
Minimal Dosage of Porcine Circovirus Type 2d Based Virus-like Particles to Induce Stable Protective Immunity against Infection



Abstract
:1. Introduction
2. Material and Methods
2.1. PCV2d Capsid Expression and VLP Confirmation
2.2. The Vaccine Formulation with Carbopol Adjuvant
2.3. Animal Studies
- (1)
- Pig challenge experiment: 4 weeks old, colostrum-fed, cross-bred, conventional piglets were purchased from a PCV2 seronegative farm. A total 30 piglets were tested for anti PCV2 antibodies and the only seronegative individuals were selected. They were confirmed as non-viremic for PCV2 by real-time polymerase chain reaction (PCR). The blood samples collected from the piglets were also examined for PRRSV infection using real-time PCR (Forward primer: 5’-GAAGAGAAACCCGGAGAAGC-3’; Reverse primer: 5’-GAAGAGAAACCCGGAGAAGC-3’) and PRRSV-specific antibodies were detected using ELISA (HerdCheck PRRS 3XR™, IDEXX Laboratories Inc., Westbrook, Maine, USA). All piglets were confirmed as PRRSV-free.
- (2)
- Guinea pig inoculation experiment: A total 20 Specific-pathogen-free 250 to 300 g outbred male Hartley strain guinea pigs were purchased from Charles River Breeding Laboratories. They were inoculated intramuscularly in the hind legs. The same formulated vaccines with the pig inoculation were administered as half dose (0.5 mL) with VLP 1 μg (group 1), VLP 5 μg (group 2), VLP 10 μg (group 3) and VLP 30 μg (group 4). Boosting vaccination was performed with the same administration 14 days after priming vaccination. Blood samples were collected 14 days after second vaccination.
2.4. Serum Antibody Test
2.5. Clinical, Virological and Pathological Evaluation
2.6. Statistical Analysis
3. Results
3.1. VLP Was Purified and Quantified
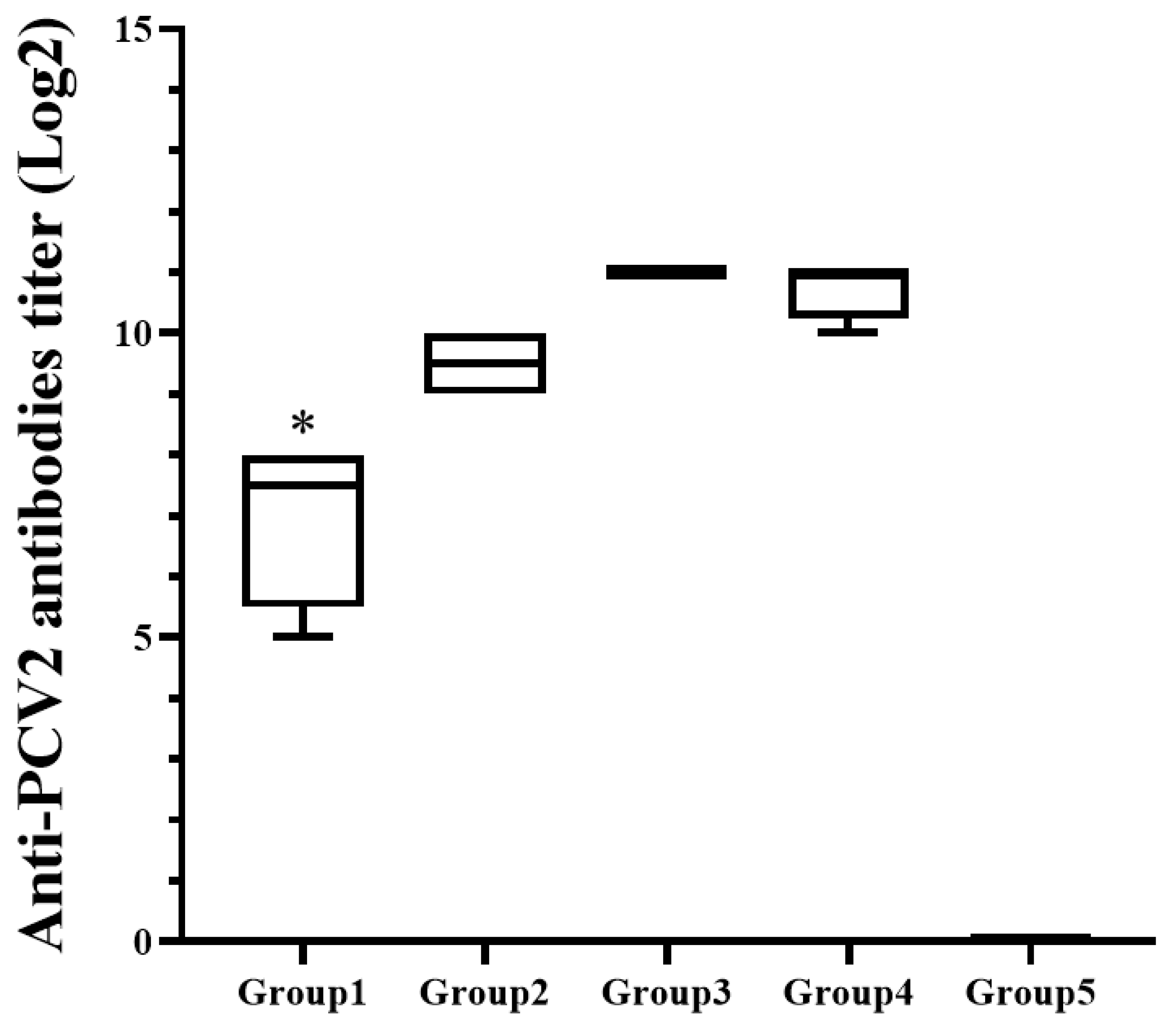
3.2. The Total Antibodies Showed Significant Difference among the Inoculated Guinea Pig Groups
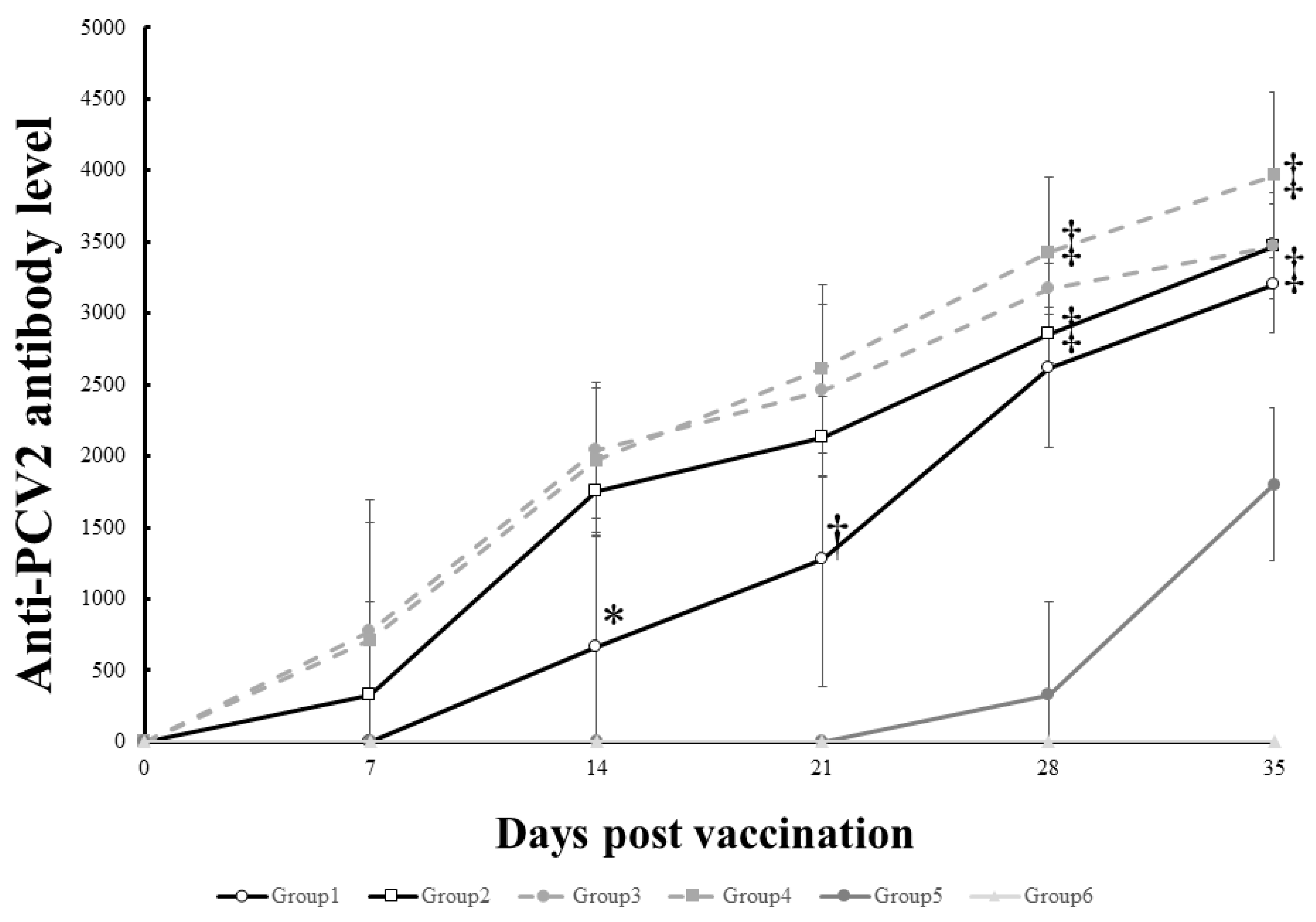
3.3. The Antibody Level of Vaccinated Pigs Was Significantly Higher Than That of Unvaccinated Controls
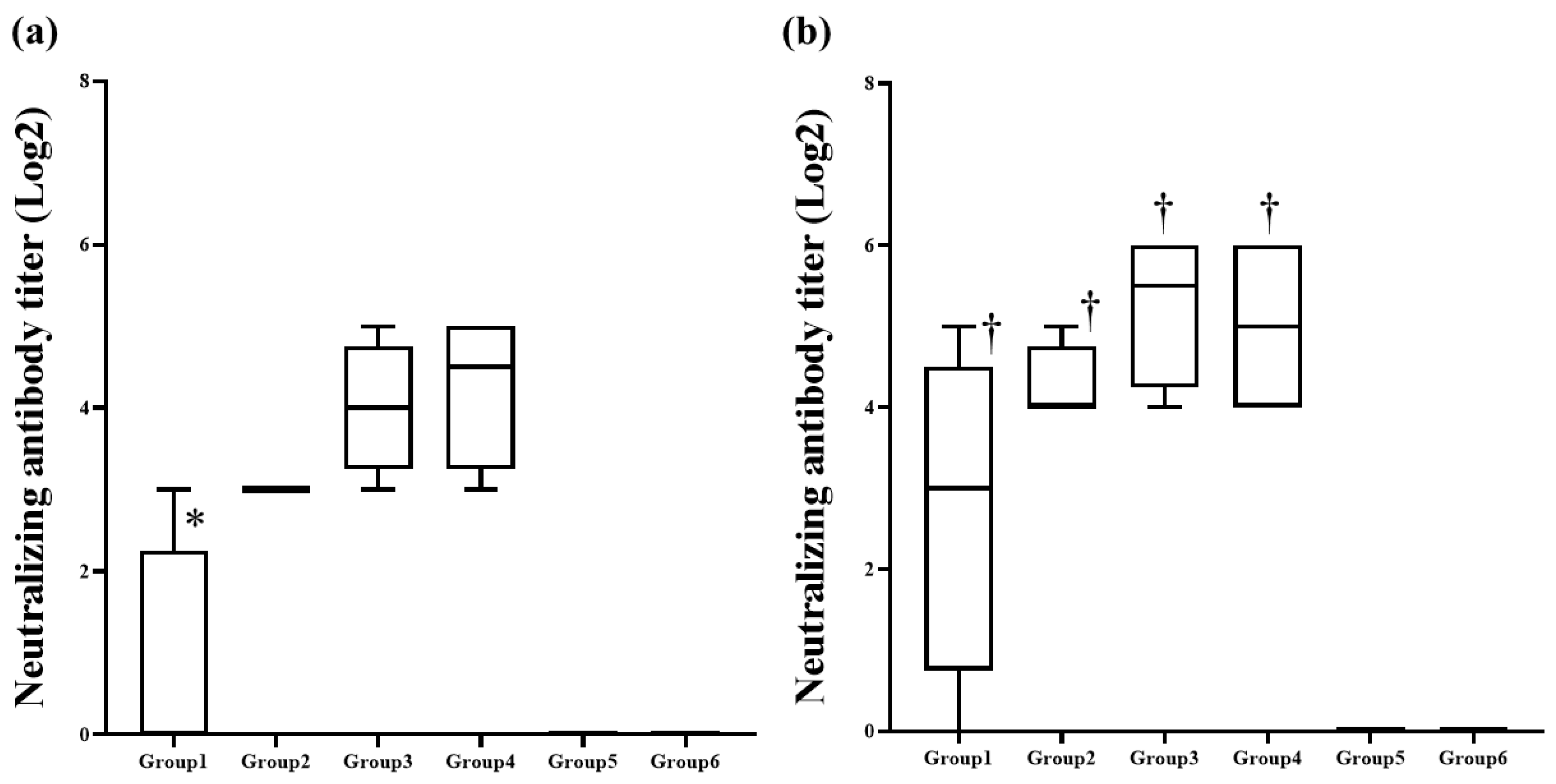
3.4. The Neutralizing Antibodies Were Stably Detected in the Groups Inoculated with More Than 10 μg VLP
3.5. There Was No Difference in Average Daily Weight Gain (ADWG) between the Pigs Challenged with the Virulent PCV2d Isolate Regardless of Vaccination
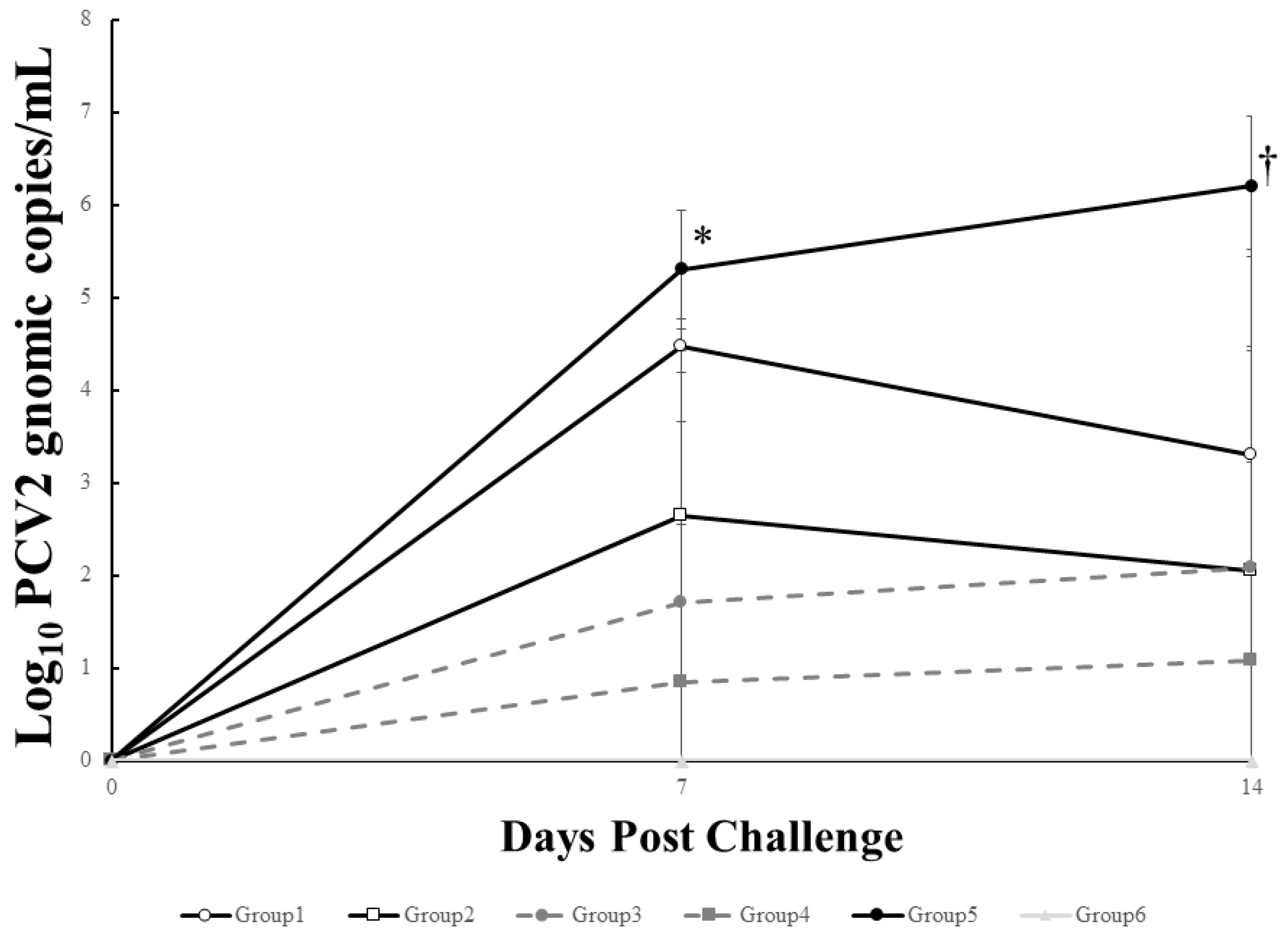
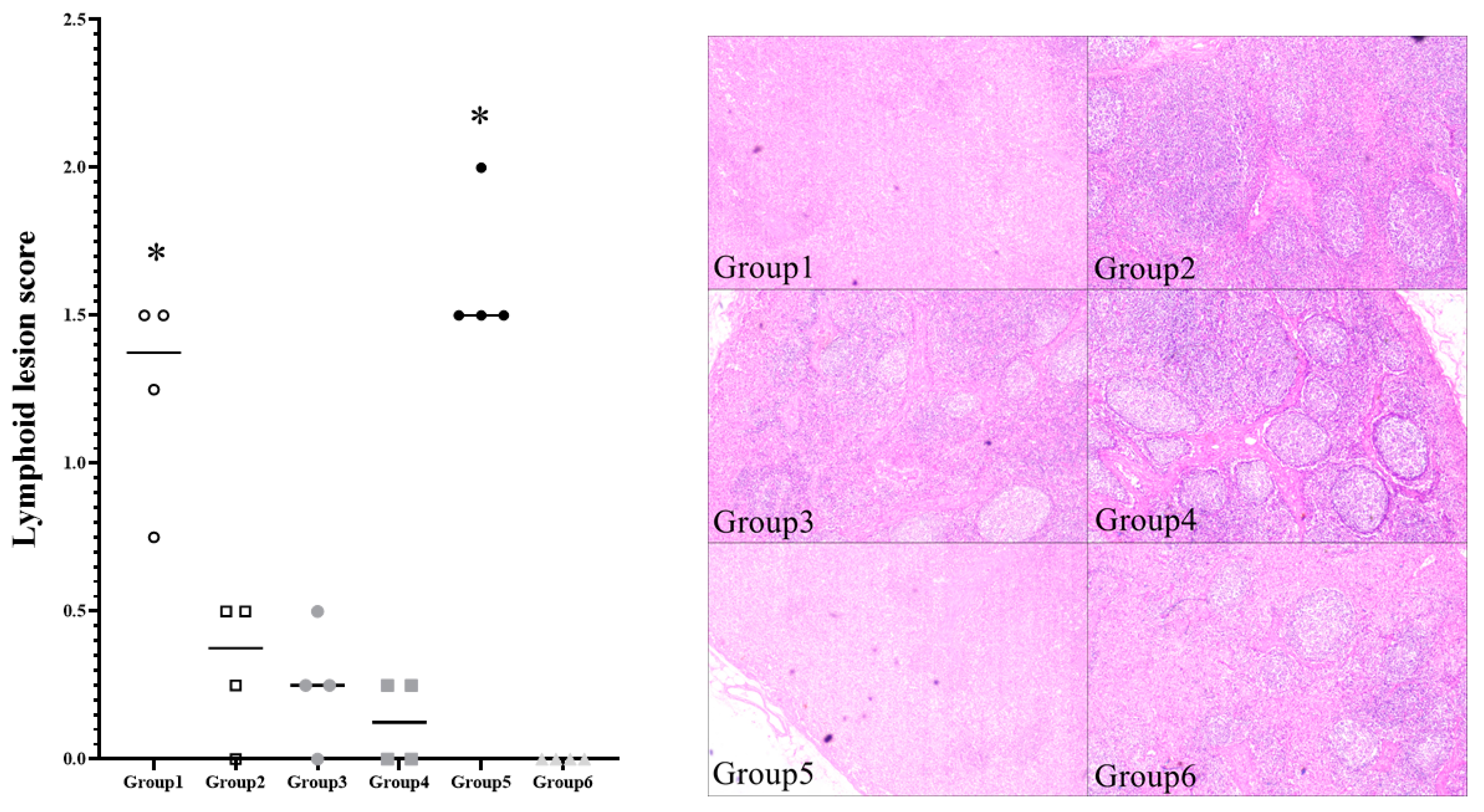
3.6. Viremia and Pathologic Lesion Score of Vaccinated Pigs Were Significantly Lower Than That of Non-Vaccinated Controls
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Opriessnig, T.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 associated disease: Update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Investig. 2007, 19, 591–615. [Google Scholar] [CrossRef]
- Segalés, J.; Allan, G.M.; Domingo, M. Porcine circovirus diseases. Anim. Health Res. Rev. 2005, 6, 119–142. [Google Scholar] [CrossRef]
- Jacobsen, B.; Krueger, L.; Seeliger, F.; Bruegmann, M.; Segalés, J.; Baumgaertner, W. Retrospective study on the occurrence of porcine circovirus 2 infection and associated entities in Northern Germany. Vet. Microbiol. 2009, 138, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Drolet, R.; Thibault, S.; D’Allaire, S.; Thomson, J.R.; Done, S.H. Porcine dermatitis and nephropathy syndrome (PDNS): An overview of the disease. J. Swine Health Prod. 1999, 7, 283–285. [Google Scholar]
- Segalés, J.; Piella, J.; Marco, E.; de-Mateu-Antonio, E.M.; Espuna, E.; Domingo, M. Porcine dermatitis and nephropathy syndrome in Spain. Vet. Rec. 1998, 142, 483–486. [Google Scholar] [CrossRef]
- Segalés, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Franzo, G.; Cortey, M.; Segalés, J.; Hughes, J.; Drigo, M. Phylodynamic analysis of porcine circovirus type 2 reveals global waves of emerging genotypes and the circulation of recombinant forms. Mol. Phylogenet. Evol. 2016, 100, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firth, C.; Charleston, M.A.; Duffy, S.; Shapiro, B.; Holmes, E.C. Insights into the Evolutionary History of an Emerging Livestock Pathogen: Porcine Circovirus 2. J. Virol. 2009, 83, 12813–12821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenaux, M.; Halbur, P.G.; Gill, M.; Toth, T.E.; Meng, X.J. Genetic characterization of type 2 porcine circovirus (PCV-2) from pigs with postweaning multisystemic wasting syndrome in different geographic regions of North America and development of a differential PCR-restriction fragment length polymorphism assay to detect and differentiate between infections with PCV-1 and PCV-2. J. Clin Microbiol. 2000, 38, 2494–2503. [Google Scholar]
- Larochelle, R.; Magar, R.; D’Allaire, S. Genetic characterization and phylogenetic analysis of porcine circovirus type 2 (PCV2) strains from cases presenting various clinical conditions. Virus Res. 2002, 90, 101–112. [Google Scholar] [CrossRef]
- Cheung, A.K.; Lager, K.M.; Kohutyuk, O.I.; Vincent, A.L.; Henry, S.C.; Baker, R.B.; Rowland, R.R.; Dunham, A.G. Detection of two porcine circovirus type 2 genotypic groups in United States swine herds. Arch. Virol. 2007, 152, 1035–1044. [Google Scholar] [CrossRef]
- Gagnon, C.A.; Tremblay, D.; Tijssen, P.; Venne, M.H.; Houde, A.; Elahi, S.M. The emergence of porcine circovirus 2b genotype (PCV-2b) in swine in Canada. Can. Vet. J. 2007, 48, 811–819. [Google Scholar] [PubMed]
- Dupont, K.; Nielsen, E.O.; Bækbo, P.; Larsen, L.E. Genomic analysis of PCV2 isolates from Danish archives and a current PMWS case–control study supports a shift in genotypes with time. Vet. Microbiol. 2008, 128, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J. Gen. Virol. 2015, 96, 1830–1841. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Xiao, C.T.; Halbur, P.G.; Gerber, P.F.; Matzinger, S.R.; Meng, X.J. A commercial porcine circovirus (PCV) type 2a-based vaccine reduces PCV2d viremia and shedding and prevents PCV2d transmission to naïve pigs under experimental conditions. Vaccine 2017, 35, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.W.; Park, C.; Kang, I.; Choi, K.; Jeong, J.; Park, S.J.; Chae, C. Genetic and antigenic characterization of a newly emerging porcine circovirus type 2b mutant first isolated in cases of vaccine failure in Korea. Arch. Virol. 2014, 159, 3107–3111. [Google Scholar] [CrossRef]
- Chae, C. Commercial porcine circovirus type 2 vaccines: Efficacy and clinical application. Vet. J. 2012, 194, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Gerber, P.F.; Xiao, C.-T.; Mogler, M.; Halbur, P.G. A commercial vaccine based on PCV2a and an experimental vaccine based on a variant mPCV2b are both effective in protecting pigs against challenge with a 2013 U.S. variant mPCV2b strain. Vaccine 2014, 32, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.; Andraud, M.; Bigault, L.; Jestin, A.; Grasland, B. A commercial PCV2a-based vaccine significantly reduces PCV2b transmission in experimental conditions. Vaccine 2016, 34, 3738–3745. [Google Scholar] [CrossRef]
- Fort, M.; Sibila, M.; Allepuz, A.; Mateu, E.; Roerink, F.; Segales, J. Porcine circovirus type 2 (PCV2) vaccination of conventional pigs prevents viremia against PCV2 isolates of different genotypes and geographic origins. Vaccine 2008, 26, 1063–1071. [Google Scholar] [CrossRef]
- Cimica, V.; Galarza, J.M. Adjuvant formulations for virus-like particle (VLP) based vaccines. Clin. Immunol. 2017, 183, 99–108. [Google Scholar] [CrossRef]
- Vicente, T.; Roldão, A.; Peixoto, C.; Carrondo, M.J.T.; Alves, P.M. Large-scale production and purification of VLP-based vaccines. J. Invertebr. Pathol. 2011, 107, S42–S48. [Google Scholar] [CrossRef]
- Khurana, S.; Wu, J.; Verma, N.; Verma, S.; Raghunandan, R.; Manischewitz, J.; King, L.R.; Kpamegan, E.; Pincus, S.; Smith, G.; et al. H5N1 virus-like particle vaccine elicits cross-reactive neutralizing antibodies that preferentially bind to the oligomeric form of influenza virus hemagglutinin in humans. J. Virol. 2011, 85, 10945–10954. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, M.F.; Zinkernagel, R.M. Neutralizing antiviral B cell responses. Annu. Rev. Immunol. 1997, 15, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Meerts, P.; Misinzo, G.; Lefebvre, D.; Nielsen, J.; Botner, A.; Kristensen, C.S.; Nauwynck, H.J. Correlation between the presence of neutralizing antibodies against porcine circovirus 2 (PCV2) and protection against replication of the virus and development of PCV2-associated disease. BMC Vet. Res. 2006, 2, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Park, C.; Cho, S.H.; Chung, J. The level of decoy epitope in PCV2 vaccine affects the neutralizing activity of sera in the immunized animals. Biochem. Biophys. Res. Commun. 2018, 496, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Fort, M.; Olvera, A.; Sibila, M.; Segalés, J.; Mateu, E. Detection of neutralizing antibodies in postweaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs. Vet. Microbiol. 2007, 125, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Pogranichnyy, R.M.; Yoon, K.J.; Harms, P.A.; Swenson, S.L.; Zimmerman, J.J.; Sorden, S.D. Characterization of immune response of young pigs to porcine circovirus type 2 infection. Viral Immunol. 2000, 13, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Han, K.; Oh, Y.; Park, C.; Chae, C. Efficacy of a reformulated inactivated chimeric PCV1-2 vaccine based on clinical, virological, pathological and immunological examination under field conditions. Vaccine 2012, 30, 6671–6677. [Google Scholar] [CrossRef]
- Gagnon, C.A.; del Castillo, J.R.; Music, N.; Fontaine, G.; Harel, J.; Tremblay, D. Development and use of a multiplex real-time quantitative polymerase chain reaction assay for detection and differentiation of Porcine circovirus-2 genotypes 2a and 2b in an epidemiological survey. J. Vet. Diagn. Investig. 2008, 20, 545–558. [Google Scholar] [CrossRef] [Green Version]
- Fenaux, M.; Halbur, P.G.; Haqshenas, G. Cloned genomic DNA of type 2 porcine circovirus is infectious when injected directly into the liver and lymph nodes of pigs: Characterization of clinical disease, virus distribution, and pathologic lesions. J. Virol. 2002, 76, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Chae, C. A comparison of virus isolation, polymerase chain reaction, immunohistochemistry, and in situ hybridization for the detection of porcine circovirus 2 and porcine parvovirus in experimentally and naturally coinfected pigs. J. Vet. Diagn. Investig. 2004, 16, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.; Park, C.; Choi, K.; Chae, C. Comparison of three commercial one-dose porcine circovirus type 2 (PCV2) vaccines in a herd with concurrent circulation of PCV2b and mutant PCV2b. Vet. Microbiol. 2015, 177, 43–52. [Google Scholar] [CrossRef]
- Rodriguez, A.G.M.; Segales, J.; Rosell, C.; Quintana, J.; Ayllon, S.; Camprodon, A.; Domingo, M. Aujeszky’s disease virus infection concurrent with postweaning multisystemic wasting syndrome in pigs. Vet. Rec. 1999, 144, 152–153. [Google Scholar] [CrossRef]
- Perez-Martin, E.; Gomez-Sebastian, S.; Argilaguet, J.M.; Sibila, M.; Fort, M.; Nofrarias, M.; Kurtz, S.; Escribano, J.M.; Segalés, J.; Rodríguez, F. Immunity conferred by an experimental vaccine based on the recombinant PCV2 Cap protein expressed in Trichoplusia ni-larvae. Vaccine 2010, 28, 2340–2349. [Google Scholar] [CrossRef]
- Fenaux, M.; Opriessnig, T.; Halbur, P.G.; Elvinger, F.; Meng, X.J. A chimeric porcine circovirus (PCV) with the immunogenic capsid gene of the pathogenic PCV type 2 (PCV2) cloned into the genomic backbone of the nonpathogenic PCV1 induces protective immunity against PCV2 infection in pigs. J. Virol. 2004, 78, 6297–6303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, P.; Ferrari, L.; Morganti, M.; De Angelis, E.; Bonilauri, P.; Guazzetti, S.; Caleffi, A.; Borghetti, P. One dose of a porcine circovirus 2 subunit vaccine induces humoral and cell-mediated immunity and protects against porcine circovirus-associated disease under field conditions. Vet. Microbiol. 2011, 149, 339–351. [Google Scholar] [CrossRef] [Green Version]
- Plummer, E.M.; Manchester, M. Viral nanoparticles and virus-like particles: Platforms for contemporary vaccine design. Wiley Interdiscip. Rev Nanomed. Nanobiotechnol. 2011, 3, 174–196. [Google Scholar] [CrossRef]
- Murata, K.; Lechmann, M.; Qiao, M.; Gunji, T.; Alter, H.; Liang, T.J. Immunization with hepatitis C virus-like particles protects mice from recombinant hepatitis C virus-vaccinia infection. Proc. Natl. Acad. Sci. USA 2003, 100, 6753–6758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paliard, X.; Liu, Y.; Wagner, R.; Wolf, H.; Baenziger, J.; Walker, C.M. Priming of strong, broad, and long-lived HIV type 1 p55gag-specific CD8+ cytotoxic T cells after administration of a virus-like particle vaccine in rhesus macaques. AIDS Res. Hum. Retrovir. 2000, 16, 273–282. [Google Scholar] [CrossRef]
- Tissot, A.C.; Renhofa, R.; Schmitz, N.; Cielens, I.; Meijerink, E.; Ose, V.; Jennings, G.T.; Saudan, P.; Pumpens, P.; Bachmann, M.F. Versatile virus-like particle carrier for epitope based vaccines. PLoS ONE 2010, 5, e9809. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Guinea Pig Experiment | ||||||
| Group 1 | Group 2 | Group 3 | Group 4 | Group 5 | ||
| 1st vaccination | 1 μg VLP | 5 μg VLP | 10 μg VLP | 30 μg VLP | None | |
| 2nd vaccination | 1 μg VLP | 5 μg VLP | 10 μg VLP | 30 μg VLP | None | |
| Pig Challenge Experiment | ||||||
| Group 1 | Group 2 | Group 3 | Group 4 | Group 5 | Group 6 | |
| Vaccination | 2 μg VLP | 10 μg VLP | 20 μg VLP | 60 μg VLP | None | None |
| Challenge | PCV2d | PCV2d | PCV2d | PCV2d | PCV2d | None |
| Pig Challenge Experiment | ||||||
|---|---|---|---|---|---|---|
| Group 1 | Group 2 | Group 3 | Group 4 | Group 5 | Group 6 | |
| 0~7 dpv | ||||||
| ADWG | 132 ± 21 | 442 ± 141 | 289 ± 374 | 157 ± 141 | 267 ± 133 | 310 ± 118 |
| 7~14 dpv | ||||||
| ADWG | 375 ± 86 | 414 ± 117 | 389 ± 102 | 428 ± 76 | 425 ± 114 | 464 ± 37 |
| 14~21 dpv | ||||||
| ADWG | 560 ± 101 | 410 ± 155 | 414 ± 192 | 621 ± 99 | 467 ± 44 | 582 ± 105 |
| 21~28 dpv | ||||||
| ADWG | 432 ± 147 | 653 ± 81 | 582 ± 119 | 553 ± 108 | 471 ± 144 | 564 ± 66 |
| 28~35 dpv | ||||||
| ADWG | 703 ± 131 | 514 ± 182 | 567 ± 97 | 635 ± 158 | 492 ± 245 | 621 ± 170 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, J.-H.; Cha, S.-H.; Cho, S.-H.; Lee, M.-S.; Park, C. Minimal Dosage of Porcine Circovirus Type 2d Based Virus-like Particles to Induce Stable Protective Immunity against Infection. Pathogens 2021, 10, 1644. https://doi.org/10.3390/pathogens10121644
Baek J-H, Cha S-H, Cho S-H, Lee M-S, Park C. Minimal Dosage of Porcine Circovirus Type 2d Based Virus-like Particles to Induce Stable Protective Immunity against Infection. Pathogens. 2021; 10(12):1644. https://doi.org/10.3390/pathogens10121644
Chicago/Turabian StyleBaek, Jong-Hyuk, Sang-Ho Cha, Sun-Hee Cho, Myung-Shin Lee, and Changhoon Park. 2021. "Minimal Dosage of Porcine Circovirus Type 2d Based Virus-like Particles to Induce Stable Protective Immunity against Infection" Pathogens 10, no. 12: 1644. https://doi.org/10.3390/pathogens10121644
APA StyleBaek, J. -H., Cha, S. -H., Cho, S. -H., Lee, M. -S., & Park, C. (2021). Minimal Dosage of Porcine Circovirus Type 2d Based Virus-like Particles to Induce Stable Protective Immunity against Infection. Pathogens, 10(12), 1644. https://doi.org/10.3390/pathogens10121644