.3. Detection of Tick-Borne Pathogen DNA in Ticks
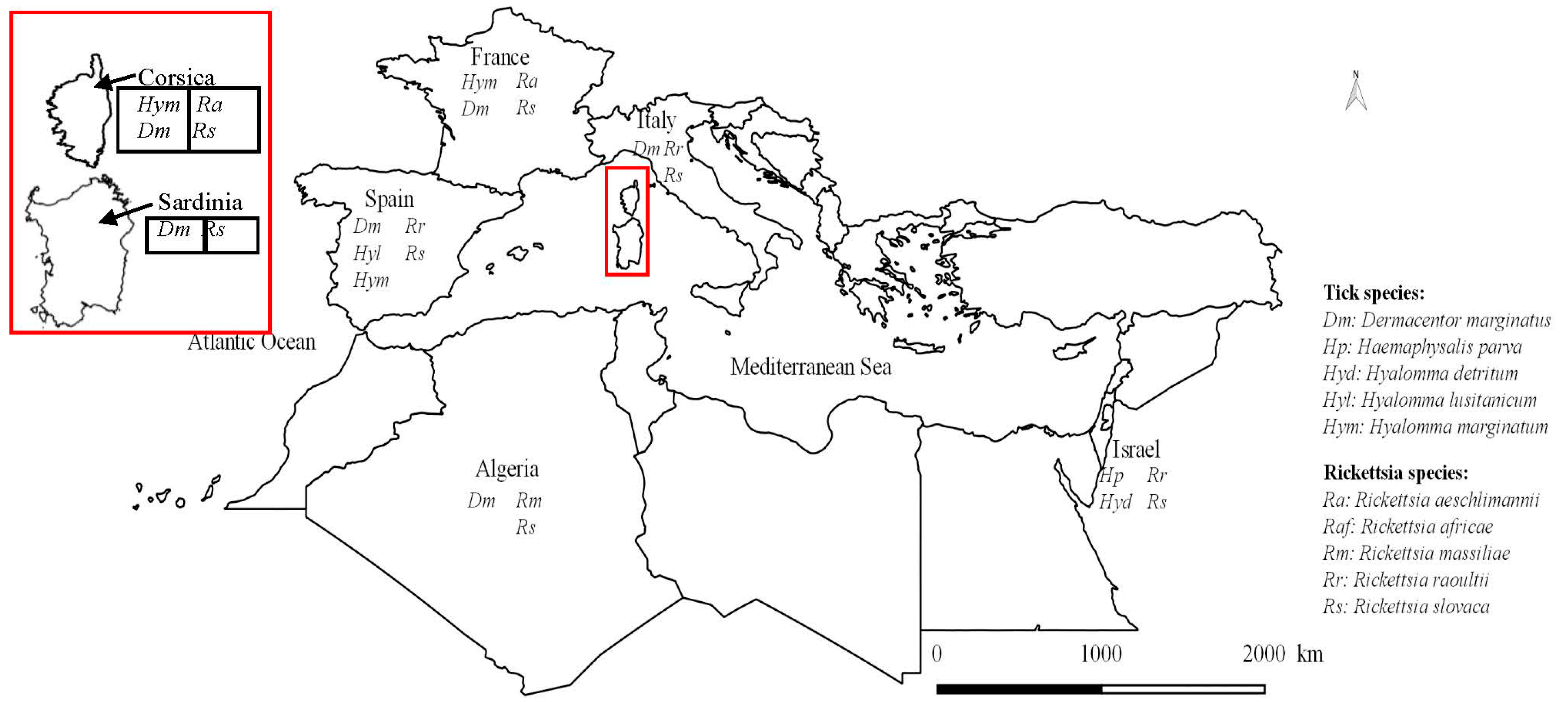
3. Discussion
3.1. Ticks on Wild Boar Population in Corsican Wetland

{kind=link}
{kind=link}
| Area (Island) | Main Tick Species | Other Tick Species | Pathogens Found in Ticks (Bacterium b, Parasite p and Virus v) | Pathogens Found in Wild Boars (Bacterium b, Parasite p and Virus v) | References |
|---|---|---|---|---|---|
| Algeria | D. marginatus | H. punctata, Rh. sanguineus | R. massiliaeb, R. slovacab | Not found | [46] |
| Algeria | Not screened | R. slovacab | [47] | ||
| France (Corsica) | D. marginatus | Hy. marginatum, H.scupense, I. ricinus | A. phagocytophilumb, R. spp. b, R. aeschlimannii b, R. slovaca b | Babesia spp. b | Present study |
| France (Corsica) | D.marginatus | Hy. marginatum, I. ricinus, Rh. bursa | R. aeschlimanniib, R. slovacab | Not screened | [32] |
| France (Corsica) | D. marginatus | Hy. marginatum, I. ricinus, Rh. bursa, Rh. sanguineus | A. phagocytophilumb, Ba. henselaeb, R. aeschlimanniib, R. slovacab | Not screened | [33] |
| France (Corsica) | Hy. marginatum | R. aeschlimanniib | Not screened | [48] | |
| France | Not screened | Tick-borne encephalitis virus v, Louping ill v | [49] | ||
| France | Not screened | A. phagocytophilumb | [50] | ||
| France | D. marginatus | Rickettsia spp. b, R. slovaca b | Not search | [51] | |
| Israel | H. parva | H. aleri, Hyalomma spp., Rh. turanicus | R. massiliaeb | Not screened | [52] |
| Israel | Hy. detritum | R. africaeb | Not screened | [53] | |
| Italy | Not screened | A. phagocytophilumb, Bo. burgdoferi s.l. b | [54] | ||
| Italy | D. marginatus | R. raoultiib, R. slovacab | Not screened | [55] | |
| Italy (Sardinia) | D. marginatus | R.slovacab | Not screened | [56] | |
| Italy (Sardinia) | D. marginatus | R. spp. b | Not screened | [57] | |
| Italy | D. marginatus | R. slovacab | Not screened | [58] | |
| Italy | D. marginatus | E. canisb | Not screened | [59] | |
| Italy | Not screened | B.bigeminap, Theileria spp. p | [60] | ||
| Slovenia | Not screened | A. phagocytophilumb | [61] | ||
| Slovenia | Not screened | A. phagocytophilumb | [62] | ||
| Spain | Hy. marginatum | D. marginatus, Rh. bursa | A. marginale b, A. phagocytophilum b, Ehrlichia spp. b, Piroplasmid p Rickettsia spp. b, R. slovaca b | Not screened | [22] |
| Spain | D. marginatus | D. reticulatus, Rh. bursa | Not found | C. burnetiib | [63] |
| Spain | D. marginatus | R.slovacab | Not screened | [64] | |
| Spain | D. marginatus | R. slovacab | Not screened | [65] | |
| Spain | Hy. lusitanicum | D. marginatus | R. raoultiib, R. slovacab | Not screened | [66] |
| Turkey | D. marginatus | H. parva, Hy. excavatum, Hy. marginatum, Rh. turanicus | B. crassap, B.occultansp, B. rossip | Not found | [67] |
3.2. Pathogens Detected in Wild Boar and Their Ticks in the Corsican Wetlands
3.2.1. Babesia spp.
3.2.2. Rickettsia spp.
3.2.3. Anaplasma spp.
4. Materials and Methods
4.1. Animals and Sampling
4.2. DNA Extraction and PCR Pre-Amplification
4.3. Assay Design
4.4. DNA Amplification and Microfluidic Real-Time PCR
4.5. Confirmation of Pathogen Detection
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Massei, G.; Genov, P.V. The environmental impact of Wild Boar. Galemys 2004, 16, 135–145. [Google Scholar]
- Courchamp, F.; Chapuis, J.-L.; Pascal, M. Mammal invaders on islands: Impact, control and control impact. Biol. Rev. 2003, 78, 347–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulding, M.J. Possible genetic sources of free-living wild boar (Sus scrofa) in southern England. Mammal. Rev. 2001, 31, 245–248. [Google Scholar] [CrossRef]
- Massei, G.; Genov, P.V.; Staines, B.W. Diet, food availability and reproduction of wild boar in a mediterranean coastal area. Acta Theriol. 1996, 41, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Barth, S.A.; Blome, S.; Cornelis, D.; Pietschmann, J.; Laval, M.; Maestrini, O.; Geue, L.; Charrier, F.; Etter, E.; Menge, C.; et al. Faecal Escherichia coli as biological indicator of spatial interaction between domestic pigs and wild boar (Sus scrofa) in Corsica. Transbound. Emerg. Dis. 2018, 65, 746–757. [Google Scholar] [CrossRef] [Green Version]
- Barasona, J.A.; Latham, M.C.; Acevedo, P.; Armenteros, J.A.; Latham, A.D.M.; Gortazar, C.; Carro, F.; Soriguer, R.C.; Vicente, J. Spatiotemporal interactions between wild boar and cattle: Implications for cross-species disease transmission. Vet. Res. 2014, 45, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowie, C.E.; Hutchings, M.R.; Barasona, J.A.; Gortázar, C.; Vicente, J.; White, P.C.L. Interactions between four species in a complex wildlife: Livestock disease community: Implications for Mycobacterium bovis maintenance and transmission. Eur. J. Wildl. Res. 2016, 62, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Kazimírová, M.; Hamšíková, Z.; Špitalská, E.; Minichová, L.; Mahríková, L.; Caban, R.; Sprong, H.; Fonville, M.; Schnittger, L.; Kocianová, E. Diverse tick-borne microorganisms identified in free-living ungulates in Slovakia. Parasit. Vectors 2018, 11, 495. [Google Scholar] [CrossRef]
- Kriz, B.; Daniel, M.; Benes, C.; Maly, M. The role of game (wild boar and roe deer) in the spread of tick-borne encephalitis in the Czech Republic. Vector-Borne Zoonotic Dis. 2014, 14, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, V.; Gortazar, C.; Vicente, J.; de la Fuente, J. Evidence of the role of european wild boar as a reservoir of Mycobacterium tuberculosis complex. Vet. Microbiol. 2008, 127, 1–9. [Google Scholar] [CrossRef]
- Silaghi, C.; Pfister, K.; Overzier, E. Molecular investigation for bacterial and protozoan tick-borne pathogens in wild boars (Sus scrofa) from southern Germany. Vector-Borne Zoonotic Dis. 2014, 14, 371–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrier, F.; Rossi, S.; Jori, F.; Maestrini, O.; Richomme, C.; Casabianca, F.; Ducrot, C.; Jouve, J.; Pavio, N.; Le Potier, M.-F. Aujeszky’s disease and Hepatitis E Viruses transmission between domestic pigs and wild boars in corsica: Evaluating the importance of wild/domestic interactions and the efficacy of management measures. Front. Vet. Sci. 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Vanschoenwinkel, B.; Waterkeyn, A.; Vandecaetsbeek, T.; Pineau, O.; Grillas, P.; Brendonck, L. Dispersal of freshwater invertebrates by large terrestrial mammals: A case study with wild boar (Sus scrofa) in mediterranean wetlands. Freshw. Biol. 2008, 53, 2264–2273. [Google Scholar] [CrossRef]
- Bacon, P.R. Use of wetlands for tourism in the insular caribbean. Ann. Tour. Res. 1987, 14, 104–117. [Google Scholar] [CrossRef]
- Hammer, D.A. Constructed Wetlands for Wastewater Treatment: Municipal, Industrial and Agricultural; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Jourdain, E.; Gauthier-Clerc, M.; Bicout, D.; Sabatier, P. Bird migration routes and risk for pathogen dispersion into western mediterranean wetlands. Emerg. Infect. Dis. 2007, 13, 365–372. [Google Scholar] [CrossRef]
- Rabou, A.F.N.A.; Yassin, M.M.; Sharif, F.A.; Al-Agha, M.R.; Daher, K.S.A.; Ali, A.K.S.; Hamad, D.M. Inhabitants knowledge on mosquito proliferation and control in the wetland ecosystem of Wadi Gaza, Gaza Strip, Palestine. IUG J. Nat. Stud. 2015, 16, 41–61. [Google Scholar]
- Rey, J.R.; Walton, W.E.; Wolfe, R.J.; Connelly, R.; O’Connell, S.M.; Berg, J.; Sakolsky-Hoopes, G.E.; Laderman, A.D. North american wetlands and mosquito control. Int. J. Environ. Res. Public. Health 2012, 9, 4537–4605. [Google Scholar] [CrossRef] [PubMed]
- Dudek, K. Impact of biodiversity on tick-borne diseases. Przegl. Epidemiol. 2014, 68, 681–684. [Google Scholar] [PubMed]
- De la Fuente, J.; Gortazar, C. Wild Boars as hosts of human-pathogenic Anaplasma Phagocytophilum variants. Emerg. Infect. Dis. 2012, 18, 2094–2095. [Google Scholar] [CrossRef] [Green Version]
- Fredriksson-Ahomaa, M. Wild boar: A reservoir of foodborne zoonoses. Foodborne Pathog. Dis. 2018, 16, 153–165. [Google Scholar] [CrossRef]
- De la Fuente, J.; Naranjo, V.; Ruiz-Fons, F.; Vicente, J.; Estrada-Peña, A.; Almazán, C.; Kocan, K.M.; Martín, M.P.; Gortázar, C. Prevalence of tick-borne pathogens in ixodid ticks (Acari: Ixodidae) collected from european wild boar (Sus scrofa) and iberian red deer (Cervus elaphus hispanicus) in central Spain. Eur. J. Wildl. Res. 2004, 50, 187–196. [Google Scholar] [CrossRef]
- Grech-Angelini, S.; Hervé, S.; Rose, N.; Barbier, N.; Casabianca, F.; Maestrini, O.; Falchi, A.; Simon, G. Serological survey of influenza a viruses in domestic and wild suidae in Corsica (France), a mediterranean island environment. Prev. Vet. Med. 2018, 157, 94–98. [Google Scholar] [CrossRef]
- Foata, J.; Mouillot, D.; Culioli, J.-L.; Marchand, B. Influence of season and host age on wild boar parasites in Corsica using indicator species analysis. J. Helminthol. 2006, 80, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Oncfs—Grands Ongulés—Tableaux de Chasse Départementaux. Available online: http://www.oncfs.gouv.fr/Grands-ongules-Tableaux-de-chasse-ru248/Grands-ongules-Tableaux-de-chasse-departementaux-ar1480 (accessed on 18 February 2020).
- Pavio, N.; Laval, M.; Maestrini, O.; Casabianca, F.; Charrier, F.; Jori, F. Possible foodborne transmission of hepatitis e virus from domestic pigs and wild boars from Corsica. Emerg. Infect. Dis. 2016, 22, 2194–2199. [Google Scholar] [CrossRef] [Green Version]
- Richomme, C.; Lacour, S.A.; Ducrot, C.; Gilot-Fromont, E.; Casabianca, F.; Maestrini, O.; Vallée, I.; Grasset, A.; van der Giessen, J.; Boireau, P. Epidemiological survey of trichinellosis in wild boar (Sus scrofa) and fox (Vulpes vulpes) in a french insular region, Corsica. Vet. Parasitol. 2010, 172, 150–154. [Google Scholar] [CrossRef]
- Umhang, G.; Richomme, C.; Hormaz, V.; Boucher, J.-M.; Boué, F. Pigs and wild boar in Corsica harbor Echinococcus canadensis G6/7 at levels of concern for public health and local economy. Acta Trop. 2014, 133, 64–68. [Google Scholar] [CrossRef]
- Richomme, C.; Aubert, D.; Gilot-Fromont, E.; Ajzenberg, D.; Mercier, A.; Ducrot, C.; Ferté, H.; Delorme, D.; Villena, I. Genetic characterization of Toxoplasma gondii from wild boar (Sus scrofa) in France. Vet. Parasitol. 2009, 164, 296–300. [Google Scholar] [CrossRef]
- Richomme, C.; Afonso, E.; Tolon, V.; Ducrot, C.; Halos, L.; Alliot, A.; Perret, C.; Thomas, M.; Boireau, P.; Gilot-Fromont, E. Seroprevalence and factors associated with Toxoplasma gondii infection in wild boar (Sus scrofa) in a mediterranean island. Epidemiol. Infect. 2010, 138, 1257–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grech-Angelini, S.; Stachurski, F.; Lancelot, R.; Boissier, J.; Allienne, J.-F.; Marco, S.; Maestrini, O.; Uilenberg, G. Ticks (Acari: Ixodidae) infesting cattle and some other domestic and wild hosts on the french mediterranean island of Corsica. Parasit. Vectors 2016, 9, 582. [Google Scholar] [CrossRef]
- Cicculli, V.; Maestrini, O.; Casabianca, F.; Villechenaud, N.; Charrel, R.; de Lamballerie, X.; Falchi, A. Molecular detection of spotted-fever group Rickettsiae in ticks collected from domestic and wild animals in Corsica, France. Pathogens 2019, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Grech-Angelini, S.; Stachurski, F.; Vayssier-Taussat, M.; Devillers, E.; Casabianca, F.; Lancelot, R.; Uilenberg, G.; Moutailler, S. Tick-borne pathogens in ticks (Acari: Ixodidae) collected from various domestic and wild hosts in corsica (France), a mediterranean island environment. Transbound. Emerg. Dis. 2020, 67, 745–757. [Google Scholar] [CrossRef]
- Selmi, M.; Ballardini, M.; Salvato, L.; Ricci, E. Rickettsia Spp. in Dermacentor marginatus ticks: Analysis of the host-vector-pathogen interactions in a northern mediterranean area. Exp. Appl. Acarol. 2017, 72, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Hrazdilová, K.; Lesiczka, P.M.; Bardoň, J.; Vyroubalová, Š.; Šimek, B.; Zurek, L.; Modrý, D. Wild boar as a potential reservoir of zoonotic tick-borne pathogens. Ticks Tick-Borne Dis. 2021, 12, 101558. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Raoult, D. Ticks and tickborne bacterial diseases in humans: An emerging infectious threat. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2001, 32, 897–928. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L.; Sonenshine, D.E.; Noden, B.H.; Brown, R.N. Chapter 27—Ticks (Ixodida). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 603–672. [Google Scholar] [CrossRef]
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. J. Virtual Libr. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [Green Version]
- Sonenshine, E.D. Biology of Ticks; Oxford University Press: New York, NY, USA; Oxford, UK, 1991; Volume 1. [Google Scholar]
- Eldridge, B.F.; Edman, J. Medical Entomology: A Textbook on Public Health and Veterinary Problems Caused by Arthropods; Springer Science & Business Media: Berlin, Germany, 2003. [Google Scholar]
- Pérez-Eid, C. Les Tiques: Identification, Biologie, Importance Médicale et Vétérinaire; Lavoisier: Cachan, France, 2007. [Google Scholar]
- Cicculli, V.; Capai, L.; Quilichini, Y.; Masse, S.; Fernández-Alvarez, A.; Minodier, L.; Bompard, P.; Charrel, R.; Falchi, A. Molecular investigation of tick-borne pathogens in ixodid ticks infesting domestic animals (cattle and sheep) and small rodents (black rats) of Corsica, France. Ticks Tick-Borne Dis. 2019, 10, 606–613. [Google Scholar] [CrossRef]
- Cicculli, V.; de Lamballerie, X.; Charrel, R.; Falchi, A. First molecular detection of Rickettsia africae in a tropical bont tick, Amblyomma variegatum, collected in Corsica, France. Exp. Appl. Acarol. 2019, 77, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A. Ticks of Domestic Animals in the Mediterranean Region: A Guide to Identification of Species; University of Zaragoza: Zaragoza, Aragon, Spain, 2004. [Google Scholar]
- Raad, M.; Azar, D.; Perotti, M.A. First report of the ticks Haemaphysalis punctata Canestrini et Fanzago, 1878, Haemaphysalis parva (Neumann, 1897) and Dermacentor marginatus (Sulzer, 1776) (Acari, Amblyommidae) from humans in Lebanon. Acta Parasitol. 2020, 65, 541–545. [Google Scholar] [CrossRef] [Green Version]
- Leulmi, H.; Aouadi, A.; Bitam, I.; Bessas, A.; Benakhla, A.; Raoult, D.; Parola, P. Detection of Bartonella tamiae, Coxiella burnetii and Rickettsiae in arthropods and tissues from wild and domestic animals in northeastern Algeria. Parasit. Vectors 2016, 9, 27. [Google Scholar] [CrossRef]
- Zeroual, F.; Leulmi, H.; Bitam, I.; Benakhla, A. Molecular evidence of Rickettsia slovaca in spleen of wild boars in northeastern Algeria. New Microbes New Infect. 2018, 24, 17–20. [Google Scholar] [CrossRef]
- Matsumoto, K.; Parola, P.; Brouqui, P.; Raoult, D. Rickettsia aeschlimannii in Hyalomma ticks from Corsica. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 732–734. [Google Scholar] [CrossRef]
- Bournez, L.; Umhang, G.; Faure, E.; Boucher, J.-M.; Boué, F.; Jourdain, E.; Sarasa, M.; Llorente, F.; Jiménez-Clavero, M.A.; Moutailler, S.; et al. Exposure of wild ungulates to the Usutu and Tick-borne encephalitis viruses in France in 2009-2014: Evidence of undetected flavivirus circulation a decade ago. Viruses 2019, 12, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugat, T.; Zanella, G.; Véran, L.; Lesage, C.; Girault, G.; Durand, B.; Lagrée, A.-C.; Boulouis, H.-J.; Haddad, N. Multiple-locus variable-number tandem repeat analysis potentially reveals the existence of two groups of Anaplasma phagocytophilum circulating in cattle in France with different wild reservoirs. Parasit. Vectors 2016, 9, 596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanogo, Y.O.; Davoust, B.; Parola, P.; Camicas, J.L.; Brouqui, P.; Raoult, D. Prevalence of Rickettsia spp. in Dermacentor marginatus ticks removed from game pigs (Sus scrofa) in southern France. Ann. N. Y. Acad. Sci. 2003, 990, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Keysary, A.; Eremeeva, M.E.; Leitner, M.; Din, A.B.; Wikswo, M.E.; Mumcuoglu, K.Y.; Inbar, M.; Wallach, A.D.; Shanas, U.; King, R.; et al. Spotted fever group Rickettsiae in ticks collected from wild animals in Israel. Am. J. Trop. Med. Hyg. 2011, 85, 919–923. [Google Scholar] [CrossRef] [Green Version]
- Waner, T.; Keysary, A.; Eremeeva, M.E.; Din, A.B.; Mumcuoglu, K.Y.; King, R.; Atiya-Nasagi, Y. Rickettsia africae and Candidatus Rickettsia barbariae in ticks in Israel. Am. J. Trop. Med. Hyg. 2014, 90, 920–922. [Google Scholar] [CrossRef] [Green Version]
- Ebani, V.; Bertelloni, F.; Cecconi, G.; Sgorbini, M.; Cerri, D. Zoonotic tick-borne bacteria among wild boars (Sus scrofa) in central Italy. Asian Pac. J. Trop. Dis. 2017, 7, 141–143. [Google Scholar] [CrossRef]
- Raele, D.A.; Galante, D.; Pugliese, N.; Salandra, G.L.; Cafiero, M.A. Spotted fever group Rickettsiae associated with ixodid ticks in wild environment in southern Italy. MicrobiologyOpen 2018, 7, e527. [Google Scholar] [CrossRef] [Green Version]
- Chisu, V.; Masala, G.; Foxi, C.; Socolovschi, C.; Raoult, D.; Parola, P. Rickettsia conorii israelensis in Rhipicephalus sanguineus ticks, Sardinia, Italy. Ticks Tick-Borne Dis. 2014, 5, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Chisu, V.; Foxi, C.; Mannu, R.; Satta, G.; Masala, G. A five-year survey of tick species and identification of tick-borne bacteria in Sardinia, Italy. Ticks Tick-Borne Dis. 2018, 9, 678–681. [Google Scholar] [CrossRef]
- Pascucci, I.; Di Domenico, M.; Curini, V.; Cocco, A.; Averaimo, D.; D’Alterio, N.; Cammà, C. Diversity of Rickettsia in ticks collected in Abruzzi and Molise regions (central Italy). Microorganisms 2019, 7, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satta, G.; Chisu, V.; Cabras, P.; Fois, F.; Masala, G. Pathogens and symbionts in ticks: A survey on tick species distribution and presence of tick-transmitted micro-organisms in Sardinia, Italy. J. Med. Microbiol. 2011, 60, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Zanet, S.; Trisciuoglio, A.; Bottero, E.; de Mera, I.G.F.; Gortazar, C.; Carpignano, M.G.; Ferroglio, E. Piroplasmosis in wildlife: Babesia and Theileria affecting free-ranging ungulates and carnivores in the italian alps. Parasit Vectors 2014, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Smrdel, K.S.; Bidovec, A.; Malovrh, T.; Petrovec, M.; Duh, D.; Zupanc, T.A. Detection of Anaplasma phagocytophilum in wild boar in Slovenia. Clin. Microbiol. Infect. 2009, 15, 50–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zele, D.; Avberšek, J.; Gruntar, I.; Ocepek, M.; Vengušt, G. Evidence of Anaplasma phagocytophilum in game animals from Slovenia. Acta Vet. Hung. 2012, 60, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Astobiza, I.; Barral, M.; Ruiz-Fons, F.; Barandika, J.F.; Gerrikagoitia, X.; Hurtado, A.; García-Pérez, A.L. Molecular investigation of the occurrence of Coxiella burnetii in wildlife and ticks in an endemic area. Vet. Microbiol. 2011, 147, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Oteo, J.A.; Portillo, A.; Santibáñez, S.; Pérez-Martínez, L.; Blanco, J.R.; Jiménez, S.; Ibarra, V.; Pérez-Palacios, A.; Sanz, M. Prevalence of spotted fever group Rickettsia species detected in ticks in La Rioja, Spain. Ann. N. Y. Acad. Sci. 2006, 1078, 320–323. [Google Scholar] [CrossRef]
- Ortuño, A.; Quesada, M.; López, S.; Miret, J.; Cardeñosa, N.; Castellà, J.; Anton, E.; Segura, F. Prevalence of Rickettsia slovaca in Dermacentor marginatus ticks removed from wild boar (Sus scrofa) in northeastern Spain. Ann. N. Y. Acad. Sci. 2006, 1078, 324–327. [Google Scholar] [CrossRef]
- Márquez, F.J. Spotted fever group Rickettsia in ticks from southeastern Spain natural parks. Exp. Appl. Acarol. 2008, 45, 185–194. [Google Scholar] [CrossRef]
- Orkun, Ö.; Karaer, Z. Molecular characterization of Babesia species in wild animals and their ticks in Turkey. Infect. Genet. Evol. 2017, 55, 8–13. [Google Scholar] [CrossRef]
- Beuglet, F.; Moreau, Y. Babesiosis. New Dev. Major Vector-Borne Dis. Part 2 Important Dis. Vet. 2015, 34, 627–639. [Google Scholar] [CrossRef]
- Vannier, E.G.; Diuk-Wasser, M.A.; Ben Mamoun, C.; Krause, P.J. Babesiosis. Infect. Dis. Clin. N. Am. 2015, 29, 357–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoust, B.; Mediannikov, O.; Marié, J.-L.; Socolovschi, C.; Parola, P.; Raoult, D. Les animaux vertébrés sont-ils réservoirs de rickettsies? Bull. Académie Vét. Fr. 2010, 163, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Blanton, L.S. The rickettsioses: A practical update. Infect. Dis. Clin. N. Am. 2019, 33, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Nicolas, D.; De Micco, P.; Gallais, H.; Casanova, P. Epidemiologic aspects of mediterranean boutonneuse fever in the south of Corsica. Bull. Soc. Pathol. Exot. Filiales 1985, 78, 446–451. [Google Scholar] [PubMed]
- Rehácěk, J. Rickettsia slovaca, the Organism and Its Ecology; Academia: San Francisco, CA, USA, 1984. [Google Scholar]
- De Sousa, R.; Pereira, B.I.; Nazareth, C.; Cabral, S.; Ventura, C.; Crespo, P.; Marques, N.; da Cunha, S. Rickettsia slovaca infection in humans, Portugal. Emerg. Infect. Dis. 2013, 19, 1627–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouqui, P.; Parola, P.; Fournier, P.E.; Raoult, D. Spotted fever rickettsioses in southern and eastern europe. FEMS Immunol. Med. Microbiol. 2007, 49, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Selmi, M.; Martello, E.; Bertolotti, L.; Bisanzio, D.; Tomassone, L. Rickettsia slovaca and Rickettsia raoultii in Dermacentor marginatus ticks collected on wild boars in Tuscany, Italy. J. Med. Entomol. 2009, 46, 1490–1493. [Google Scholar] [CrossRef]
- Spitalská, E.; Stefanidesová, K.; Kocianová, E.; Boldiš, V. Rickettsia slovaca and Rickettsia raoultii in Dermacentor marginatus and Dermacentor reticulatus ticks from Slovak Republic. Exp. Appl. Acarol. 2012, 57, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Blanda, V.; Torina, A.; La Russa, F.; D’Agostino, R.; Randazzo, K.; Scimeca, S.; Giudice, E.; Caracappa, S.; Cascio, A.; de la Fuente, J. A retrospective study of the characterization of Rickettsia species in ticks collected from humans. Ticks Tick-Borne Dis. 2017, 8, 610–614. [Google Scholar] [CrossRef]
- Masala, G.; Chisu, V.; Satta, G.; Socolovschi, C.; Raoult, D.; Parola, P. Rickettsia slovaca from Dermacentor marginatus ticks in Sardinia, Italy. Ticks Tick-Borne Dis. 2012, 3, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Chisu, V.; Leulmi, H.; Masala, G.; Piredda, M.; Foxi, C.; Parola, P. Detection of Rickettsia hoogstraalii, Rickettsia helvetica, Rickettsia massiliae, Rickettsia slovaca and Rickettsia aeschlimannii in ticks from Sardinia, Italy. Ticks Tick-Borne Dis. 2017, 8, 347–352. [Google Scholar] [CrossRef]
- Beati, L.; Meskini, M.; Thiers, B.; Raoult, D. Rickettsia aeschlimannii Sp. Nov., a New spotted fever group Rickettsia associated with Hyalomma marginatum ticks. Int. J. Syst. Bacteriol. 1997, 47, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Raoult, D.; Fournier, P.-E.; Abboud, P.; Caron, F. First documented human Rickettsia aeschlimannii infection. Emerg. Infect. Dis. 2002, 8, 748–749. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Soto, P. Rickettsia aeschlimannii in Spain: Molecular evidence in Hyalomma marginatum and five other tick species that feed on humans. Emerg. Infect. Dis. 2003, 9, 889–890. [Google Scholar] [CrossRef]
- Battisti, E.; Urach, K.; Hodžić, A.; Fusani, L.; Hufnagl, P.; Felsberger, G.; Ferroglio, E.; Duscher, G.G. Zoonotic Pathogens in ticks from migratory birds, Italy. Emerg. Infect. Dis. 2020, 26, 2986–2988. [Google Scholar] [CrossRef]
- Wallménius, K.; Barboutis, C.; Fransson, T.; Jaenson, T.G.; Lindgren, P.-E.; Nyström, F.; Olsen, B.; Salaneck, E.; Nilsson, K. Spotted fever Rickettsia species in Hyalomma and Ixodes ticks infesting migratory birds in the european mediterranean area. Parasit. Vectors 2014, 7, 318. [Google Scholar] [CrossRef] [Green Version]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging tick-borne diseases. Clin. Microbiol. Rev. 2020, 33, e00083-18. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, G.D.; Scoles, G.A.; Burkhardt, N.Y.; Schloeder, B.; Kurtti, T.J.; Munderloh, U.G. Transovarial transmission of Francisella-like endosymbionts and Anaplasma phagocytophilum variants in Dermacentor albipictus (Acari: Ixodidae). J. Med. Entomol. 2009, 46, 625. [Google Scholar] [CrossRef] [Green Version]
- Taank, V.; Ramasamy, E.; Sultana, H.; Neelakanta, G. An efficient microinjection method to generate human anaplasmosis agent Anaplasma phagocytophilum-infected ticks. Sci. Rep. 2020, 10, 15994. [Google Scholar] [CrossRef]
- Guzman, N.; Yarrarapu, S.N.S.; Beidas, S.O. Anaplasma phagocytophilum. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Kocan, K.M.; De La Fuente, J.; Cabezas-Cruz, A. The genus Anaplasma: New challenges after reclassification. Rev. Sci. Tech. 2015, 34, 577. [Google Scholar] [CrossRef]
- Nguyen, A.H.L.; Tiawsirisup, S.; Kaewthamasorn, M. Molecular Detection and genetic characterization of Anaplasma marginale and Anaplasma platys-like (rickettsiales: Anaplasmataceae) in water buffalo from eight provinces of Thailand. BMC Vet. Res. 2020, 16, 380. [Google Scholar] [CrossRef] [PubMed]
- Cicculli, V.; Decarreaux, D.; Ayhan, N.; Casabianca, F.; Lamballerie, X.; Charrel, R.; Falchi, A. Molecular screening of Anaplasmataceae in ticks collected from cattle in Corsica, France. Exp. Appl. Acarol. 2020, 81, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Gallois, M.; Fontugne, M.; Allain, E.; Denoual, M.; Moutailler, S.; Devillers, E.; Zientara, S.; Memmi, M.; Chauvin, A.; et al. Epidemiology and genetic diversity of Anaplasma ovis in goats in Corsica, France. Parasit. Vectors 2019, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahmani, M.; Davoust, B.; Tahir, D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Molecular investigation and phylogeny of Anaplasmataceae species infecting domestic animals and ticks in Corsica, France. Parasit. Vectors 2017, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Gondard, M.; Michelet, L.; Nisavanh, A.; Devillers, E.; Delannoy, S.; Fach, P.; Aspan, A.; Ullman, K.; Chirico, J.; Hoffmann, B.; et al. Prevalence of tick-borne viruses in Ixodes ricinus assessed by high-throughput real-time PCR. Pathog. Dis. 2018, 76, fty083. [Google Scholar] [CrossRef] [Green Version]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]


| Municipalities (No) | Sampled Wild Boar/Positive | D. marginatus | Hy. marginatum | Hy. scupense | Rh. sanguineus s.l. | Total Ticks |
|---|---|---|---|---|---|---|
| - | Total/Positive (A/B/R/Ra/Rs) | Total/Positive (A/B/R/Ra/Rs) | Total/Positive (A/B/R/Ra/Rs) | Total/Positive (A/B/R/Ra/Rs) | Total/Positive (A/B/R/Ra/Rs) | Total/Positive (A/B/R/Ra/Rs) |
| Aléria (1) | 73/0 | 47/39 (Rs) | - | - | - | 47/39 (Rs) |
| Appietto (2) | 2/0 | - | - | - | - | - |
| Bonifacio (3) | 3/0 | - | - | - | - | - |
| Chiatra (4) | 11/3 (B) | 24/1 (R), 24 (Rs) | 2/1 (Ra) | - | 1/1 (Ra) | 27/1 (R), 2 (Ra), 24 (Rs) |
| Ersa (5) | 3/0 | 5/1 (A), 4 (Rs) | - | - | - | 5/1 (A), 4 (Rs) |
| Ghisonaccia (6) | 10/0 | - | - | - | - | - |
| Linguizetta (7) | 11/0 | 4/1 (Rs) | - | 1/1 (Ra) | - | 5/1 (Rs), 1(Ra) |
| Luciana (8) | 1/0 | - | - | - | - | - |
| Penta di casinca (9) | 2/0 | 8/4 (Rs) | - | - | - | 8/4 (Rs) |
| Pianottoli-Caldarello (10) | 1/0 | - | - | - | - | - |
| Prunelli-di-Casaconi (11) | 2/0 | - | - | - | - | - |
| Solaro (12) | 5/0 | 7/1 (Ra), 1 (Rs) | - | - | - | 7/1 (Ra), 1 (Rs) |
| Taglio-Isolaccio (13) | 2/0 | 5/1 (R), 3 (Rs) | - | - | - | 5/1 (R), 3 (Rs) |
| Tallone (14) | 20/0 | 8/1 (R), 2 (Ra), 4 (Rs) | - | - | - | 8/1 (R), 2 (Ra), 4 (Rs) |
| Vescovato (15) | 3/0 | - | - | - | - | - |
| Ventiseri (16) | 7/0 | 1/1 | - | - | - | 1 |
| Volpajola (17) | 2/0 | - | - | - | - | - |
| Total/Positive | 158/3 (B) | 109/1 (A), 3 (R), 3 (Ra), 80 (Rs) | 2/1 (Ra) | 1/1 (Ra) | 1/1 (Ra) | 113/1 (A), 3 (R), 6 (Ra), 80 (Rs) |
| Pathogen | Name | Sequence | Length (Nucleotide) | Genes | Genes References |
|---|---|---|---|---|---|
| Aujeszky’s virus | Aujv_gp50_F | CTTTATCGAGTACGCCGACTG | 225 | gp50 | Y14834.1 |
| Av_gp50_R | AACGGGCACTCTTGCCCC | ||||
| Av_gp50_P | CAGATCTTTGGGCGCTGCCGGC | ||||
| Ch. psitacci | Chl_psi_16S-23S_F | ACGCCGTGAATACGTTCCC | 214 | 16S-23S rRNA | U68450.1 |
| Chl_psi_16S-23S_R | AGTCAAACCGTCCTAAGACAG | ||||
| Chl_psi_16S-23S_P | CCTTGTACACACCGCCCGTCACATC | ||||
| Haemoproteus spp. | Hae_cytB_F | ATATGCATGCTACTGGTGCTAC | 240 | cytochrome B | AF465579.1 |
| Hae_cytB_R | CAAATCCATGAAACAAGTCCAGG | ||||
| Hae_cytB_P | CGGTTGCACCCCAGAAACTCATTGAC | ||||
| Leptospira spp. | Lep_Lipl32_F | CTCTATGTTTGGATTCCTGCC | 158 | LipL32 | MK541891.1 |
| Lep_Lipl32_R | CCAAGTATCAAACCAATGTGGC | ||||
| Lep_Lipl32_P | ATTGATTTTTCTTCTGGGGTAGCCGCTTTG | ||||
| Leucocytozoon spp. | Leu_cytB_F | GGGTTATGTCTTACCATGGGG | 177 | cytochrome B | KF717066.1 |
| Leu_cytB_R | AATTGCTAGTGCTACGAATGGG | ||||
| Leu_cytB_P | AAATGAGTTTTTGGGGAGCAACCGTTATTAC | ||||
| Plasmodium spp. | Pla_ssrRNA_F | ATATAGAAACTGCGAACGGCTC | 339 | ssrRNA | MK650620.1 |
| Pla_ssrRNA_R | TTTCTCAGGCTCCCTCTCC | ||||
| Pla_ssrRNA_P | CTCTAATTCCCCGTTACCCGTCATAGC | ||||
| R. monacensis | Ric_mon_F | CTCGGTGCCGGTACTTTAAAC | 192 | ompB | KU961543.1 |
| Ric_mon_R | GAGCACCGCCAATAAGAGC | ||||
| Ric_mon_P | AGTGCCGATGCAAATACTCCGGTGAC | ||||
| Trypanosoma spp. | Try_18SRNA_F | GTAATTCCAGCTCCAAAAGCG | 178 | 18SRNA | EU596263.1 |
| Try_18SRNA_R | TCAGGAAGGAACCACTCCC | ||||
| Try_18SRNA_P | ACCTCAAGGGCATGGGTCACCAATCC |
| Pathogen | Name | Sequence | Length (Nucleotide) | Genes | Genes References |
|---|---|---|---|---|---|
| Aujeszky’s virus | Aujv_gp50_F2 | AACATCCTCACCGACTTCATG | 158 | gp50 | Y14834.1 |
| Av_gp50_R2 | CTGGTAGAACGGCGTCAGG | ||||
| Av_gp50_P2 | AATCGCATCACGTCCACGCCCCC | ||||
| Ch. psitacci | Chl_psi_16S-23S_F2 | AGTAATCTTCGGCGAGCTGG | 177 | 16S-23S rRNA | U68450.1 |
| Chl_psi_16S-23S_R2 | CGCTACTTAGGGAATCTCTTT G | ||||
| Chl_psi_16S-23S_P2 | TATAAAGCTATGACCCGGAGGTCTCCG | ||||
| Haemoproteus spp. | Hae_cytB_F2 | CCTTGGGGTCAAATGAGTTTC | 231 | cytochrome B | AF465579.1 |
| Hae_cytB_R2 | AAGCCGTATCATATCCTAAAGG | ||||
| Hae_cytB_P2 | CCTGGACTTGTTTCATGGATTTGTGGAGG | ||||
| Leucocytozoon spp. | Leu_cytB_F2 | GAGTTTCTGGGGAGCTACTG | 197 | cytochrome B | KU842391.1 |
| Leu_cytB_R2 | GGATTAGTGCTACCTTGAATATG | ||||
| Leu_cytB_P2 | TGAATAAATACAATTGCTAGTGCTACGAATGG | ||||
| Plasmodium spp. | Pla_ssrRNA_F2 | TCGAGTTTCTGACCTATCAGC | 264 | ssrRNA | MK650620.1 |
| Pla_ssrRNA_R2 | AGACTTGCCCTCCAATTGTTA C | ||||
| Pla_ssrRNA_P2 | TGGCCTTGCATTGTTATTTCTTGTCACTACC | ||||
| Trypanosoma spp. | Try_18SRNA_F2 | CAACACGGGGAACTTTACCAG | 141 | 18SRNA | EU596263.1 |
| Try_18SRNA_R2 | ATCCTACTGGGCAGCTTGG | ||||
| Try_18SRNA_P2 | CAGGGTGAGGATTGACAGATTGAGTGTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Defaye, B.; Moutailler, S.; Pietri, C.; Galon, C.; Grech-Angelini, S.; Pasqualini, V.; Quilichini, Y. Molecular Detection of Zoonotic and Non-Zoonotic Pathogens from Wild Boars and Their Ticks in the Corsican Wetlands. Pathogens 2021, 10, 1643. https://doi.org/10.3390/pathogens10121643
Defaye B, Moutailler S, Pietri C, Galon C, Grech-Angelini S, Pasqualini V, Quilichini Y. Molecular Detection of Zoonotic and Non-Zoonotic Pathogens from Wild Boars and Their Ticks in the Corsican Wetlands. Pathogens. 2021; 10(12):1643. https://doi.org/10.3390/pathogens10121643
Chicago/Turabian StyleDefaye, Baptiste, Sara Moutailler, Christian Pietri, Clemence Galon, Sébastien Grech-Angelini, Vanina Pasqualini, and Yann Quilichini. 2021. "Molecular Detection of Zoonotic and Non-Zoonotic Pathogens from Wild Boars and Their Ticks in the Corsican Wetlands" Pathogens 10, no. 12: 1643. https://doi.org/10.3390/pathogens10121643
APA StyleDefaye, B., Moutailler, S., Pietri, C., Galon, C., Grech-Angelini, S., Pasqualini, V., & Quilichini, Y. (2021). Molecular Detection of Zoonotic and Non-Zoonotic Pathogens from Wild Boars and Their Ticks in the Corsican Wetlands. Pathogens, 10(12), 1643. https://doi.org/10.3390/pathogens10121643