
Porcine Circovirus (PCV) Genotype 2d-Based Virus-like Particles (VLPs) Induced Broad Cross-Neutralizing Antibodies against Diverse Genotypes and Provided Protection in Dual-Challenge Infection of a PCV2d Virus and a Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV)

Abstract
:1. Introduction
2. Results
2.1. Production of VLPs Expressed by Baculovirus Expression System
2.2. Cross-Neutralization by VLP-Induced Antisera
2.3. Protection Efficacy of rPCV2d Vaccination in Pigs
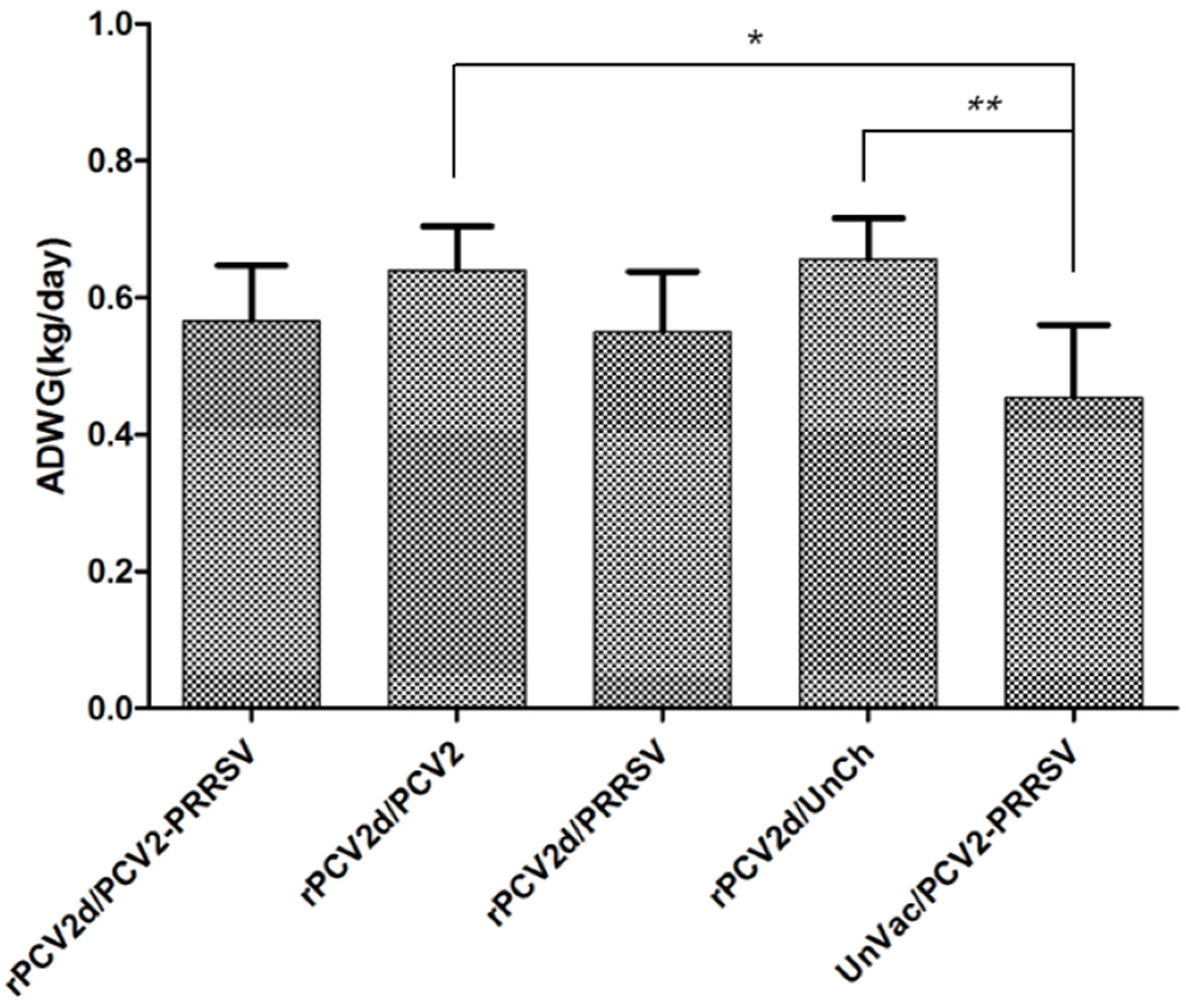
2.3.1. Clinical Signs and ADWG
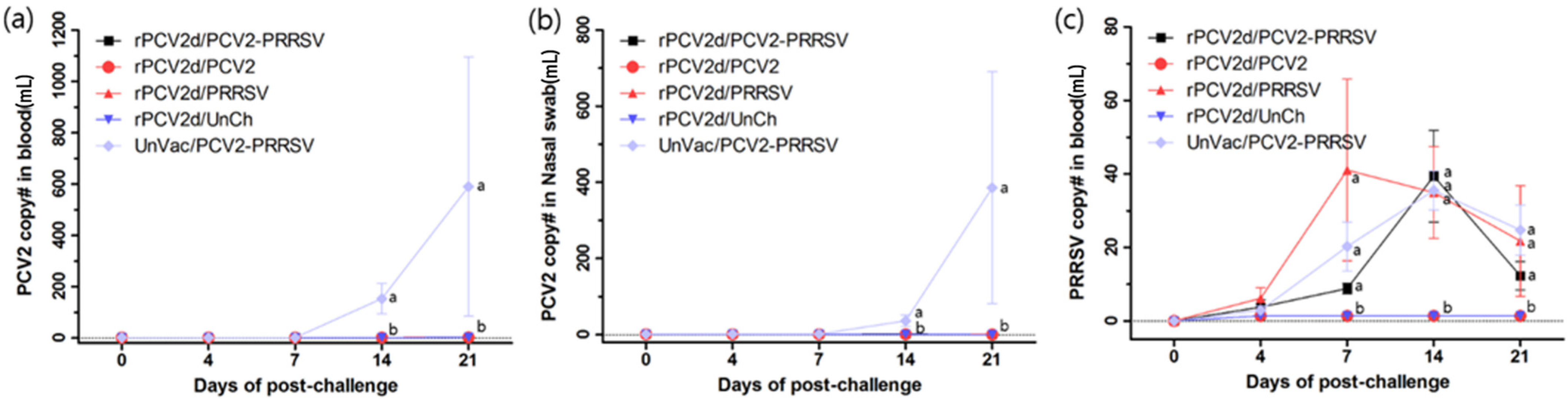
2.3.2. Viral Load of PCV2 and PRRSV in Blood, Nasal Swab and Tissues
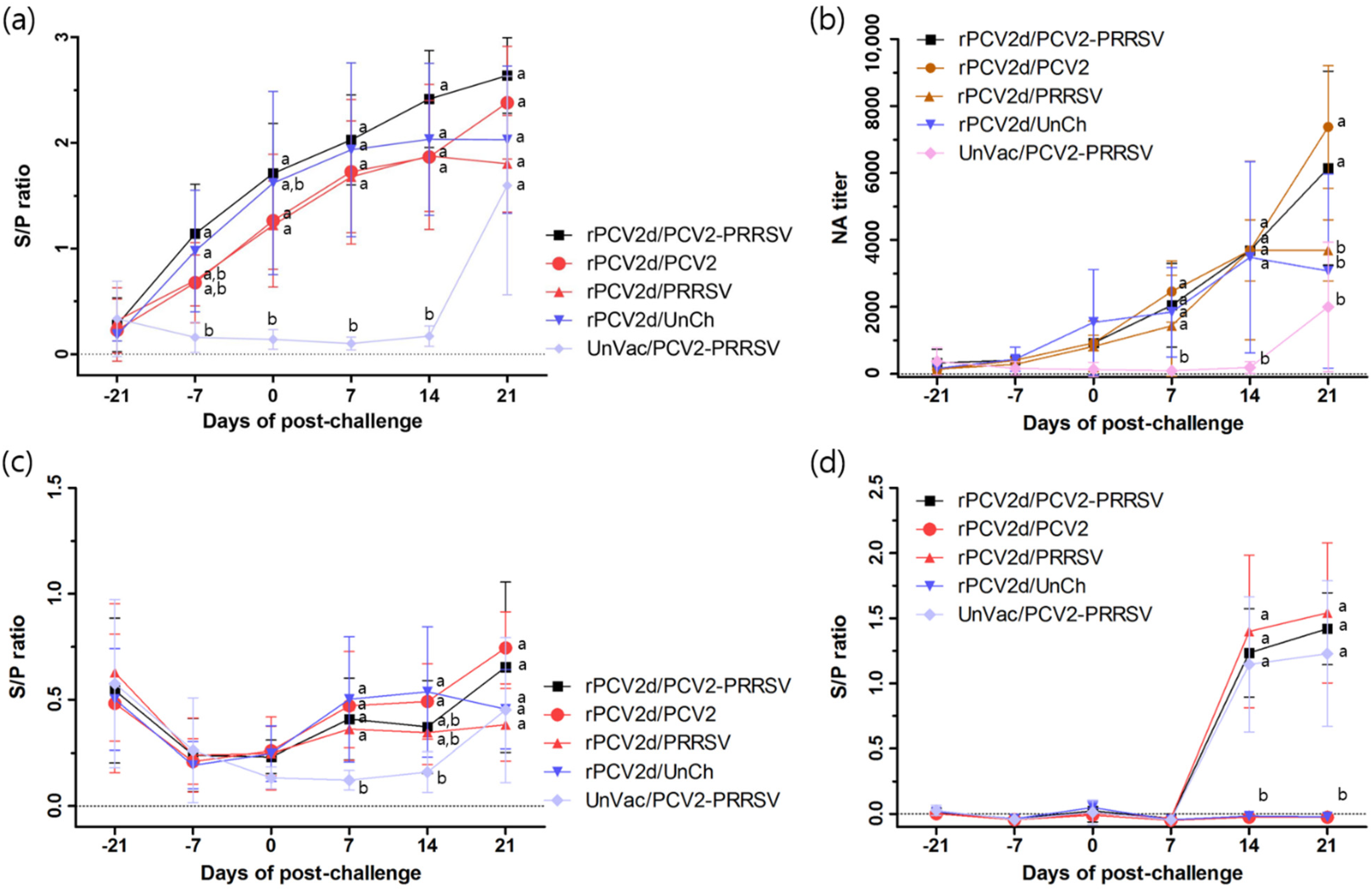
2.3.3. Serology of PCV2 and PRRSV
2.3.4. Gross and Histopathological Lesions
3. Discussion
4. Materials and Methods
4.1. Viruses and Cells
4.2. Production of Recombinant VLP Using Baculovirus Expression System (BES)
4.3. Analysis of PCV2 Virus-like Particles (VLPs)
4.4. Cross-Neutralization by rPCV2a, rPCV2b and rPCV2d-Induced Antisera
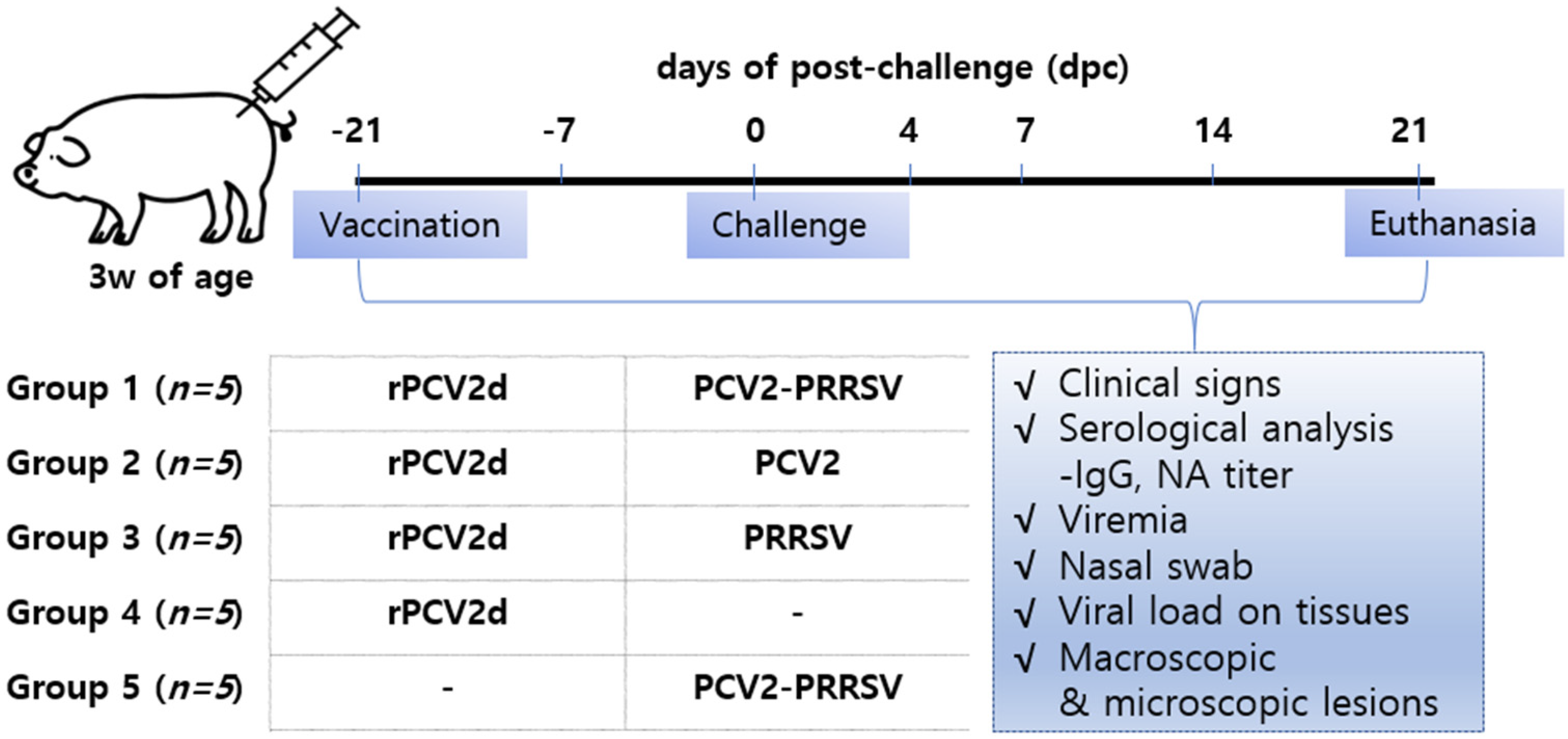
4.5. Protection Efficacy by rPCV2d Vaccination in Pigs
4.5.1. Pig Studies
4.5.2. Clinical Signs and Average Daily Weight Gain (ADWG)
4.5.3. Quantification of PCV2 and PRRSV
4.5.4. ELISA Analysis
4.5.5. Viral Neutralization Assay
4.5.6. Gross Pathology and Histopathology
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, J.; Huang, L.; Wei, Y.; Wang, Y.; Chen, D.; Du, W.; Wu, H.; Liu, C. Prevalence of emerging porcine parvoviruses and their co-infections with porcine circovirus type 2 in China. Arch. Virol. 2015, 160, 1339–1344. [Google Scholar] [CrossRef]
- Wang, H.; Feng, Z.; Wu, Y.; Wei, Y.; Gan, Y.; Hua, L.; Li, B.; Wang, X.; Liu, M.; Xiong, Q.; et al. The effects of Mycoplasma hyopneumoniae on porcine circovirus type 2 replication in vitro PK-15 cells. Res. Vet. Sci. 2016, 105, 56–61. [Google Scholar] [CrossRef]
- Yi, J.; Liu, C. Molecular characterization of porcine circovirus 2 isolated from diseased pigs co-infected with porcine reproductive and respiratory syndrome virus. J. Virol. 2010, 7, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, T.; Lee, D.U.; Yoo, S.J.; Je, S.H.; Shin, J.Y.; Lyoo, Y.S. Genotypic diversity of porcine circovirus type 2 (PCV2) and genotype shift to PCV2d in Korean pig population. Virus Res. 2017, 228, 24–29. [Google Scholar] [CrossRef]
- Kang, S.J.; Kang, H.; You, S.H.; Lee, H.J.; Lee, N.; Hyun, B.H.; Cha, S.H. Genetic diversity and different cross-neutralization capability of porcine circovirus type 2 isolates recently circulating in South Korea. BMC Vet. Res. 2020, 16, 334. [Google Scholar] [CrossRef] [PubMed]
- Tsai, G.T.; Lin, Y.C.; Lin, W.H.; Lin, J.H.; Chiou, M.T.; Liu, H.F.; Lin, C.N. Phylogeographic and genetic characterization of porcine circovirus type 2 in Taiwan from 2001-2017. Sci. Rep. 2019, 9, 10782. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.T.; Harmon, K.M.; Halbur, P.G.; Opriessnig, T. PCV2d-2 is the predominant type of PCV2 DNA in pig samples collected in the U.S. during 2014-2016. Vet. Microbiol. 2016, 197, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Karuppannan, A.K.; Opriessnig, T. Porcine Circovirus Type 2 (PCV2) Vaccines in the Context of Current Molecular Epidemiology. Viruses 2017, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Xiao, C.T.; Halbur, P.G.; Gerber, P.F.; Matzinger, S.R.; Meng, X.J. A commercial porcine circovirus (PCV) type 2a-based vaccine reduces PCV2d viremia and shedding and prevents PCV2d transmission to naive pigs under experimental conditions. Vaccine 2017, 35, 248–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddicks, M.; Fux, R.; Szikora, F.; Eddicks, L.; Majzoub-Altweck, M.; Hermanns, W.; Sutter, G.; Palzer, A.; Banholzer, E.; Ritzmann, M. Detection of a new cluster of porcine circovirus type 2b strains in domestic pigs in Germany. Vet. Microbiol. 2015, 176, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.L.; Vidigal, P.M.; de Souza, L.F.; Onofre, T.S.; Gonzaga, N.F.; Eller, M.R.; Bressan, G.C.; Fietto, J.L.; Almeida, M.R.; Silva Junior, A. Identification of an Emergent Porcine Circovirus-2 in Vaccinated Pigs from a Brazilian Farm during a Postweaning Multisystemic Wasting Syndrome Outbreak. Genome Announc. 2014, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.W.; Park, C.; Kang, I.; Choi, K.; Jeong, J.; Park, S.J.; Chae, C. Genetic and antigenic characterization of a newly emerging porcine circovirus type 2b mutant first isolated in cases of vaccine failure in Korea. Arch. Virol. 2014, 159, 3107–3111. [Google Scholar] [CrossRef]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Complete genome sequence of a novel porcine circovirus type 2b variant present in cases of vaccine failures in the United States. J. Virol. 2012, 86, 12469. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Soria, S.; Sibila, M.; Nofrarias, M.; Calsamiglia, M.; Manzanilla, E.G.; Ramirez-Mendoza, H.; Minguez, A.; Serrano, J.M.; Marin, O.; Joisel, F.; et al. Effect of porcine circovirus type 2 (PCV2) load in serum on average daily weight gain during the postweaning period. Vet. Microbiol. 2014, 174, 296–301. [Google Scholar] [CrossRef]
- Lopez-Sagaseta, J.; Malito, E.; Rappuoli, R.; Bottomley, M.J. Self-assembling protein nanoparticles in the design of vaccines. Comput. Struct. Biotechnol. J. 2016, 14, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Crisci, E.; Barcena, J.; Montoya, M. Virus-like particles: The new frontier of vaccines for animal viral infections. Vet. Immunol. Immunopathol. 2012, 148, 211–225. [Google Scholar] [CrossRef]
- Tegerstedt, K.; Franzen, A.V.; Andreasson, K.; Joneberg, J.; Heidari, S.; Ramqvist, T.; Dalianis, T. Murine polyomavirus virus-like particles (VLPs) as vectors for gene and immune therapy and vaccines against viral infections and cancer. Anticancer. Res. 2005, 25, 2601–2608. [Google Scholar]
- Liu, F.; Wu, X.; Li, L.; Ge, S.; Liu, Z.; Wang, Z. Virus-like particles: Promising platforms with characteristics of DIVA for veterinary vaccine design. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Roldao, A.; Mellado, M.C.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.M. Recombinant protein vaccines produced in insect cells. Vaccine 2012, 30, 1759–1766. [Google Scholar] [CrossRef]
- Huan, C.; Fan, M.; Cheng, Q.; Wang, X.; Gao, Q.; Wang, W.; Gao, S.; Liu, X. Evaluation of the Efficacy and Cross-Protective Immunity of Live-Attenuated Chimeric PCV1-2b Vaccine Against PCV2b and PCV2d Subtype Challenge in Pigs. Front. Microbiol. 2018, 9, 455. [Google Scholar] [CrossRef] [Green Version]
- Allan, G.M.; McNeilly, F.; Ellis, J.; Krakowka, S.; Meehan, B.; McNair, I.; Walker, I.; Kennedy, S. Experimental infection of colostrum deprived piglets with porcine circovirus 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) potentiates PCV2 replication. Arch. Virol. 2000, 145, 2421–2429. [Google Scholar] [CrossRef]
- Opriessnig, T.; Ramamoorthy, S.; Madson, D.M.; Patterson, A.R.; Pal, N.; Carman, S.; Meng, X.J.; Halbur, P.G. Differences in virulence among porcine circovirus type 2 isolates are unrelated to cluster type 2a or 2b and prior infection provides heterologous protection. J. Gen. Virol. 2008, 89, 2482–2491. [Google Scholar] [CrossRef] [PubMed]
- Rovira, A.; Balasch, M.; Segales, J.; Garcia, L.; Plana-Duran, J.; Rosell, C.; Ellerbrok, H.; Mankertz, A.; Domingo, M. Experimental inoculation of conventional pigs with porcine reproductive and respiratory syndrome virus and porcine circovirus 2. J. Virol. 2002, 76, 3232–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fort, M.; Olvera, A.; Sibila, M.; Segales, J.; Mateu, E. Detection of neutralizing antibodies in postweaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs. Vet. Microbiol. 2007, 125, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Hahn, T.W. Evaluation of novel recombinant porcine circovirus type 2d (PCV2d) vaccine in pigs naturally infected with PCV2d. Vaccine 2021, 39, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Oh, Y.; Seo, H.W.; Han, K.; Chae, C. Comparative effects of vaccination against porcine circovirus type 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) in a PCV2-PRRSV challenge model. Clin. Vaccine Immunol. 2013, 20, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, A.; Milek, D.; Matyba, P.; Stadejek, T. Real-Time PCR Detection Patterns of Porcine Circovirus Type 2 (PCV2) in Polish Farms with Different Statuses of Vaccination against PCV2. Viruses 2019, 11, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, F.M.; Silva Junior, A.; Vidigal, P.M.; Oliveira, C.R.; Viana, V.W.; Silva, C.H.; Vargas, M.I.; Fietto, J.L.; Almeida, M.R. Porcine circovirus-2 viral load versus lesions in pigs: Perspectives for post-weaning multisystemic wasting syndrome. J. Comp. Pathol. 2011, 144, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, P.; Mahe, D.; Cariolet, R.; Truong, C.; Le Dimna, M.; Arnauld, C.; Rose, N.; Eveno, E.; Albina, E.; Madec, F.; et al. An ORF2 protein-based ELISA for porcine circovirus type 2 antibodies in post-weaning multisystemic wasting syndrome. Vet. Microbiol. 2003, 94, 183–194. [Google Scholar] [CrossRef]
- Meerts, P.; Van Gucht, S.; Cox, E.; Vandebosch, A.; Nauwynck, H.J. Correlation between type of adaptive immune response against porcine circovirus type 2 and level of virus replication. Viral Immunol. 2005, 18, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Halbur, P.G.; Paul, P.S.; Frey, M.L.; Landgraf, J.; Eernisse, K.; Meng, X.J.; Lum, M.A.; Andrews, J.J.; Rathje, J.A. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 1995, 32, 648–660. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCV2 Isolate (Genotype) | Anti-Mouse Sera | Anti-Pig Sera | ||||
|---|---|---|---|---|---|---|
| rPCV2a | rPCV2b | rPCV2d | rPCV2a | rPCV2b | rPCV2d | |
| QIA215 (2a) | 2867 ± 1351 | 2509 ± 1449 | 4915 ± 1389 | 2047 ± 1023 | 2389 ± 903 | 5632 ± 2560 |
| QIA418 (2b) | 4506 ± 1505 | 2688 ± 1422 | 6553 ± 1003 | 4096 ± 2048 | 4437 ± 2076 | 5802 ± 2389 |
| QIA169 (2d) | 2150 ± 570 | 1075 ± 400 | 5325 ± 1229 | 1024 ± 512 | 3413 ± 683 | 2816 ± 1280 |
| QIA244 (2d) | 64 ± 18 | 58 ± 19 | 819 ± 307 | 75 ± 28 | 149 ± 56 | 725 ± 299 |
| Groups | PCV2 Genomic DNA (per 200 ng Total DNA) | PRRSV Genomic RNA (per 200 ng Total RNA) | ||||||
|---|---|---|---|---|---|---|---|---|
| Lung | Tonsil | Mesenteric LN | Inguinal LN | Lung | Tonsil | MesentericLN | Inguinal LN | |
| 1 | 5.1 ± 1.1 a | 2.8 ± 0.6 a | 0.5 ± 0.3 a | 1.3 ± 0.9 a | 155.3 ± 16.1 a | 117.2 ± 62.0 a | 5.5 ± 0.0 a | 44.8 ± 2.8 a |
| 2 | 6.0 ± 1.6 a | 4.2 ± 0.6 a | 0.4 ± 0.1 a | 1.4 ± 0.8 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b |
| 3 | 1.5 ± 0.1 a | 1.6 ± 0.2 a | 0.1 ± 0.0 a | 0.1 ± 0.0 a | 101.6 ± 11.4 c | 319.1 ± 48.2 c | 2.9 ± 0.0 a | 113.4 ± 1.6 c |
| 4 | 1.3 ± 0.1 a | 2.1 ± 0.7 a | 0.8 ± 0.7 a | 0.1 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b | 0.0 ± 0.0 b |
| 5 | 5237 ± 3380 b | 19,720 ± 11,795 b | 11,172 ± 8150 b | 3742 ± 2488 b | 106.9 ± 14.1 c | 171.4 ± 16.4 a | 25.8 ± 3.2 c | 48.9 ± 7.4 a |
| Groups | Macroscopic Lesions | Microscopic Lesions | ||||||
|---|---|---|---|---|---|---|---|---|
| Lung | Tonsil | Mesenteric LN | Inguinal LN | Lung | Tonsil | Mesenteric LN | Inguinal LN | |
| 1 | 0.4 ± 0.6 a,c | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 a | 0.8 ± 0.5 a | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 a |
| 2 | 0.2 ± 0.5 a | 0.0 ± 0.0 | 0.2 ± 0.5 | 0.2 ± 0.5 a | 0.0 ± 0.0 b | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 a |
| 3 | 1.0 ± 0.0 b,c | 0.0 ± 0.0 | 0.0 ± 0.0 | 1.0 ± 0.0 b | 0.3 ± 0.5 a,b | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 a |
| 4 | 0.0 ± 0.0 a | 0.0 ± 0.0 | 0.2 ± 0.5 | 0.0 ± 0.0 a | 0.0 ± 0.0 b | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 a |
| 5 | 1.2 ± 0.5 b | 0.0 ± 0.0 | 0.4 ± 0.6 | 1.2 ± 0.5 b | 1.4 ± 0.6 c | 0.6 ± 0.9 | 0.4 ± 0.6 | 0.8 ± 0.5 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.-J.; Bae, S.-M.; Lee, H.-J.; Jeong, Y.-J.; Lee, M.-A.; You, S.-H.; Lee, H.-S.; Hyun, B.-H.; Lee, N.; Cha, S.-H. Porcine Circovirus (PCV) Genotype 2d-Based Virus-like Particles (VLPs) Induced Broad Cross-Neutralizing Antibodies against Diverse Genotypes and Provided Protection in Dual-Challenge Infection of a PCV2d Virus and a Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Pathogens 2021, 10, 1145. https://doi.org/10.3390/pathogens10091145
Kang S-J, Bae S-M, Lee H-J, Jeong Y-J, Lee M-A, You S-H, Lee H-S, Hyun B-H, Lee N, Cha S-H. Porcine Circovirus (PCV) Genotype 2d-Based Virus-like Particles (VLPs) Induced Broad Cross-Neutralizing Antibodies against Diverse Genotypes and Provided Protection in Dual-Challenge Infection of a PCV2d Virus and a Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Pathogens. 2021; 10(9):1145. https://doi.org/10.3390/pathogens10091145
Chicago/Turabian StyleKang, Seok-Jin, Sung-Min Bae, Hye-Jeong Lee, Young-Ju Jeong, Min-A Lee, Su-Hwa You, Hyang-Sim Lee, Bang-Hun Hyun, Nakhyung Lee, and Sang-Ho Cha. 2021. "Porcine Circovirus (PCV) Genotype 2d-Based Virus-like Particles (VLPs) Induced Broad Cross-Neutralizing Antibodies against Diverse Genotypes and Provided Protection in Dual-Challenge Infection of a PCV2d Virus and a Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV)" Pathogens 10, no. 9: 1145. https://doi.org/10.3390/pathogens10091145
APA StyleKang, S. -J., Bae, S. -M., Lee, H. -J., Jeong, Y. -J., Lee, M. -A., You, S. -H., Lee, H. -S., Hyun, B. -H., Lee, N., & Cha, S. -H. (2021). Porcine Circovirus (PCV) Genotype 2d-Based Virus-like Particles (VLPs) Induced Broad Cross-Neutralizing Antibodies against Diverse Genotypes and Provided Protection in Dual-Challenge Infection of a PCV2d Virus and a Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Pathogens, 10(9), 1145. https://doi.org/10.3390/pathogens10091145