
Differential Effects of Antiseptic Mouth Rinses on SARS-CoV-2 Infectivity In Vitro

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Viral Infection
2.4. Cytotoxicity Assay
2.5. Statistical Analysis
3. Results
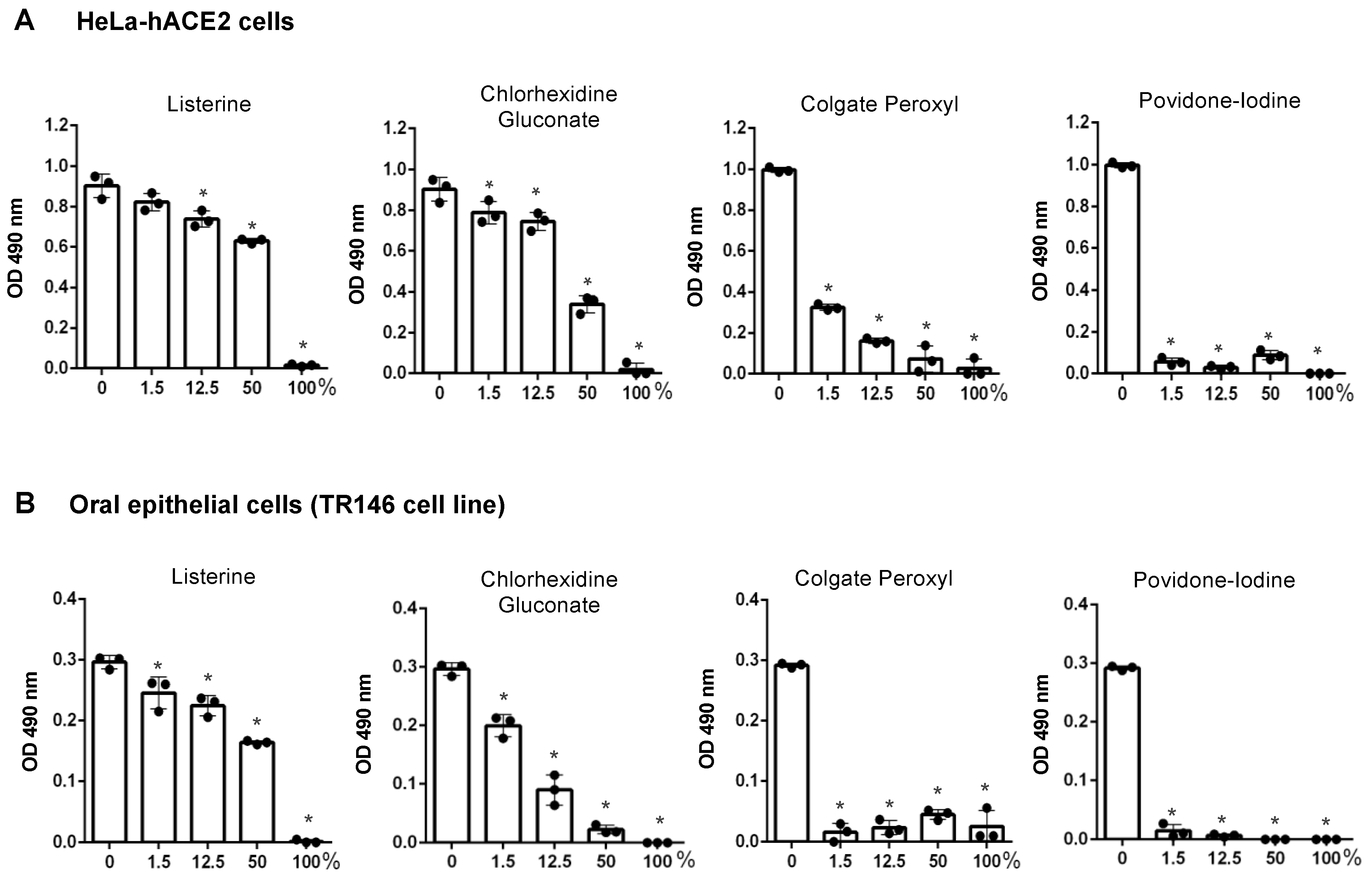
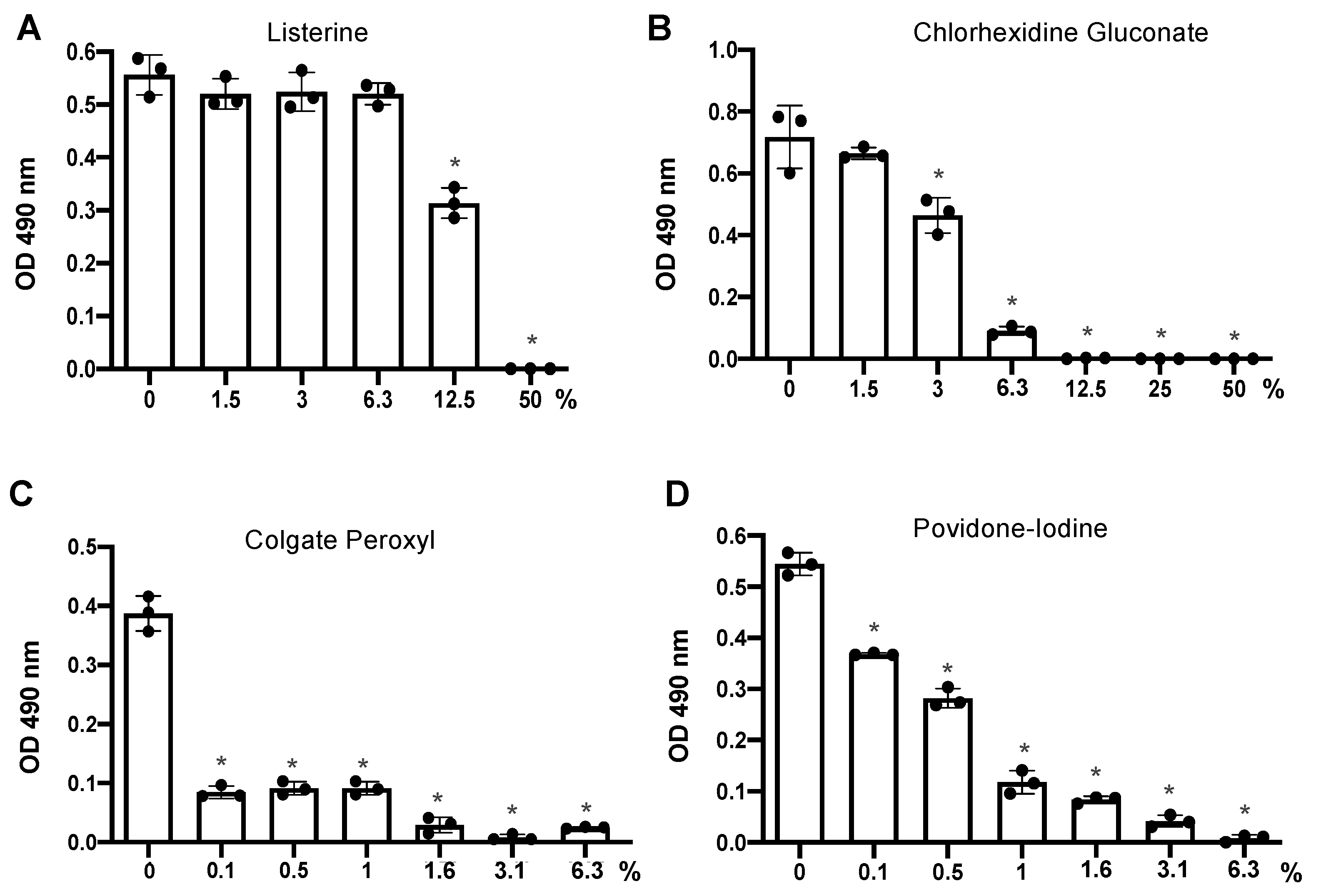
3.1. Differential Effects of Mouth Rinses on Cell Viability
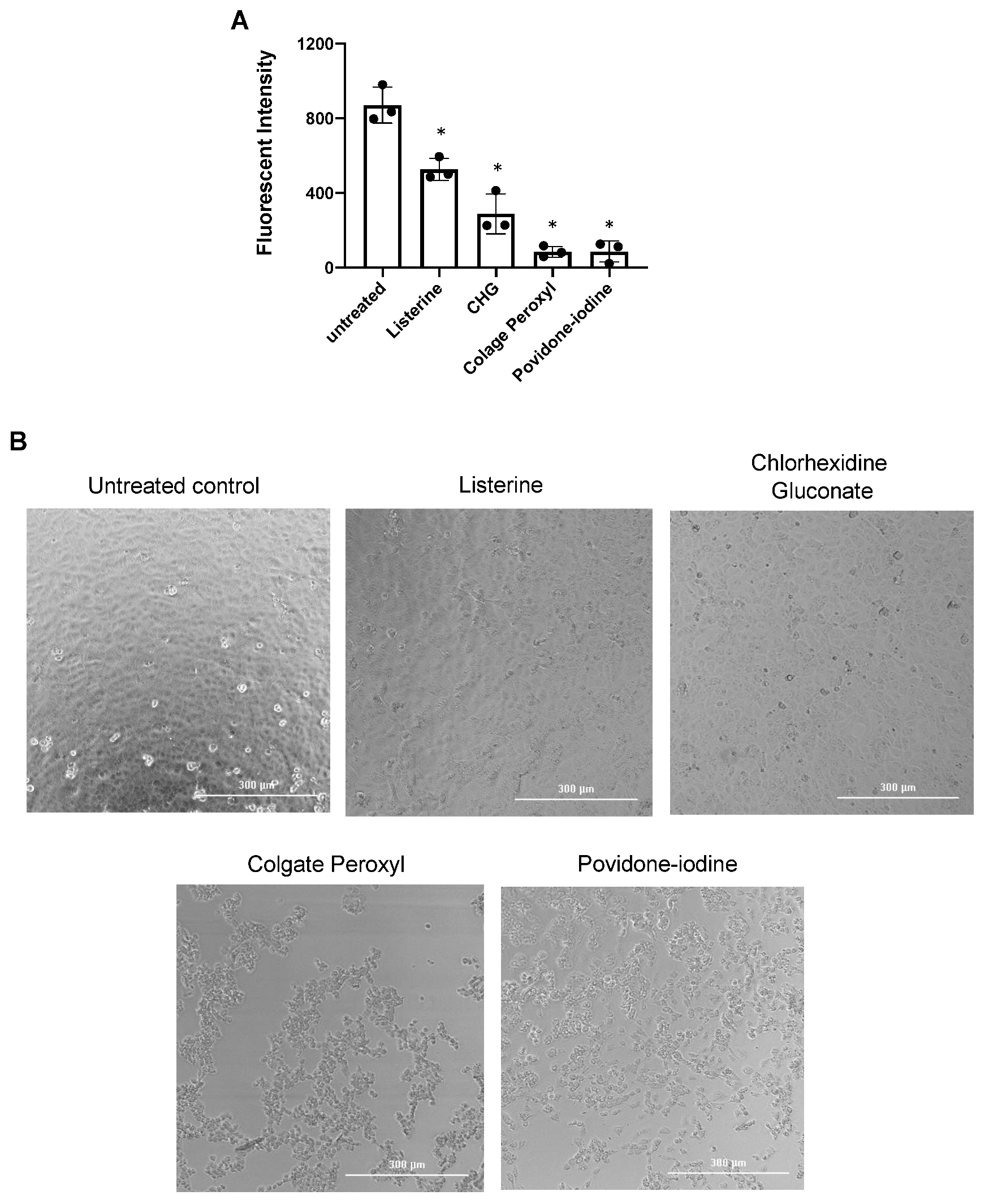
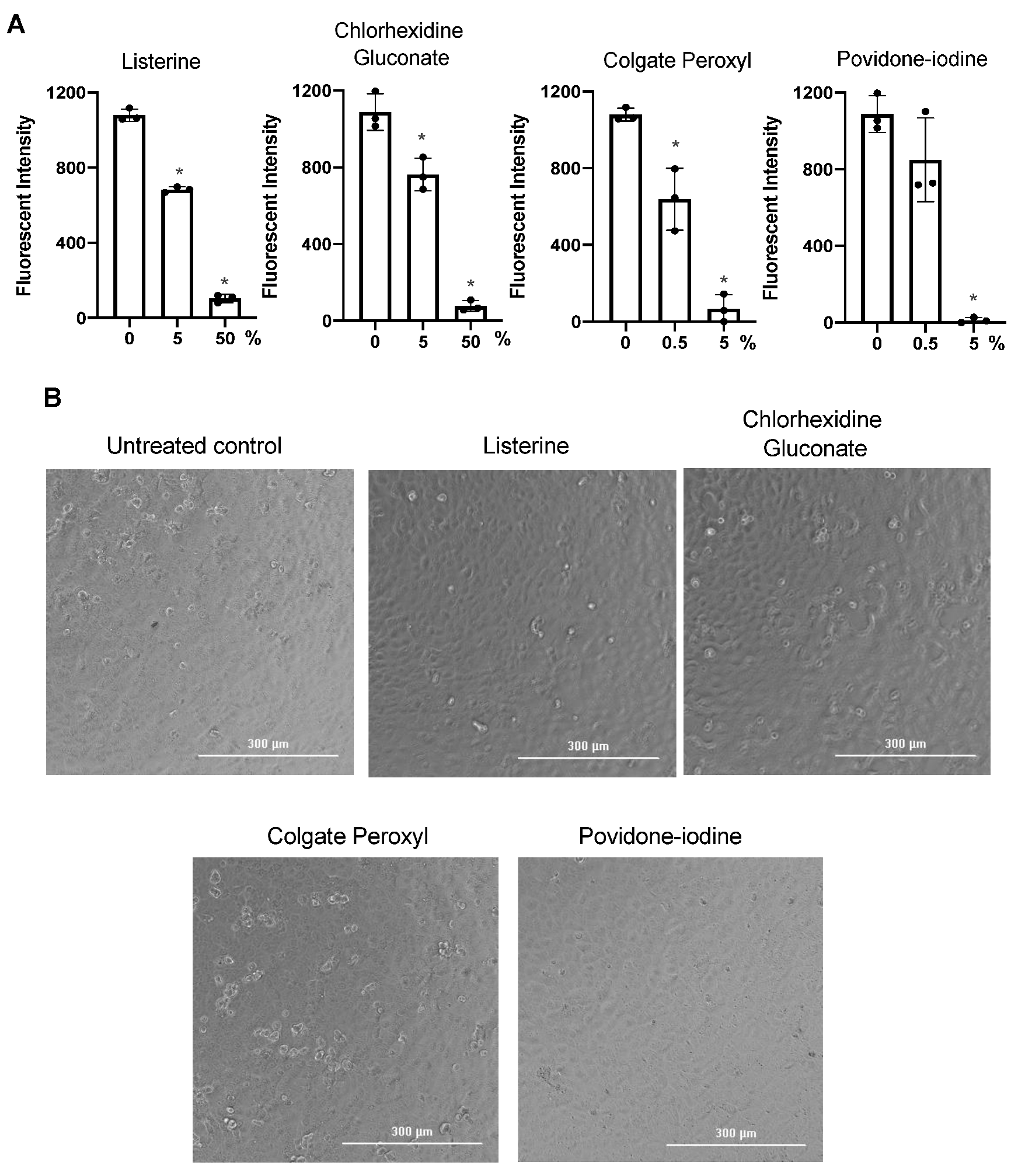
3.2. Antiviral Effect of Diluted Povidone-Iodine or Colgate Peroxl Was Associated with Cytotoxicity
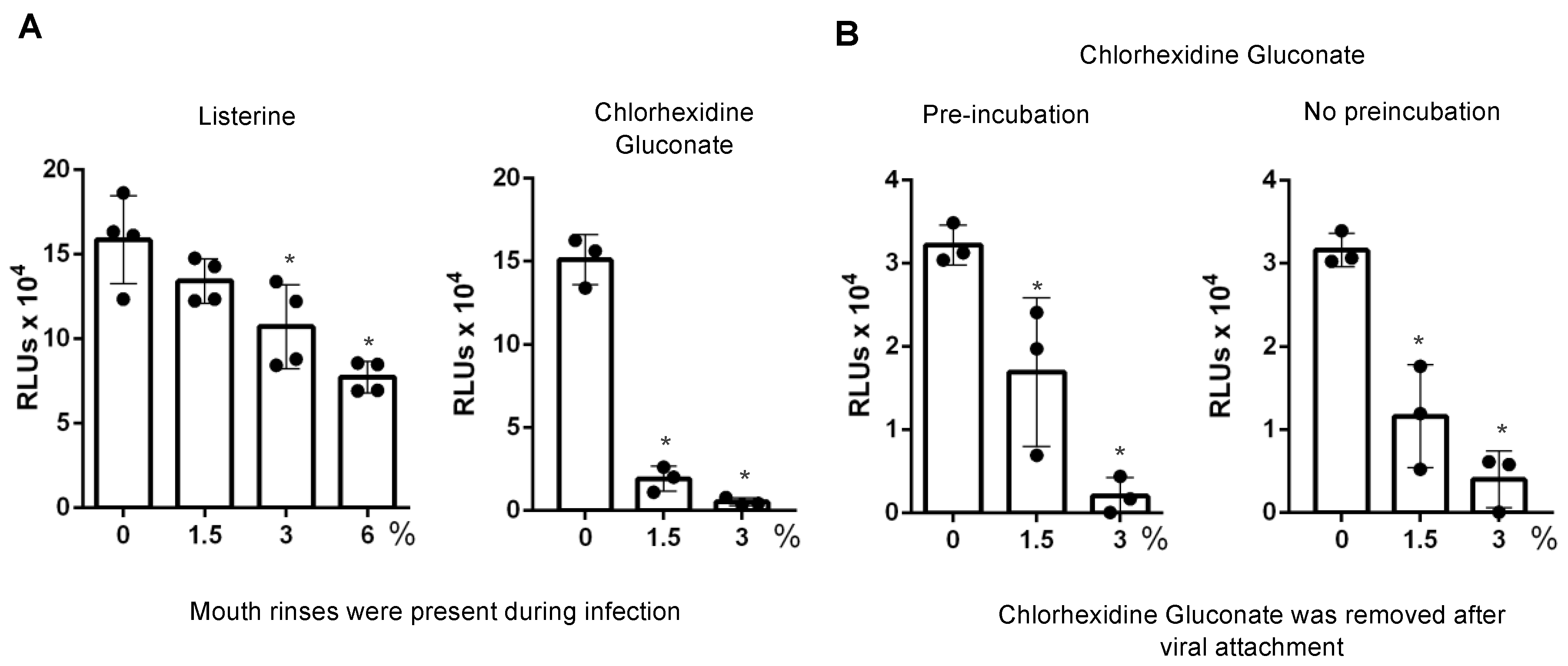
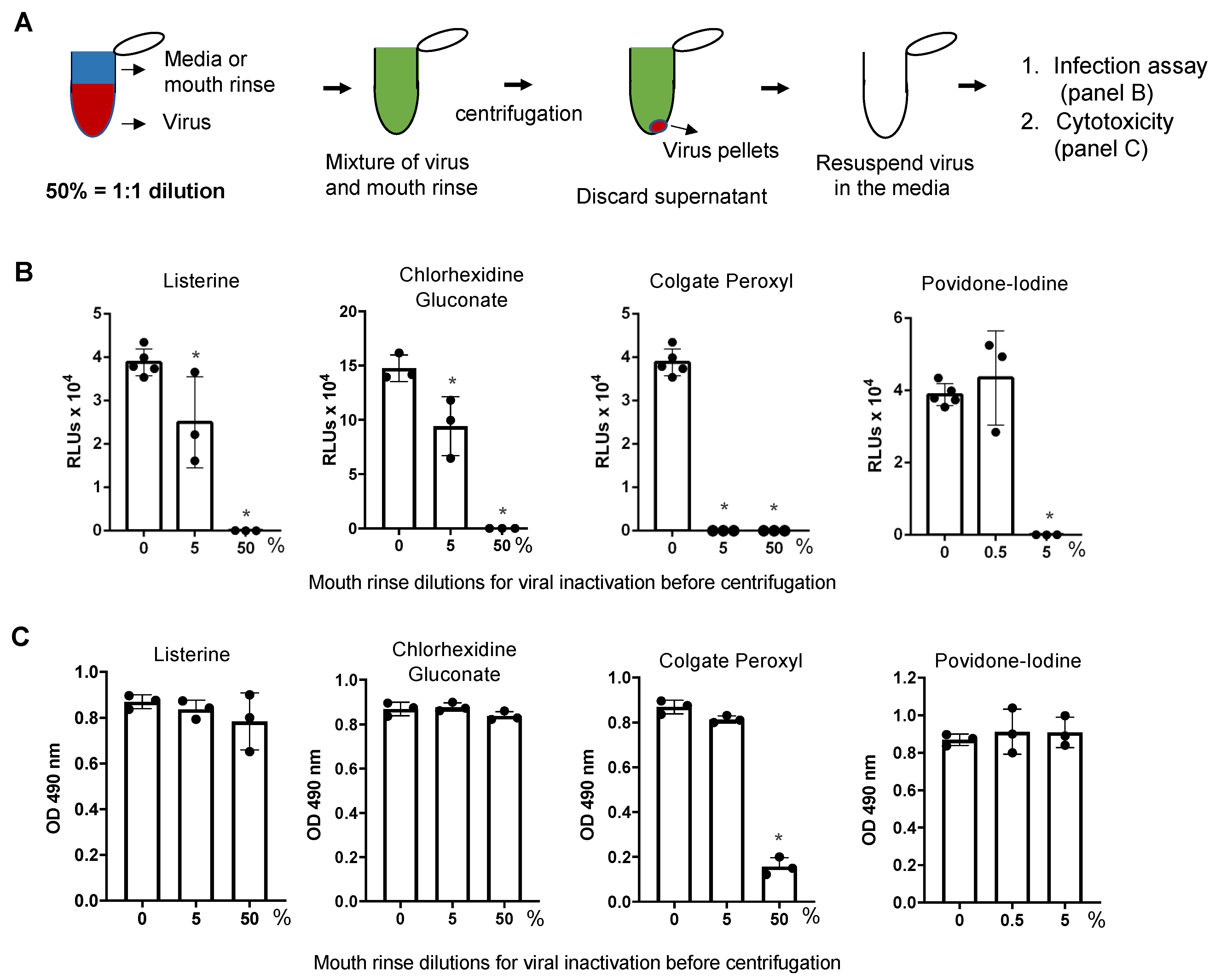
3.3. Direct Effect of Mouth Rinses on Viruses
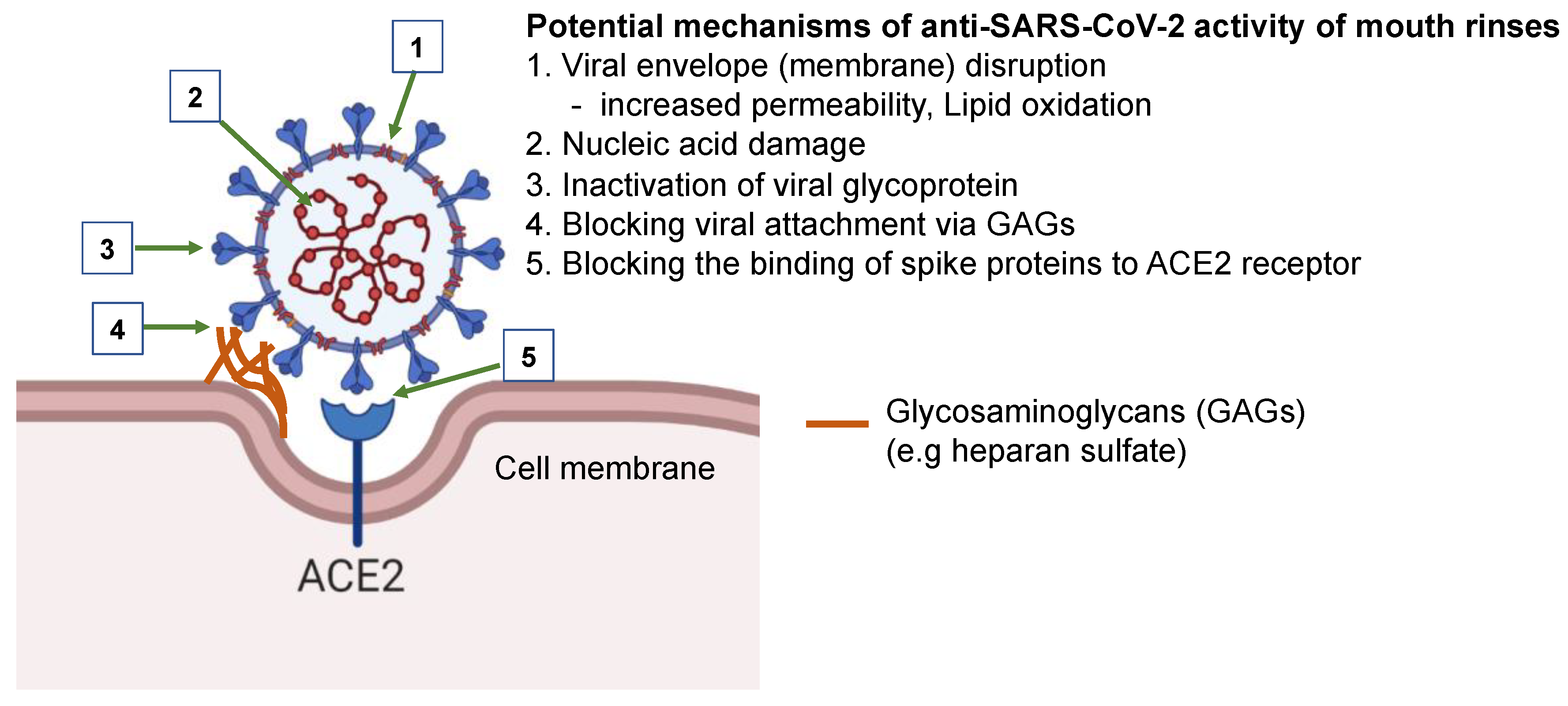
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Available online: https://covid19.who.int/ (accessed on 20 February 2021).
- CDC. Available online: https://covid.cdc.gov/covid-data-tracker/#cases_casesper100klast7days (accessed on 20 February 2021).
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Manjunath, K.; Ranjan, R.K.; Kaushik, S.; Kumar, S.; Verma, V. COVID-19 pandemic: Insights into structure, function, and hACE2 receptor recognition by SARS-CoV-2. PLoS Pathog. 2020, 16, e1008762. [Google Scholar] [CrossRef]
- Zeidler, A.; Karpinski, T.M. SARS-CoV, MERS-CoV, SARS-CoV-2 Comparison of Three Emerging Coronaviruses. Jundishapur. J. Microbiol. 2020, 13, e103744. [Google Scholar] [CrossRef]
- WHO. Transmission of SARS-CoV-2: Implication for Infection Prevention Precautions. World Health Organization. 2020. Available online: https://www.who.int/news-room/commentaries/detail/transmission-of-sars-cov-2-implications-for-infection-prevention-precautions (accessed on 9 July 2020).
- To, K.K.; Tsang, O.T.; Leung, W.S.; Tam, A.R.; Wu, T.C.; Lung, D.C.; Yip, C.C.; Cai, J.P.; Chan, J.M.; Chik, T.S.; et al. Temporal profiles of viral load in posterior oropharyngeal saliva samples and serum antibody responses during infection by SARS-CoV-2: An observational cohort study. Lancet Infect. Dis. 2020, 20, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Lin, J.; Duan, X.; Huang, W.; Lu, X.; Zhou, J.; Zong, Z. Asymptomatic COVID-19 Patients Can Contaminate Their Surroundings: An Environment Sampling Study. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, T.; Lee, E.; Lee, C.; Kim, H.; Rhee, H.; Park, S.Y.; Son, H.-J.; Yu, S.; Park, J.W.; et al. Clinical Course and Molecular Viral Shedding Among Asymptomatic and Symptomatic Patients With SARS-CoV-2 Infection in a Community Treatment Center in the Republic of Korea. JAMA Intern. Med. 2020, 180, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Fine, D.H.; Korik, I.; Furgang, D.; Myers, R.; Olshan, A.; Barnett, M.L.; Vincent, J. Assessing pre-procedural subgingival irrigation and rinsing with an antiseptic mouthrinse to reduce bacteremia. J. Am. Dent. Assoc. 1996, 127, 645–646. [Google Scholar] [CrossRef]
- Fine, D.H.; Furgang, D.; Korik, I.; Olshan, A.; Barnett, M.L.; Vincent, J.W. Reduction of viable bacteria in dental aerosols by preprocedural rinsing with an antiseptic mouthrinse. Am. J. Dent. 1993, 6, 219–221. [Google Scholar]
- Koletsi, D.; Belibasakis, G.N.; Eliades, T. Interventions to Reduce Aerosolized Microbes in Dental Practice: A Systematic Review with Network Meta-analysis of Randomized Controlled Trials. J. Dent. Res. 2020, 99, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Meiller, T.F.; Silva, A.; Ferreira, S.M.; Jabra-Rizk, M.A.; Kelley, J.I.; DePaola, L.G. Efficacy of Listerine Antiseptic in reducing viral contamination of saliva. J. Clin. Periodontol. 2005, 32, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Park, N.H. Effect of chlorhexidine on the in vitro and in vivo herpes simplex virus infection. Oral. Surg. Oral. Med. Oral. Pathol. 1989, 67, 149–153. [Google Scholar] [CrossRef]
- O’Donnell, V.B.; Thomas, D.; Stanton, R.; Maillard, J.-Y.; Murphy, R.C.; Jones, S.A.; Humphreys, I.; Wakelam, M.J.O.; Fegan, C.; Wise, M.P.; et al. Potential role of oral rinses targeting the viral lipid envelope in SARS-CoV-2 infection. Function 2020, 1, zqaa002. [Google Scholar] [CrossRef]
- Carrouel, F.; Conte, M.P.; Fisher, J.; Gonçalves, L.S.; Dussart, C.; Llodra, J.C.; Bourgeois, D. COVID-19: A Recommendation to Examine the Effect of Mouthrinses with β-Cyclodextrin Combined with Citrox in Preventing Infection and Progression. J. Clin. Med. 2020, 9, 1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrouel, F.; Gonçalves, L.S.; Conte, M.P.; Campus, G.; Fisher, J.; Fraticelli, L.; Gadea-Deschamps, E.; Ottolenghi, L.; Bourgeois, D. Antiviral Activity of Reagents in Mouth Rinses against SARS-CoV-2. J. Dent. Res. 2021, 100, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Bidra, A.S.; Pelletier, J.S.; Westover, J.B.; Frank, S.; Brown, S.M.; Tessema, B. Rapid In-Vitro Inactivation of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Using Povidone-Iodine Oral Antiseptic Rinse. J. Prosthodont. 2020, 29, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Meister, T.L.; Bruggemann, Y.; Todt, D.; Conzelmann, C.; Muller, J.A.; Gross, R.; Munch, J.; Krawczyk, A.; Steinmann, J.; Steinmann, J.; et al. Virucidal efficacy of different oral rinses against SARS-CoV-2. J. Infect. Dis 2020, 222, 1289–1292. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.E.; Sivalingam, V.; Kang, A.E.Z.; Ananthanarayanan, A.; Arumugam, H.; Jenkins, T.M.; Hadjiat, Y.; Eggers, M. Povidone-Iodine Demonstrates Rapid In Vitro Virucidal Activity Against SARS-CoV-2, The Virus Causing COVID-19 Disease. Infect. Dis. 2020, 9, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Muruato, A.; Lokugamage, K.G.; Narayanan, K.; Zhang, X.; Zou, J.; Liu, J.; Schindewolf, C.; Bopp, N.E.; Aguilar, P.V.; et al. An Infectious cDNA Clone of SARS-CoV-2. Cell Host Microbe 2020, 27, 841–848.e3. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, T.F.; Zhao, F.; Huang, D.; Beutler, N.; Burns, A.; He, W.-t.; Limbo, O.; Smith, C.; Song, G.; Woehl, J.; et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 2020, 369, 956–963. [Google Scholar] [CrossRef]
- Holmes, M.; Zhang, F.; Bieniasz, P.D. Single-Cell and Single-Cycle Analysis of HIV-1 Replication. PLoS Pathog. 2015, 11, e1004961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and influenza pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef]
- To, K.K.; Tsang, O.T.; Yip, C.C.; Chan, K.H.; Wu, T.C.; Chan, J.M.; Leung, W.S.; Chik, T.S.; Choi, C.Y.; Kandamby, D.H.; et al. Consistent Detection of 2019 Novel Coronavirus in Saliva. Clin. Infect. Dis 2020, 71, 841–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggers, M.; Koburger-Janssen, T.; Eickmann, M.; Zorn, J. In Vitro Bactericidal and Virucidal Efficacy of Povidone-Iodine Gargle/Mouthwash Against Respiratory and Oral Tract Pathogens. Infect. Dis. 2018, 7, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, J.F.; Blakely, W.F.; Joner, E.I. Mammalian cells are not killed by DNA single-strand breaks caused by hydroxyl radicals from hydrogen peroxide. Radiat Res. 1985, 103, 383–392. [Google Scholar] [CrossRef] [PubMed]
- THOMSON, D.L. The Effect of Hydrogen Peroxide on the Permeability of the Cell. J. Exp. Biol. 1928, 5, 252–257. [Google Scholar]
- Sriwilaijaroen, N.; Hiramatsu, H.; Takahashi, T.; Suzuki, T.; Ito, M.; Ito, Y.; Tashiro, M.; Suzuki, Y. Mechanisms of the action of povidone-iodine against human and avian influenza A viruses: Its effects on hemagglutination and sialidase activities. Virol. J. 2009, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- WHO Guidelines on Hand Hygiene in Health Care: First Global Patient Safety Challenge Clean Care is Safer Care. 2009. Available online: https://www.paho.org/col/index.php?option=com_docman&view=download&alias=264-who-guidelines-on-hand-hygiene&category_slug=publicaciones-ops-oms-colombia&Itemid=688 (accessed on 9 July 2020).
- Baqui, A.; Kelley, J.; Jabra-Rizk, M.A.; DePaola, L.; Falkler, W.; Meiller, T. In vitro effect of oral antiseptics on human immunodeficiency virus-1 and Herpes simplex virus type 1. J. Clin. Periodontol. 2001, 28, 610–616. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Products | Manufacturers | Active Ingredients |
|---|---|---|
| Listerine Antiseptic Original | Johnson & Johnson Consumer Inc., Skillman, NJ, USA | 20–30% ethanol Thymol 0.064% Methyl salicylate 0.06% Menthol (racementhol) 0.042% Eucalyptol 0.092% |
| Chlorhexidine gluconate | Xttrium Laboratories Inc., Mount Prospect, IL, USA | 0.12% chlorhexidine |
| Colgate Peroxyl | Colgate-Palmolive Inc., New York, NY, USA | 1.5% w/v hydrogen peroxide |
| Povidone-iodine | CVS Pharmacy Inc., Woonsocket, RI, USA | 10% solution (1% available iodine) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Wang, A.; Hoskin, E.R.; Cugini, C.; Markowitz, K.; Chang, T.L.; Fine, D.H. Differential Effects of Antiseptic Mouth Rinses on SARS-CoV-2 Infectivity In Vitro. Pathogens 2021, 10, 272. https://doi.org/10.3390/pathogens10030272
Xu C, Wang A, Hoskin ER, Cugini C, Markowitz K, Chang TL, Fine DH. Differential Effects of Antiseptic Mouth Rinses on SARS-CoV-2 Infectivity In Vitro. Pathogens. 2021; 10(3):272. https://doi.org/10.3390/pathogens10030272
Chicago/Turabian StyleXu, Chuan, Annie Wang, Eileen R. Hoskin, Carla Cugini, Kenneth Markowitz, Theresa L. Chang, and Daniel H. Fine. 2021. "Differential Effects of Antiseptic Mouth Rinses on SARS-CoV-2 Infectivity In Vitro" Pathogens 10, no. 3: 272. https://doi.org/10.3390/pathogens10030272
APA StyleXu, C., Wang, A., Hoskin, E. R., Cugini, C., Markowitz, K., Chang, T. L., & Fine, D. H. (2021). Differential Effects of Antiseptic Mouth Rinses on SARS-CoV-2 Infectivity In Vitro. Pathogens, 10(3), 272. https://doi.org/10.3390/pathogens10030272