
Brucella: Reservoirs and Niches in Animals and Humans



{kind=link}
Abstract
:1. Introduction
1.1. Brucellosis, a Zoonosis
1.2. Infection and Dissemination
1.3. Acute and Chronic Infections
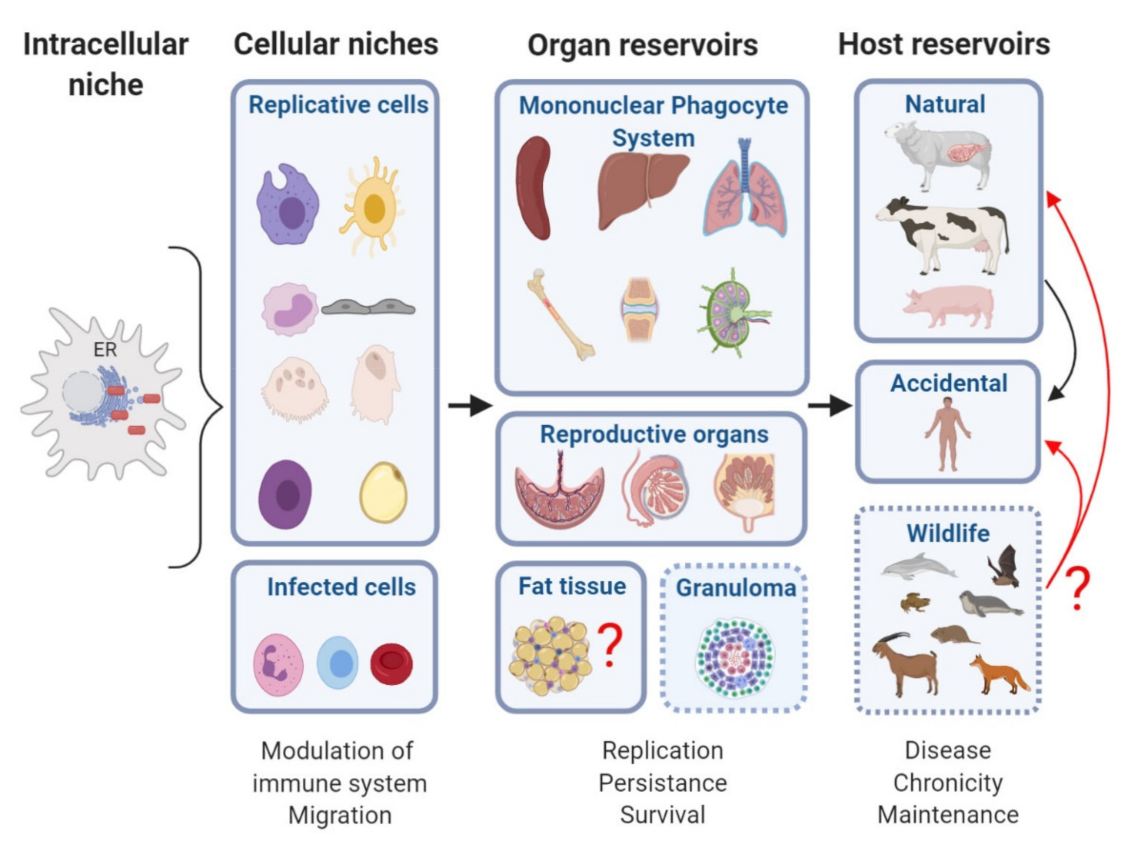
2. Niche and Reservoir
2.1. Definitions
2.2. Intracellular Niche
3. Gold Organs in Brucellosis
3.1. The Reticuloendothelial System
3.2. Genital-Reproductive Organs: Placenta and Epididymis
4. Reservoirs
4.1. Bone Marrow
4.1.1. Bone Marrow in the Mouse Model
4.1.2. Bone Marrow in Humans
4.1.3. Bone Marrow Environment
- (i)
- Colonization of an immune-privileged organ that lacks intracellular antibacterial mechanisms in early-stage stem cells is easier.
- (ii)
- Expression of drug efflux pumps by bone marrow cells facilitates resistance to antibiotic therapy.
- (iii)
- Cells from bone marrow are highly motile and reside in close proximity to arterial vessels thus offering a wide distribution system for dissemination of the pathogen to target organs.
- (iv)
- Brucella’s control of HSC release might explain the splenomegaly observed in human cases as a result of the signal triggered in the bone marrow niche by the apoptotic infected neutrophils to the macrophages, which promotes ensuing cycles of HSC release and extramedullary hematopoiesis.
- (v)
- All Brucella preferential target cells differentiate from progenitor stem cells in the bone marrow.
4.2. Lymph Nodes
4.3. Adipose Tissue
- (i)
- Fat tissues are enriched in immune cells as aforementioned. With respect to macrophage populations, M2 polarized ones together with other immune cells recruited locally, like neutrophils, monocytes, and DC, might help to maintain Brucella survival and take control of the immune response during infection as suggested by the anti-inflammatory polarization of adipose tissue macrophages potentiated by chronic T. cruzi infection [145].
- (ii)
- The majority of drugs are lipophilic compounds, meaning that efficient distribution within the adipose tissue is hampered, as illustrated by some HIV treatment failures. In human brucellosis, 4% of cases undergo to chronic phase [151]. This incidence may rise because of delayed administration of antibiotics or its inefficient delivery to specific organs, such as fat tissue, in the acute phase of infection.
- (iii)
- The localization of fat depots in the perigonadal region of rodents, humans, and ruminants [152] and the preference of Brucella to infect epididymis might not just be a simple coincidence and reflect an essential role of adipose tissue in the persistence of this bacteria in reproductive organs. It also suggests that fat tissue provides Brucella with a proper environment, i.e., a rich source of nutrients.
- (iv)
- FALCs respond dynamically to stimuli. They expand in size and numbers in response to acute or chronic peritoneal insult [153]. More specifically, an increase of B1 cells, macrophages, and neutrophils recruited via the high endothelial venules has been described in omental FALCs upon injection of E. coli LPS or infection [154]. FALCs also support a unique population of CD4+ regulatory cells producers of IL-10 called visceral adipocyte tissue-associated (VAT) Tregs, mostly studied in large fat depots like epididymal fat [155] but also present in the omentum, where it likely regulates local immune responses [156]. Given the essential role of IL-10 in promoting B. abortus persistence and pathology [157] and the balance between activation and regulation of immune state that exists in adipocyte tissue, it is conceivable that pathogens, and Brucella, in particular, find a golden reservoir here, in which regulatory cells would control the local immune response and create a tolerogenic environment for progression to chronicity.
5. Reservoirs in Wildlife
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fouskis, I.; Sandalakis, V.; Christidou, A.; Tsatsaris, A.; Tzanakis, N.; Tselentis, Y.; Psaroulaki, A. The epidemiology of Brucellosis in Greece, 2007–2012: A ‘One Health’ approach. Trans. R. Soc. Trop. Med. Hyg. 2018, 112, 124–135. [Google Scholar] [CrossRef]
- Guzmán-Hernández, R.L.; Contreras-Rodríguez, A.; Ávila-Calderón, E.D.; Morales-García, M.R. Brucelosis: Zoonosis de importancia en México. Rev. Chil. Infectol. 2016, 33, 656–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Mora, G.; Bonilla-Montoya, R.; Barrantes-Granados, O.; Esquivel-Suárez, A.; Montero-Caballero, D.; González-Barrientos, R.; Fallas-Monge, Z.; Palacios-Alfaro, J.D.; Baldi, M.; Campos, E.; et al. Brucellosis in mammals of Costa Rica: An epidemiological survey. PLoS ONE 2017, 12, e0182644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.; Xie, S.; Lu, X.; Sun, L.; Zhou, Y.; Zhang, Y.; Wang, K. A Systematic Review and Meta-Analysis of Epidemiology and Clinical Manifestations of Human Brucellosis in China. Biomed. Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, M.P.; Mulder, M.; Gilman, R.H.; Smits, H.L. Human brucellosis. Lancet Infect. Dis. 2007, 7, 775–786. [Google Scholar] [CrossRef]
- O’Callaghan, D. Human brucellosis: Recent advances and future challenges. Infect. Dis. Poverty 2020, 9, 101. [Google Scholar] [CrossRef]
- Bardhan, D.; Kumar, S.; Verma, M.R.; Bangar, Y.C. Economic losses due to brucellosis in India. Indian J. Comp. Microbiol. Immunol. Infect. Dis. 2020, 41, 19. [Google Scholar] [CrossRef]
- Zeng, J.Y.; Robertson, I.D.; Ji, Q.M.; Dawa, Y.L.; Bruce, M. Evaluation of the economic impact of brucellosis in domestic yaks of Tibet. Transbound. Emerg. Dis. 2019, 66, 476–487. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Esquivel, M.; Chaves-Olarte, E.; Moreno, E.; Guzmán-Verri, C. Brucella Genomics: Macro and Micro Evolution. Int. J. Mol. Sci. 2020, 21, 7749. [Google Scholar] [CrossRef]
- Foster, G.; Osterman, B.S.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti sp. nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [Google Scholar] [CrossRef] [Green Version]
- Jaÿ, M.; Freddi, L.; Mick, V.; Durand, B.; Girault, G.; Perrot, L.; Taunay, B.; Vuilmet, T.; Azam, D.; Ponsart, C.; et al. Brucella microti-like prevalence in French farms producing frogs. Transbound. Emerg. Dis. 2020, 67, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Scholz, H.C.; Nöckler, K.; Göllner, C.; Bahn, P.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Kämpfer, P.; Cloeckaert, A.; Maquart, M.; et al. Brucella inopinata sp. nov., isolated from a breast implant infection. Int. J. Syst. Evol. Microbiol. 2010, 60, 801–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, H.C.; Revilla-Fernández, S.; Dahouk, S.A.; Hammerl, J.A.; Zygmunt, M.S.; Cloeckaert, A.; Koylass, M.; Whatmore, A.M.; Blom, J.; Vergnaud, G.; et al. Brucella vulpis sp. nov., isolated from mandibular lymph nodes of red foxes (Vulpes vulpes). Int. J. Syst. Evol. Microbiol. 2016, 66, 2090–2098. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Esquivel, M.; Baker, K.S.; Ruiz-Villalobos, N.; Hernández-Mora, G.; Barquero-Calvo, E.; González-Barrientos, R.; Castillo-Zeledón, A.; Jiménez-Rojas, C.; Chacón-Díaz, C.; Cloeckaert, A.; et al. Brucella Genetic Variability in Wildlife Marine Mammals Populations Relates to Host Preference and Ocean Distribution. Genome Biol. Evol. 2017, 9, 1901–1912. [Google Scholar] [CrossRef]
- Whatmore, A.M.; Davison, N.; Cloeckaert, A.; Al Dahouk, S.; Zygmunt, M.S.; Brew, S.D.; Perrett, L.L.; Koylass, M.S.; Vergnaud, G.; Quance, C.; et al. Brucella papionis sp. nov., isolated from baboons (Papio spp.). Int. J. Syst. Evol. Microbiol. 2014, 64, 4120–4128. [Google Scholar] [CrossRef] [Green Version]
- Whatmore, A.M.; Dale, E.J.; Stubberfield, E.; Muchowski, J.; Koylass, M.; Dawson, C.; Gopaul, K.K.; Perrett, L.L.; Jones, M.; Lawrie, A. Isolation of Brucella from a White’s tree frog (Litoria caerulea). JMM Case Rep. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Godfroid, J.; Al Dahouk, S.; Pappas, G.; Roth, F.; Matope, G.; Muma, J.; Marcotty, T.; Pfeiffer, D.; Skjerve, E. A “One Health” surveillance and control of brucellosis in developing countries: Moving away from improvisation. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Ay, N.; Kaya, S.; Anil, M.; Alp, V.; Beyazit, U.; Yuksel, E.; Danis, R. Pulmonary Involvement in Brucellosis, a Rare Complication of Renal Transplant: Case Report and Brief Review. Exp. Clin. Transplant. Off. J. Middle East. Soc. Organ. Transplant. 2018, 16, 757–760. [Google Scholar] [CrossRef]
- Bosilkovski, M.; Arapović, J.; Keramat, F. Human brucellosis in pregnancy—An overview. Bosn. J. Basic Med. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ting, I.-W.; Ho, M.-W.; Sung, Y.-J.; Tien, N.; Chi, C.-Y.; Ho, H.-C.; Huang, C.-C. Brucellosis in a renal transplant recipient. Transpl. Infect. Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Gondolfo, R.B.; Cerchiari, N. Human-to-human transmission of Brucella—A systematic review. Trop. Med. Int. Health 2017, 22, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Wallach, J.C.; Samartino, L.E.; Efron, A.; Baldi, P.C. Human infection by Brucella melitensis: An outbreak attributed to contact with infected goats. FEMS Immunol. Med. Microbiol. 1997, 19, 315–321. [Google Scholar] [CrossRef]
- Aygen, B.; Doğanay, M.; Sümerkan, B.; Yildiz, O.; Kayabaş, Ü. Clinical manifestations, complications and treatment of brucellosis: A retrospective evaluation of 480 patients. Médecine Mal. Infect. 2002, 32, 485–493. [Google Scholar] [CrossRef]
- Buzgan, T.; Karahocagil, M.K.; Irmak, H.; Baran, A.I.; Karsen, H.; Evirgen, O.; Akdeniz, H. Clinical manifestations and complications in 1028 cases of brucellosis: A retrospective evaluation and review of the literature. Int. J. Infect. Dis. 2010, 14, e469–e478. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.; Splitter, G.A. Molecular Host-Pathogen Interaction in Brucellosis: Current Understanding and Future Approaches to Vaccine Development for Mice and Humans. Clin. Microbiol. Rev. 2003, 16, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Köse, Ş.; Serin Senger, S.; Akkoçlu, G.; Kuzucu, L.; Ulu, Y.; Ersan, G.; Oğuz, F. Clinical manifestations, complications, and treatment of brucellosis: Evaluation of 72 cases. Turk. J. Med. Sci. 2014, 44, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Moriyón, I. The Genus Brucella. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 315–456. [Google Scholar]
- Mcdermott, J.J.; Grace, D.; Zinsstag, J. Economics of brucellosis impact and control in low-income countries. Rev. Sci. Tech. De L’oie 2013, 32, 249–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, S.C.; Palmer, M.V. Advancement of Knowledge of Brucella over the Past 50 Years. Vet. Pathol. 2014, 51, 1076–1089. [Google Scholar] [CrossRef]
- Moreno, E. Retrospective and prospective perspectives on zoonotic brucellosis. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Poester, F.P.; Samartino, L.E.; Santos, R.L. Pathogenesis and pathobiology of brucellosis in livestock. Rev. Sci. Tech. De L’oie 2013, 32, 105–115. [Google Scholar] [CrossRef] [Green Version]
- von Bargen, K.; Gagnaire, A.; Arce-Gorvel, V.; de Bovis, B.; Baudimont, F.; Chasson, L.; Bosilkovski, M.; Papadopoulos, A.; Martirosyan, A.; Henri, S.; et al. Cervical Lymph Nodes as a Selective Niche for Brucella during Oral Infections. PLoS ONE 2015, 10, e0121790. [Google Scholar] [CrossRef] [Green Version]
- Moreno, E.; Barquero-Calvo, E. The Role of Neutrophils in Brucellosis. Microbiol. Mol. Biol. Rev. 2020, 84, e00048-20. [Google Scholar] [CrossRef]
- Anderson, T.D.; Meador, V.P.; Cheville, N.F. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. I. Gross and Histologic Lesions. Vet. Pathol. 1986, 23, 219–226. [Google Scholar] [CrossRef]
- Meador, V.P.; Deyoe, B.L. Intracellular Localization of Brucella abortus in Bovine Placenta. Vet. Pathol. 1989, 26, 513–515. [Google Scholar] [CrossRef]
- Tobias, L.; Cordes, D.O.; Schurig, G.G. Placental Pathology of the Pregnant Mouse Inoculated with Brucella abortus Strain 2308. Vet. Pathol. 1993, 30, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Harmon, B.G.; Adams, L.G.; Frey, M. Survival of rough and smooth strains of Brucella abortus in bovine mammary gland macrophages. Am. J. Vet. Res. 1988, 49, 1092–1097. [Google Scholar] [PubMed]
- Hull, N.C.; Schumaker, B.A. Comparisons of brucellosis between human and veterinary medicine. Infect. Ecol. Epidemiol. 2018, 8, 1500846. [Google Scholar] [CrossRef] [PubMed]
- Amjadi, O.; Rafiei, A.; Mardani, M.; Zafari, P.; Zarifian, A. A review of the immunopathogenesis of Brucellosis. Infect. Dis. 2019, 51, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Pabuccuoglu, O.; Ecemis, T.; El, S.; Coskun, A.; Akcali, S.; Sanlidag, T. Evaluation of serological tests for diagnosis of brucellosis. Jpn. J. Infect. Dis. 2011, 64, 272–276. [Google Scholar] [PubMed]
- Brucellosis in Humans and Animals; Weltgesundheitsorganisation; FAO (Eds.) WHO: Geneva, Switzerland, 2006; p. 89. [Google Scholar]
- Megid, J.; Antonio Mathias, L.; Robles, C. Clinical Manifestations of Brucellosis in Domestic Animals and Humans. Open Vet. Sci. J. 2010, 4, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Barquero-Calvo, E.; Chaves-Olarte, E.; Weiss, D.S.; Guzmán-Verri, C.; Chacón-Díaz, C.; Rucavado, A.; Moriyón, I.; Moreno, E. Brucella abortus Uses a Stealthy Strategy to Avoid Activation of the Innate Immune System during the Onset of Infection. PLoS ONE 2007, 2, e631. [Google Scholar] [CrossRef] [PubMed]
- Martirosyan, A.; Moreno, E.; Gorvel, J.-P. An evolutionary strategy for a stealthy intracellular Brucella pathogen: Brucella pathogenenesis. Immunol. Rev. 2011, 240, 211–234. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Verri, C.; Manterola, L.; Sola-Landa, A.; Parra, A.; Cloeckaert, A.; Garin, J.; Gorvel, J.-P.; Moriyon, I.; Moreno, E.; Lopez-Goni, I. The two-component system BvrR/BvrS essential for Brucella abortus virulence regulates the expression of outer membrane proteins with counterparts in members of the Rhizobiaceae. Proc. Natl. Acad. Sci. USA 2002, 99, 12375–12380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro-Cerdá, J.; Desjardins, M.; Moreno, E.; Akira, S.; Gorvel, J.P. Modulation of endocytosis in nuclear factor IL-6(−/−) macrophages is responsible for a high susceptibility to intracellular bacterial infection. J. Immunol. 1999, 162, 3519–3526. [Google Scholar] [PubMed]
- Gomes, M.T.R.; Campos, P.C.; de Almeida, L.A.; Oliveira, F.S.; Costa, M.M.S.; Marim, F.M.; Pereira, G.S.M.; Oliveira, S.C. The role of innate immune signals in immunity to Brucella abortus. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Polechová, J.; Storch, D. Ecological Niche. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 72–80. [Google Scholar]
- Gorvel, J.P.; Moreno, E. Brucella intracellular life: From invasion to intracellular replication. Vet. Microbiol. 2002, 90, 281–297. [Google Scholar] [CrossRef]
- Godfroid, J.; Garin-Bastuji, B.; Saegerman, C.; Blasco, J.M. Brucellosis in terrestrial wildlife. Rev. Sci. Tech. De L’oie 2013, 32, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Watarai, M.; Suzuki, H.; Makino, S.-i.; Kodama, T.; Shirahata, T. Lipid raft microdomains mediate class A scavenger receptor-dependent infection of Brucella abortus. Microb. Pathog. 2004, 37, 11–19. [Google Scholar] [CrossRef]
- Naroeni, A.; Porte, F. Role of Cholesterol and the Ganglioside GM1 in Entry and Short-Term Survival of Brucella suis in Murine Macrophages. Infect. Immun. 2002, 70, 1640–1644. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ranwez, V.; Posadas, D.M.; Van der Henst, C.; Estein, S.M.; Arocena, G.M.; Abdian, P.L.; Martín, F.A.; Sieira, R.; De Bolle, X.; Zorreguieta, A. BtaE, an adhesin that belongs to the trimeric autotransporter family, is required for full virulence and defines a specific adhesive pole of Brucella suis. Infect. Immun. 2013, 81, 996–1007. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Ranwez, V.; Posadas, D.M.; Estein, S.M.; Abdian, P.L.; Martin, F.A.; Zorreguieta, A. The BtaF Trimeric Autotransporter of Brucella suis Is Involved in Attachment to Various Surfaces, Resistance to Serum and Virulence. PLoS ONE 2013, 8, e79770. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Reynoso, B.; Lapaque, N.; Salcedo, S.; Briones, G.; Ciocchini, A.E.; Ugalde, R.; Moreno, E.; Moriyón, I.; Gorvel, J.-P. Cyclic β-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nat. Immunol. 2005, 6, 618–625. [Google Scholar] [CrossRef]
- Sola-Landa, A.; Pizarro-Cerdá, J.; Grilló, M.J.; Moreno, E.; Moriyón, I.; Blasco, J.M.; Gorvel, J.P.; López-Goñi, I. A two-component regulatory system playing a critical role in plant pathogens and endosymbionts is present in Brucella abortus and controls cell invasion and virulence. Mol. Microbiol. 1998, 29, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Döhmer, P.H.; Valguarnera, E.; Czibener, C.; Ugalde, J.E. Identification of a type IV secretion substrate of B rucella abortus that participates in the early stages of intracellular survival: A novel Brucella virB secretion substrate. Cell. Microbiol. 2014, 16, 396–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Barsy, M.; Jamet, A.; Filopon, D.; Nicolas, C.; Laloux, G.; Rual, J.-F.; Muller, A.; Twizere, J.-C.; Nkengfac, B.; Vandenhaute, J.; et al. Identification of a Brucella spp. secreted effector specifically interacting with human small GTPase Rab2: A Brucella effector interacting with Rab2. Cell. Microbiol. 2011, 13, 1044–1058. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.P.; Cotto-Rosario, A.; Borghesan, E.; Held, K.; Miller, C.N.; Celli, J. Epistatic Interplay between Type IV Secretion Effectors Engages the Small GTPase Rab2 in the Brucella Intracellular Cycle. mBio 2020, 11, e03350-19. [Google Scholar] [CrossRef] [Green Version]
- Celli, J.; de Chastellier, C.; Franchini, D.-M.; Pizarro-Cerda, J.; Moreno, E.; Gorvel, J.-P. Brucella Evades Macrophage Killing via VirB-dependent Sustained Interactions with the Endoplasmic Reticulum. J. Exp. Med. 2003, 198, 545–556. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, D.; Cazevieille, C.; Allardet-Servent, A.; Boschiroli, M.L.; Bourg, G.; Foulongne, V.; Frutos, P.; Kulakov, Y.; Ramuz, M. A homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl type IV secretion systems is essential for intracellular survival of Brucella suis: Brucella type IV secretion. Mol. Microbiol. 2002, 33, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Comerci, D.J.; Martinez-Lorenzo, M.J.; Sieira, R.; Gorvel, J.-P.; Ugalde, R.A. Essential role of the VirB machinery in the maturation of the Brucella abortus-containing vacuole. Cell. Microbiol. 2001, 3, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.C.; Tsolis, R.M.; Ficht, T.A. Identification of Genes Required for Chronic Persistence of Brucella abortus in Mice. Infect. Immun. 2000, 68, 4102–4107. [Google Scholar] [CrossRef] [Green Version]
- Byndloss, M.X.; Tsai, A.Y.; Walker, G.T.; Miller, C.N.; Young, B.M.; English, B.C.; Seyffert, N.; Kerrinnes, T.; de Jong, M.F.; Atluri, V.L.; et al. Brucella abortus Infection of Placental Trophoblasts Triggers Endoplasmic Reticulum Stress-Mediated Cell Death and Fetal Loss via Type IV Secretion System-Dependent Activation of CHOP. mBio 2019, 10, e01538-19. [Google Scholar] [CrossRef] [Green Version]
- Celli, J. The changing nature of the B rucella-containing vacuole: The Brucella -containing vacuole. Cell. Microbiol. 2015, 17, 951–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Lin, F.; Cabello, A.L.; da Costa, L.F.; Feng, X.; Feng, H.-Q.; Zhang, M.-Z.; Iwawaki, T.; Rice-Ficht, A.; Ficht, T.A.; et al. Activation of Host IRE1α-Dependent Signaling Axis Contributes the Intracellular Parasitism of Brucella melitensis. Front. Cell. Infect. Microbiol. 2018, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.; Child, R.; Wehrly Tara, D.; Hansen, B.; Hwang, S.; López-Otin, C.; Virgin Herbert, W.; Celli, J. Selective Subversion of Autophagy Complexes Facilitates Completion of the Brucella Intracellular Cycle. Cell Host Microbe 2012, 11, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Roy, C.R.; Salcedo, S.P.; Gorvel, J.-P.E. Pathogen–endoplasmic-reticulum interactions: In through the out door. Nat. Rev. Immunol. 2006, 6, 136–147. [Google Scholar] [CrossRef]
- Celli, J. Surviving inside a macrophage: The many ways of Brucella. Res. Microbiol. 2006, 157, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, S.P.; Marchesini, M.I.; Lelouard, H.; Fugier, E.; Jolly, G.; Balor, S.; Muller, A.; Lapaque, N.; Demaria, O.; Alexopoulou, L.; et al. Brucella Control of Dendritic Cell Maturation Is Dependent on the TIR-Containing Protein Btp1. PLoS Pathog. 2008, 4, e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demars, A.; Lison, A.; Machelart, A.; Van Vyve, M.; Potemberg, G.; Vanderwinden, J.-M.; De Bolle, X.; Letesson, J.-J.; Muraille, E. Route of Infection Strongly Impacts the Host-Pathogen Relationship. Front. Immunol. 2019, 10, 1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro-Cerdá, J.; Méresse, S.; Parton, R.G.; van der Goot, G.; Sola-Landa, A.; Lopez-Goñi, I.; Moreno, E.; Gorvel, J.-P. Brucella abortus Transits through the Autophagic Pathway and Replicates in the Endoplasmic Reticulum of Nonprofessional Phagocytes. Infect. Immun. 1998, 66, 5711–5724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barquero-Calvo, E.; Mora-Cartín, R.; Arce-Gorvel, V.; de Diego, J.L.; Chacón-Díaz, C.; Chaves-Olarte, E.; Guzmán-Verri, C.; Buret, A.G.; Gorvel, J.-P.; Moreno, E. Brucella abortus Induces the Premature Death of Human Neutrophils through the Action of Its Lipopolysaccharide. PLoS Pathog. 2015, 11, e1004853. [Google Scholar] [CrossRef] [Green Version]
- Goenka, R.; Guirnalda, P.D.; Black, S.J.; Baldwin, C.L. B Lymphocytes Provide an Infection Niche for Intracellular Bacterium Brucella abortus. J. Infect. Dis. 2012, 206, 91–98. [Google Scholar] [CrossRef]
- Vitry, M.-A.; Hanot Mambres, D.; Deghelt, M.; Hack, K.; Machelart, A.; Lhomme, F.; Vanderwinden, J.-M.; Vermeersch, M.; De Trez, C.; Pérez-Morga, D.; et al. Brucella melitensis invades murine erythrocytes during infection. Infect. Immun. 2014, 82, 3927–3938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo, S.P.; Chevrier, N.; Lacerda, T.L.S.; Ben Amara, A.; Gerart, S.; Gorvel, V.A.; de Chastellier, C.; Blasco, J.M.; Mege, J.-L.; Gorvel, J.-P. Pathogenic Brucellae Replicate in Human Trophoblasts. J. Infect. Dis. 2013, 207, 1075–1083. [Google Scholar] [CrossRef]
- Yona, S.; Gordon, S. From the Reticuloendothelial to Mononuclear Phagocyte System—The Unaccounted Years. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Summers, K.M.; Bush, S.J.; Hume, D.A. Network analysis of transcriptomic diversity amongst resident tissue macrophages and dendritic cells in the mouse mononuclear phagocyte system. PLoS Biol. 2020, 18, e3000859. [Google Scholar] [CrossRef]
- Cervantes, F.; Carbonell, J.; Bruguera, M.; Force, L.; Webb, S. Liver disease in brucellosis. A clinical and pathological study of 40 cases. Postgrad. Med. J. 1982, 58, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Detilleux, P.G.; Cheville, N.F.; Deyoe, B.L. Pathogenesis of Brucella abortus in Chicken Embryos. Vet. Pathol. 1988, 25, 138–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daggett, J.; Rogers, A.; Harms, J.; Splitter, G.A.; Durward-Diioia, M. Hepatic and splenic immune response during acute vs. chronic Brucella melitensis infection using in situ microscopy. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101490. [Google Scholar] [CrossRef]
- Karim, M.F.; Maruf, A.A.; Yeasmin, F.; Shafy, N.M.; Khan, A.H.N.A.; Rahman, A.K.M.A.; Bhuiyan, M.J.S.; Hasan, M.M.; Karim, M.R.; Hasan, M.T.; et al. Histopathological changes of brucellosis in experimentally infected guinea pig. Bangladesh J. Vet. Med. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- Copin, R.; Vitry, M.-A.; Hanot Mambres, D.; Machelart, A.; De Trez, C.; Vanderwinden, J.-M.; Magez, S.; Akira, S.; Ryffel, B.; Carlier, Y.; et al. In Situ Microscopy Analysis Reveals Local Innate Immune Response Developed around Brucella Infected Cells in Resistant and Susceptible Mice. PLoS Pathog. 2012, 8, e1002575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Jiménez, C.; Mora-Cartín, R.; Altamirano-Silva, P.; Chacón-Díaz, C.; Chaves-Olarte, E.; Moreno, E.; Barquero-Calvo, E. Neutrophils as Trojan Horse Vehicles for Brucella abortus Macrophage Infection. Front. Immunol. 2019, 10, 1012. [Google Scholar] [CrossRef] [Green Version]
- Peyron, P.; Vaubourgeix, J.; Poquet, Y.; Levillain, F.; Botanch, C.; Bardou, F.; Daffé, M.; Emile, J.-F.; Marchou, B.; Cardona, P.-J.; et al. Foamy Macrophages from Tuberculous Patients’ Granulomas Constitute a Nutrient-Rich Reservoir for M. tuberculosis Persistence. Plos Pathog. 2008, 4, e1000204. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.; Schnurrenberger, P.R.; Brown, R.R. Numbers of Brucella abortus in the placenta, umbilicus and fetal fluid of two naturally infected cows. Vet. Rec. 1981, 108, 500. [Google Scholar] [CrossRef]
- Smith, H.; Williams, A.E.; Pearce, J.H.; Keppie, J.; Harris-Smith, P.W.; Fitz-George, R.B.; Witt, K. Fœtal Erythritol: A Cause of the Localization of Brucella abortus in Bovine Contagious Abortion. Nature 1962, 193, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.B.K.; Graham, E.F.; Lewis, B.A.; Smith, F. D-mannitol, erythritol and glycerol in bovine semen. Reproduction 1967, 13, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Essenberg, R.C.; Seshadri, R.; Nelson, K.; Paulsen, I. Sugar metabolism by Brucellae. Vet. Microbiol. 2002, 90, 249–261. [Google Scholar] [CrossRef]
- Keppie, J.; Williams, A.E.; Witt, K.; Smith, H. The role of erythritol in the tissue localization of the brucellae. Br. J. Exp. Pathol. 1965, 46, 104–108. [Google Scholar]
- Letesson, J.-J.; Barbier, T.; Zúñiga-Ripa, A.; Godfroid, J.; De Bolle, X.; Moriyón, I. Brucella Genital Tropism: What’s on the Menu. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Samartino, L.E.; Enright, F.M. Pathogenesis of abortion of bovine brucellosis. Comp. Immunol. Microbiol. Infect. Dis. 1993, 16, 95–101. [Google Scholar] [CrossRef]
- Keppie, J.; Witt, K.; Smith, H. The Effect of Erythritol on the Growth of S19 and other Attenuated Strains of Brucella abortus. Res. Vet. Sci. 1967, 8, 294–296. [Google Scholar] [CrossRef]
- Rodríguez, M.C.; Viadas, C.; Seoane, A.; Sangari, F.J.; López-Goñi, I.; García-Lobo, J.M. Evaluation of the Effects of Erythritol on Gene Expression in Brucella abortus. PLoS ONE 2012, 7, e50876. [Google Scholar] [CrossRef] [Green Version]
- Corner, L.A.; Alton, G.G. Persistence of Brucella abortus strain 19 infection in adult cattle vaccinated with reduced doses. Res. Vet. Sci. 1981, 31, 342–344. [Google Scholar] [CrossRef]
- Alexander, D.P.; Huggett, A.S.G.; Nixon, D.A.; Widdas, W.F. The placental transfer of sugars in the sheep: The influence of concentration gradient upon the rates of hexose formation as shown in umbilical perfusion of the placenta. J. Physiol. 1955, 129, 367–383. [Google Scholar] [CrossRef] [Green Version]
- Hastein, T.; Velle, W. Placental aldose reductase activity and foetal blood fructose during bovine pregnancy. Reproduction 1968, 15, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Gagnaire, A.; Gorvel, L.; Papadopoulos, A.; Von Bargen, K.; Mège, J.-L.; Gorvel, J.-P. COX-2 Inhibition Reduces Brucella Bacterial Burden in Draining Lymph Nodes. Front. Microbiol. 2016, 7, 1987. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.D.; Cheville, N.F.; Meador, V.P. Pathogenesis of Placentitis in the Goat Inoculated with Brucella abortus. II. Ultrastructural Studies. Vet. Pathol. 1986, 23, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Queipo-Ortuño, M.I.; Colmenero, J.D.; Muñoz, N.; Baeza, G.; Clavijo, E.; Morata, P. Rapid Diagnosis of Brucella Epididymo-Orchitis by Real-Time Polymerase Chain Reaction Assay in Urine Samples. J. Urol. 2006, 176, 2290–2293. [Google Scholar] [CrossRef] [PubMed]
- Al-Tawfiq, J.; Memish, Z. Pregnancy Associated Brucellosis. Recent Pat. Anti-Infect. Drug Discov. 2013, 8, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Jiménez, C.; Hysenaj, L.; Alfaro-Alarcón, A.; Mora-Cartín, R.; Arce-Gorvel, V.; Moreno, E.; Gorvel, J.P.; Barquero-Calvo, E. Persistence of Brucella abortus in the Bone Marrow of Infected Mice. J. Immunol. Res. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Magnani, D.M.; Lyons, E.T.; Forde, T.S.; Shekhani, M.T.; Adarichev, V.A.; Splitter, G.A. Osteoarticular tissue infection and development of skeletal pathology in murine brucellosis. Dis. Models Mech. 2013, 6, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacón-Díaz, C.; Altamirano-Silva, P.; González-Espinoza, G.; Medina, M.-C.; Alfaro-Alarcón, A.; Bouza-Mora, L.; Jiménez-Rojas, C.; Wong, M.; Barquero-Calvo, E.; Rojas, N.; et al. Brucella canis Is an Intracellular Pathogen That Induces a Lower Proinflammatory Response than Smooth Zoonotic Counterparts. Infect. Immun. 2015, 83, 4861–4870. [Google Scholar] [CrossRef] [Green Version]
- Burberry, A.; Zeng, M.Y.; Ding, L.; Wicks, I.; Inohara, N.; Morrison, S.J.; Núñez, G. Infection Mobilizes Hematopoietic Stem Cells through Cooperative NOD-like Receptor and Toll-like Receptor Signaling. Cell Host Microbe 2014, 15, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, H.; Fritsch, K.; Kovtonyuk, L.V.; Saito, Y.; Yakkala, C.; Jacobs, K.; Ahuja, A.K.; Lopes, M.; Hausmann, A.; Hardt, W.-D.; et al. Pathogen-Induced TLR4-TRIF Innate Immune Signaling in Hematopoietic Stem Cells Promotes Proliferation but Reduces Competitive Fitness. Cell Stem Cell 2017, 21, 225–240.e225. [Google Scholar] [CrossRef]
- Hysenaj, L.; De Laval, B.; Arce-Gorvel, V.; Bosilkovski, M.; Gonzalez-Espinoza, G.; Debroas, G.; Sieweke, M.; Sarrazin, S.; Gorvel, J.-P. Brucella subverts myeloid commitment of hematopoietic stem cells by a SLAMF1 (CD150)-dependent mechanism. Cell Stem Cell 2021, 26, 657–674. [Google Scholar]
- Demir, C.; Karahocagil, M.K.; Esen, R.; Atmaca, M.; Gönüllü, H.; Akdeniz, H. Bone marrow biopsy findings in brucellosis patients with hematologic abnormalities. Chin. Med. J. 2012, 125, 1871–1876. [Google Scholar]
- Ganado, W.; Bannister, W. Bacteraemia in Human Brucellosis. BMJ 1960, 1, 601–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotuzzo, E.; Carrillo, C.; Guerra, J.; Llosa, L. An Evaluation of Diagnostic Methods for Brucellosis—The Value of Bone Marrow Culture. J. Infect. Dis. 1986, 153, 122–125. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Yrivarren, J.L.; Argumans, C.; Crosby, E.; Carrillo, C.; Gotuzzo, E. Evaluation of the bone marrow in patients with brucellosis. Clinico-pathological correlation. Enferm. Infecc. Y Microbiol. Clin. 1990, 8, 19–24. [Google Scholar]
- Crosby, E.; Llosa, L.; Quesada, M.M.; Carrillo, P.C.; Gotuzzo, E. Hematologic Changes in Brucellosis. J. Infect. Dis. 1984, 150, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Ertem, M.; Kürekçi, A.; Aysev, D.; Ünal, E.; Ikincioğulları, A. Brucellosis transmitted by bone marrow transplantation. Bone Marrow Transplant. 2000, 26, 225–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naparstek, E. Transmission of brucellosis by bone marrow transplantation. Lancet 1982, 319, 574–575. [Google Scholar] [CrossRef]
- Bracewell, C.; Corbel, M. An association between arthritis and persistent serological reactions to Brucella abortus in cattle from apparently brucellosis-free herds. Vet. Rec. 1980, 106, 99–101. [Google Scholar] [CrossRef] [PubMed]
- Corbel, M.J.; Stuart, F.A.; Brewer, R.A.; Jeffrey, M.; Bradley, R. Arthropathy associated with Brucella abortus strain 19 vaccination in cattle. I. Examination of field cases. Br. Vet. J. 1989, 145, 337–346. [Google Scholar] [CrossRef]
- Delpino, M.V.; Fossati, C.A.; Baldi, P.C. Proinflammatory Response of Human Osteoblastic Cell Lines and Osteoblast-Monocyte Interaction upon Infection with Brucella spp. Infect. Immun. 2009, 77, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Khalaf, O.H.; Chaki, S.P.; Garcia-Gonzalez, D.G.; Suva, L.J.; Gaddy, D.; Arenas-Gamboa, A.M. Interaction of Brucella abortus with Osteoclasts: A Step toward Understanding Osteoarticular Brucellosis and Vaccine Safety. Infect. Immun. 2020, 88, e00822-19. [Google Scholar] [CrossRef]
- Scian, R.; Barrionuevo, P.; Giambartolomei, G.H.; De Simone, E.A.; Vanzulli, S.I.; Fossati, C.A.; Baldi, P.C.; Delpino, M.V. Potential Role of Fibroblast-Like Synoviocytes in Joint Damage Induced by Brucella abortus Infection through Production and Induction of Matrix Metalloproteinases. Infect. Immun. 2011, 79, 3619–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giambartolomei, G.H.; Arriola Benitez, P.C.; Delpino, M.V. Brucella and Osteoarticular Cell Activation: Partners in Crime. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Supakorndej, T.; Krambs, J.R.; Rao, M.; Abou-Ezzi, G.; Ye, R.Y.; Li, S.; Trinkaus, K.; Link, D.C. Bone marrow dendritic cells regulate hematopoietic stem/progenitor cell trafficking. J. Clin. Investig. 2019, 129, 2920–2931. [Google Scholar] [CrossRef]
- Winkler, I.G.; Sims, N.A.; Pettit, A.R.; Barbier, V.; Nowlan, B.; Helwani, F.; Poulton, I.J.; van Rooijen, N.; Alexander, K.A.; Raggatt, L.J.; et al. Bone marrow macrophages maintain hematopoietic stem cell (HSC) niches and their depletion mobilizes HSCs. Blood 2010, 116, 4815–4828. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Acebes, M.; Pitaval, C.; Weiss, L.A.; Nombela-Arrieta, C.; Chèvre, R.; A-González, N.; Kunisaki, Y.; Zhang, D.; van Rooijen, N.; Silberstein, L.E.; et al. Rhythmic Modulation of the Hematopoietic Niche through Neutrophil Clearance. Cell 2013, 153, 1025–1035. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.C.; Saraiva, M.; Gomes, M.S. The bone marrow hematopoietic niche and its adaptation to infection. Semin. Cell Dev. Biol. 2020, S1084952119302617. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Kashino, S.S.; Pulu, I.; Kalita, D.; Swami, V.; Yeger, H.; Felsher, D.W.; Campos-Neto, A. CD271+ Bone Marrow Mesenchymal Stem Cells May Provide a Niche for Dormant Mycobacterium tuberculosis. Sci. Transl. Med. 2013, 5, 170ra113. [Google Scholar] [CrossRef] [Green Version]
- Tornack, J.; Reece, S.T.; Bauer, W.M.; Vogelzang, A.; Bandermann, S.; Zedler, U.; Stingl, G.; Kaufmann, S.H.E.; Melchers, F. Human and Mouse Hematopoietic Stem Cells Are a Depot for Dormant Mycobacterium tuberculosis. PLoS ONE 2017, 12, e0169119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, J.; Chu, P.; Contag, C.H. Foci of Listeria monocytogenes persist in the bone marrow. Dis. Models Mech. 2009, 2, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, E.E. Brucellosis as a Hazard of Blood Transfusion. BMJ 1955, 1, 27–28. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, G.; Ataide, M.; Kastenmüller, W. Lymph node—An organ for T-cell activation and pathogen defense. Immunol. Rev. 2016, 271, 200–220. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurty, A.T.; Turley, S.J. Lymph node stromal cells: Cartographers of the immune system. Nat. Immunol. 2020, 21, 369–380. [Google Scholar] [CrossRef]
- Jackson, D.G. Leucocyte Trafficking via the Lymphatic Vasculature— Mechanisms and Consequences. Front. Immunol. 2019, 10, 471. [Google Scholar] [CrossRef]
- Lynskey, N.N.; Banerji, S.; Johnson, L.A.; Holder, K.A.; Reglinski, M.; Wing, P.A.C.; Rigby, D.; Jackson, D.G.; Sriskandan, S. Rapid Lymphatic Dissemination of Encapsulated Group A Streptococci via Lymphatic Vessel Endothelial Receptor-1 Interaction. PLoS Pathog. 2015, 11, e1005137. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, W.P. The Nature of Brucellosis. Am. J. Trop. Med. Hyg. 1957, 6, 591–592. [Google Scholar] [CrossRef]
- Fensterbank, R. Some aspects of experimental bovine brucellosis. Ann. Rech. Vet. Vet. Ann. Vet. Res. 1987, 18, 421–428. [Google Scholar]
- Meador, V.P.; Deyoe, B.L.; Cheville, N.F. Effect of Nursing on Brucella abortus Infection of Mammary Glands of Goats. Vet. Pathol. 1989, 26, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Meador, V.P.; Deyoe, B.L.; Cheville, N.F. Pathogenesis of Brucella abortus Infection of the Mammary Gland and Supramammary Lymph Node of the Goat. Vet. Pathol. 1989, 26, 357–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scientific Committe on Animal Health and Animal Welfare. Brucellosis in Sheep and Goats (Brucella melitensis). 2001. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/sci-com_scah_out59_en.pdf (accessed on 15 December 2020).
- Damir, H.A.; Tageldin, M.H.; Kenyon, S.J.; Idris, O.F. Isolation of Brucella abortus from experimentally infected dromedary camels in Sudan: A preliminary report. Vet. Res. Commun. 1989, 13, 403–406. [Google Scholar] [CrossRef]
- Foster, J.T.; Walker, F.M.; Rannals, B.D.; Hussain, M.H.; Drees, K.P.; Tiller, R.V.; Hoffmaster, A.R.; Al-Rawahi, A.; Keim, P.; Saqib, M. African Lineage Brucella melitensis Isolates from Omani Livestock. Front. Microbiol. 2018, 8, 2702. [Google Scholar] [CrossRef]
- Marin, C.M.; Jimenez de Bagues, M.P.; Barberan, M.; Blasco, J.M. Comparison of two selective media for the isolation of Brucella melitensis from naturally infected sheep and goats. Vet. Rec. 1996, 138, 409–411. [Google Scholar] [CrossRef]
- Cruz-Migoni, S.; Caamaño, J. Fat-Associated Lymphoid Clusters in Inflammation and Immunity. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef]
- Ferrante, A.W. The immune cells in adipose tissue. Diabetes Obes. Metab. 2013, 15, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Bechah, Y.; Paddock, C.D.; Capo, C.; Mege, J.-L.; Raoult, D. Adipose Tissue Serves as a Reservoir for Recrudescent Rickettsia prowazekii Infection in a Mouse Model. PLoS ONE 2010, 5, e8547. [Google Scholar] [CrossRef]
- Beigier-Bompadre, M.; Montagna, G.N.; Kühl, A.A.; Lozza, L.; Weiner, J.; Kupz, A.; Vogelzang, A.; Mollenkopf, H.-J.; Löwe, D.; Bandermann, S.; et al. Mycobacterium tuberculosis infection modulates adipose tissue biology. PLoS Pathog. 2017, 13, e1006676. [Google Scholar] [CrossRef] [Green Version]
- Cabalén, M.E.; Cabral, M.F.; Sanmarco, L.M.; Andrada, M.C.; Onofrio, L.I.; Ponce, N.E.; Aoki, M.P.; Gea, S.; Cano, R.C. Chronic Trypanosoma cruzi infection potentiates adipose tissue macrophage polarization toward an anti-inflammatory M2 phenotype and contributes to diabetes progression in a diet-induced obesity model. Oncotarget 2016, 7, 13400–13415. [Google Scholar] [CrossRef] [Green Version]
- Damouche, A.; Lazure, T.; Avettand-Fènoël, V.; Huot, N.; Dejucq-Rainsford, N.; Satie, A.-P.; Mélard, A.; David, L.; Gommet, C.; Ghosn, J.; et al. Adipose Tissue Is a Neglected Viral Reservoir and an Inflammatory Site during Chronic HIV and SIV Infection. PLoS Pathog. 2015, 11, e1005153. [Google Scholar] [CrossRef] [Green Version]
- Neyrolles, O.; Hernández-Pando, R.; Pietri-Rouxel, F.; Fornès, P.; Tailleux, L.; Payán, J.A.B.; Pivert, E.; Bordat, Y.; Aguilar, D.; Prévost, M.-C.; et al. Is Adipose Tissue a Place for Mycobacterium tuberculosis Persistence? PLoS ONE 2006, 1, e43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, L. The Parasite in the Fat: A Serendipitous Finding. Trends Parasitol. 2017, 33, 255. [Google Scholar] [CrossRef]
- De Souza, T.D.; de Carvalho, T.F.; Mol, J.P.d.S.; Lopes, J.V.M.; Silva, M.F.; da Paixão, T.A.; Santos, R.L. Tissue distribution and cell tropism of Brucella canis in naturally infected canine foetuses and neonates. Sci. Rep. 2018, 8, 7203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesce Viglietti, A.I.; Giambartolomei, G.H.; Quarleri, J.; Delpino, M.V. Brucella abortus Infection Modulates 3T3-L1 Adipocyte Inflammatory Response and Inhibits Adipogenesis. Front. Endocrinol. 2020, 11, 585923. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Gao, H.; Pappas, G.; Chen, Q.; Li, M.; Xu, J.; Lai, S.; Liao, Q.; Yang, W.; Yi, Z.; et al. Clinical features of 2041 human brucellosis cases in China. PLoS ONE 2018, 13, e0205500. [Google Scholar] [CrossRef] [Green Version]
- Hansel, W. The Essentiality of the Epididymal Fat Pad for Spermatogenesis. Endocrinology 2010, 151, 5565–5567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elewa, Y.H.A.; Ichii, O.; Otsuka, S.; Hashimoto, Y.; Kon, Y. Characterization of mouse mediastinal fat-associated lymphoid clusters. Cell Tissue Res. 2014, 357, 731–741. [Google Scholar] [CrossRef]
- Buscher, K.; Wang, H.; Zhang, X.; Striewski, P.; Wirth, B.; Saggu, G.; Lütke-Enking, S.; Mayadas, T.N.; Ley, K.; Sorokin, L.; et al. Protection from septic peritonitis by rapid neutrophil recruitment through omental high endothelial venules. Nat. Commun. 2016, 7, 10828. [Google Scholar] [CrossRef]
- Cipolletta, D.; Cohen, P.; Spiegelman, B.M.; Benoist, C.; Mathis, D. Appearance and disappearance of the mRNA signature characteristic of T reg cells in visceral adipose tissue: Age, diet, and PPARγ effects. Proc. Natl. Acad. Sci. USA 2015, 112, 482–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meza-Perez, S.; Randall, T.D. Immunological Functions of the Omentum. Trends Immunol. 2017, 38, 526–536. [Google Scholar] [CrossRef]
- Xavier, M.N.; Winter, M.G.; Spees, A.M.; Nguyen, K.; Atluri, V.L.; Silva, T.M.A.; Bäumler, A.J.; Müller, W.; Santos, R.L.; Tsolis, R.M. CD4+ T Cell-derived IL-10 Promotes Brucella abortus Persistence via Modulation of Macrophage Function. PLoS Pathog. 2013, 9, e1003454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin-Bastuji, B.; Hars, J.; Drapeau, A.; Cherfa, M.-A.; Game, Y.; Le Horgne, J.-M.; Rautureau, S.; Maucci, E.; Pasquier, J.-J.; Jay, M.; et al. Reemergence of Brucella melitensis Infection in Wildlife, France. Emerg. Infect. Dis. 2014, 20, 1570–1571. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.; Gilot-Fromont, E.; Freycon, P.; Thébault, A.; Game, Y.; Toïgo, C.; Petit, E.; Barthe, M.-N.; Reynaud, G.; Jaÿ, M.; et al. High Shedding Potential and Significant Individual Heterogeneity in Naturally-Infected Alpine ibex (Capra ibex) With Brucella melitensis. Front. Microbiol. 2018, 9, 1065. [Google Scholar] [CrossRef] [PubMed]
- Banskar, S.; Bhute, S.S.; Suryavanshi, M.V.; Punekar, S.; Shouche, Y.S. Microbiome analysis reveals the abundance of bacterial pathogens in Rousettus leschenaultii guano. Sci. Rep. 2016, 6, 36948. [Google Scholar] [CrossRef]
- Bai, Y.; Urushadze, L.; Osikowicz, L.; McKee, C.; Kuzmin, I.; Kandaurov, A.; Babuadze, G.; Natradze, I.; Imnadze, P.; Kosoy, M. Molecular Survey of Bacterial Zoonotic Agents in Bats from the Country of Georgia (Caucasus). PLoS ONE 2017, 12, e0171175. [Google Scholar] [CrossRef]
- Shirima, G.M.; Kunda, J.S. Prevalence of brucellosis in the human, livestock and wildlife interface areas of Serengeti National Park, Tanzania. Onderstepoort J. Vet. Res. 2016, 83, a1032. [Google Scholar] [CrossRef]
- Tessaro, S.V.; Forbes, L.B.; Turcotte, C. A survey of brucellosis and tuberculosis in bison in and around Wood Buffalo National Park, Canada. Can. Vet. J. La Rev. Vet. Can. 1990, 31, 174–180. [Google Scholar]
- Kreizinger, Z.; Foster, J.T.; Rónai, Z.; Sulyok, K.M.; Wehmann, E.; Jánosi, S.; Gyuranecz, M. Genetic relatedness of Brucella suis biovar 2 isolates from hares, wild boars and domestic pigs. Vet. Microbiol. 2014, 172, 492–498. [Google Scholar] [CrossRef]
- Forbes, L.B.; Tessaro, S.V. Transmission of brucellosis from reindeer to cattle. J. Am. Vet. Med. Assoc. 1993, 203, 289–294. [Google Scholar]
- Foster, G.; Jahans, K.L.; Reid, R.J.; Ross, H.M. Isolation of Brucella species from cetaceans, seals and an otter. Vet. Rec. 1996, 138, 583–586. [Google Scholar] [CrossRef]
- Scholz, H.C.; Hubalek, Z.; Sedlacek, I.; Vergnaud, G.; Tomaso, H.; Al Dahouk, S.; Melzer, F.; Kampfer, P.; Neubauer, H.; Cloeckaert, A.; et al. Brucella microti sp. nov., isolated from the common vole Microtus arvalis. Int. J. Syst. Evol. Microbiol. 2008, 58, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, H.C.; Hofer, E.; Vergnaud, G.; Fleche, P.L.; Whatmore, A.M.; Dahouk, S.A.; Pfeffer, M.; Krüger, M.; Cloeckaert, A.; Tomaso, H. Isolation of Brucella microti from Mandibular Lymph Nodes of Red Foxes, Vulpes vulpes, in Lower Austria. Vector Borne Zoonotic Dis. 2009, 9, 153–156. [Google Scholar] [CrossRef]
- Kimura, M.; Une, Y.; Suzuki, M.; Park, E.-S.; Imaoka, K.; Morikawa, S. Isolation of Brucella inopinata -Like Bacteria from White’s and Denny’s Tree Frogs. Vector Borne Zoonotic Dis. 2017, 17, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Dawson, C.E.; Perrett, L.L.; Stubberfield, E.J.; Stack, J.A.; Farrelly, S.S.; Cooley, W.A.; Davison, N.J.; Quinney, S. Isolation and characterization of Brucella from the lungworms of a harbor porpoise (Phocoena phocoena). J. Wildl. Dis. 2008, 44, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenberg, T.; Riße, K.; Schauerte, N.; Geiger, C.; Blom, J.; Scholz, H.C. Isolation of a novel ‘atypical’ Brucella strain from a bluespotted ribbontail ray (Taeniura lymma). Antonie Van Leeuwenhoek 2017, 110, 221–234. [Google Scholar] [CrossRef]
- Eisenberg, T.; Schlez, K.; Fawzy, A.; Völker, I.; Hechinger, S.; Curić, M.; Schauerte, N.; Geiger, C.; Blom, J.; Scholz, H.C. Expanding the host range: Infection of a reptilian host (Furcifer pardalis) by an atypical Brucella strain. Antonie Van Leeuwenhoek 2020, 113, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Jiménez de Bagüés, M.P.; Ouahrani-Bettache, S.; Quintana Juan, F.; Mitjana, O.; Hanna, N.; Bessoles, S.; Sanchez, F.; Scholz, H.C.; Lafont, V.; Köhler, S.; et al. The New Species Brucella microti Replicates in Macrophages and Causes Death in Murine Models of Infection. J. Infect. Dis. 2010, 202, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Mora, G.; Palacios-Alfaro, J.D.; González-Barrientos, R. Wildlife reservoirs of brucellosis: Brucella in aquatic environments. Rev. Sci. Tech. 2013, 32, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.M.; Lambourn, D.M.; Jeffries, S.J.; Hall, P.B.; Rhyan, J.C.; Ewalt, D.R.; Polzin, L.M.; Cheville, N.F. Evidence of Brucella infection in Parafilaroides lungworms in a Pacific harbor seal (Phoca vitulina richardsi). J. Vet. Diagn. Investig. 1997, 9, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Perrett, L.L.; Dawson, C.E.; Davison, N.; Quinney, S. Brucella infection of lungworms from a harbour porpoise. Vet. Rec. 2004, 154, 800. [Google Scholar] [PubMed]
- Lambourn, D.M.; Garner, M.; Ewalt, D.; Raverty, S.; Sidor, I.; Jeffries, S.J.; Rhyan, J.; Gaydos, J.K. Brucella pinnipedialis infections in Pacific harbor seals (Phoca vitulina richardsi) from Washington State, USA. J. Wildl. Dis. 2013, 49, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Nymo, I.H.; Seppola, M.; Al Dahouk, S.; Bakkemo, K.R.; Jiménez de Bagüés, M.P.; Godfroid, J.; Larsen, A.K. Experimental Challenge of Atlantic Cod (Gadus morhua) with a Brucella pinnipedialis Strain from Hooded Seal (Cystophora cristata). PLoS ONE 2016, 11, e0159272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Tras, W.F.; Tayel, A.A.; Eltholth, M.M.; Guitian, J. Brucella infection in fresh water fish: Evidence for natural infection of Nile catfish, Clarias gariepinus, with Brucella melitensis. Vet. Microbiol. 2010, 141, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Tiller, R.V.; Gee, J.E.; Lonsway, D.R.; Gribble, S.; Bell, S.C.; Jennison, A.V.; Bates, J.; Coulter, C.; Hoffmaster, A.R.; De, B.K. Identification of an unusual Brucella strain (BO2) from a lung biopsy in a 52 year-old patient with chronic destructive pneumonia. BMC Microbiol. 2010, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Soler-Lloréns, P.F.; Quance, C.R.; Lawhon, S.D.; Stuber, T.P.; Edwards, J.F.; Ficht, T.A.; Robbe-Austerman, S.; O’Callaghan, D.; Keriel, A. A Brucella spp. Isolate from a Pac-Man Frog (Ceratophrys ornata) Reveals Characteristics Departing from Classical Brucellae. Front. Cell. Infect. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Rouzic, N.; Desmier, L.; Cariou, M.-E.; Gay, E.; Foster, J.T.; Williamson, C.H.D.; Schmitt, F.; Le Henaff, M.; Le Coz, A.; Lorléac’h, A.; et al. First Case of Brucellosis Caused by an Amphibian-type Brucella. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ponsart, C.; Riou, M.; Locatelli, Y.; Jacques, I.; Fadeau, A.; Jay, M.; Simon, R.; Perrot, L.; Freddi, L.; Breton, S.; et al. Brucella melitensis Rev.1 vaccination generates a higher shedding risk of the vaccine strain in Alpine ibex (Capra ibex) compared to the domestic goat (Capra hircus). Vet. Res. 2019, 50, 100. [Google Scholar] [CrossRef] [Green Version]
- Eric, J.M.; Brandon, M.S.; Paul, C.C.; Jared, D.R.; William, H.E.; Benjamin, W.; Scott, G.S.; Terry, J.K. Assessment of a Strain 19 Brucellosis Vaccination Program in Elk. Wildl. Soc. Bull. 2017, 41, 70–79. [Google Scholar] [CrossRef]
- Davis, D.S.; Elzer, P.H. Brucella vaccines in wildlife. Vet. Microbiol. 2002, 90, 533–544. [Google Scholar] [CrossRef]
- Godfroid, J.; Nielsen, K.; Saegerman, C. Diagnosis of brucellosis in livestock and wildlife. Croat. Med. J. 2010, 51, 296–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadar, M.; Shahali, Y.; Fakhri, Y.; Godfroid, J. The global epidemiology of Brucella infections in terrestrial wildlife: A meta-analysis. Transbound. Emerg. Dis. 2020, tbed.13735. [Google Scholar] [CrossRef] [PubMed]
- Batista, T.G.D.S.; Fornazari, F.; Joaquim, S.F.; Latosinski, G.S.; Teixeira, C.R.; Langoni, H. Serologic Screening for Smooth Brucella sp. in Wild Animals in Brazil. J. Wildl. Dis. 2019, 55, 721–723. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Espinoza, G.; Arce-Gorvel, V.; Mémet, S.; Gorvel, J.-P. Brucella: Reservoirs and Niches in Animals and Humans. Pathogens 2021, 10, 186. https://doi.org/10.3390/pathogens10020186
González-Espinoza G, Arce-Gorvel V, Mémet S, Gorvel J-P. Brucella: Reservoirs and Niches in Animals and Humans. Pathogens. 2021; 10(2):186. https://doi.org/10.3390/pathogens10020186
Chicago/Turabian StyleGonzález-Espinoza, Gabriela, Vilma Arce-Gorvel, Sylvie Mémet, and Jean-Pierre Gorvel. 2021. "Brucella: Reservoirs and Niches in Animals and Humans" Pathogens 10, no. 2: 186. https://doi.org/10.3390/pathogens10020186
APA StyleGonzález-Espinoza, G., Arce-Gorvel, V., Mémet, S., & Gorvel, J. -P. (2021). Brucella: Reservoirs and Niches in Animals and Humans. Pathogens, 10(2), 186. https://doi.org/10.3390/pathogens10020186