
Calendula officinalis Triterpenoid Saponins Impact the Immune Recognition of Proteins in Parasitic Nematodes

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
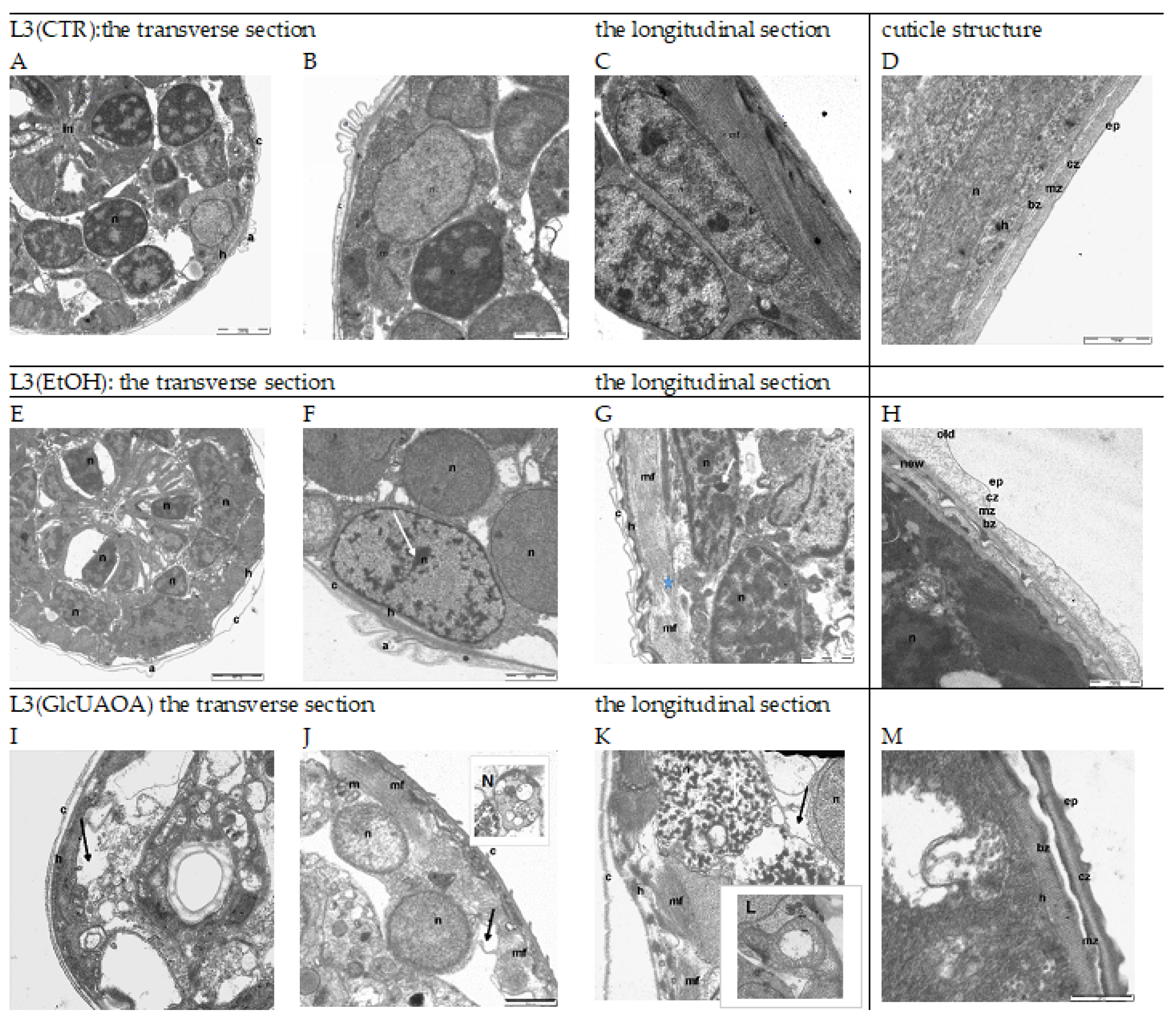
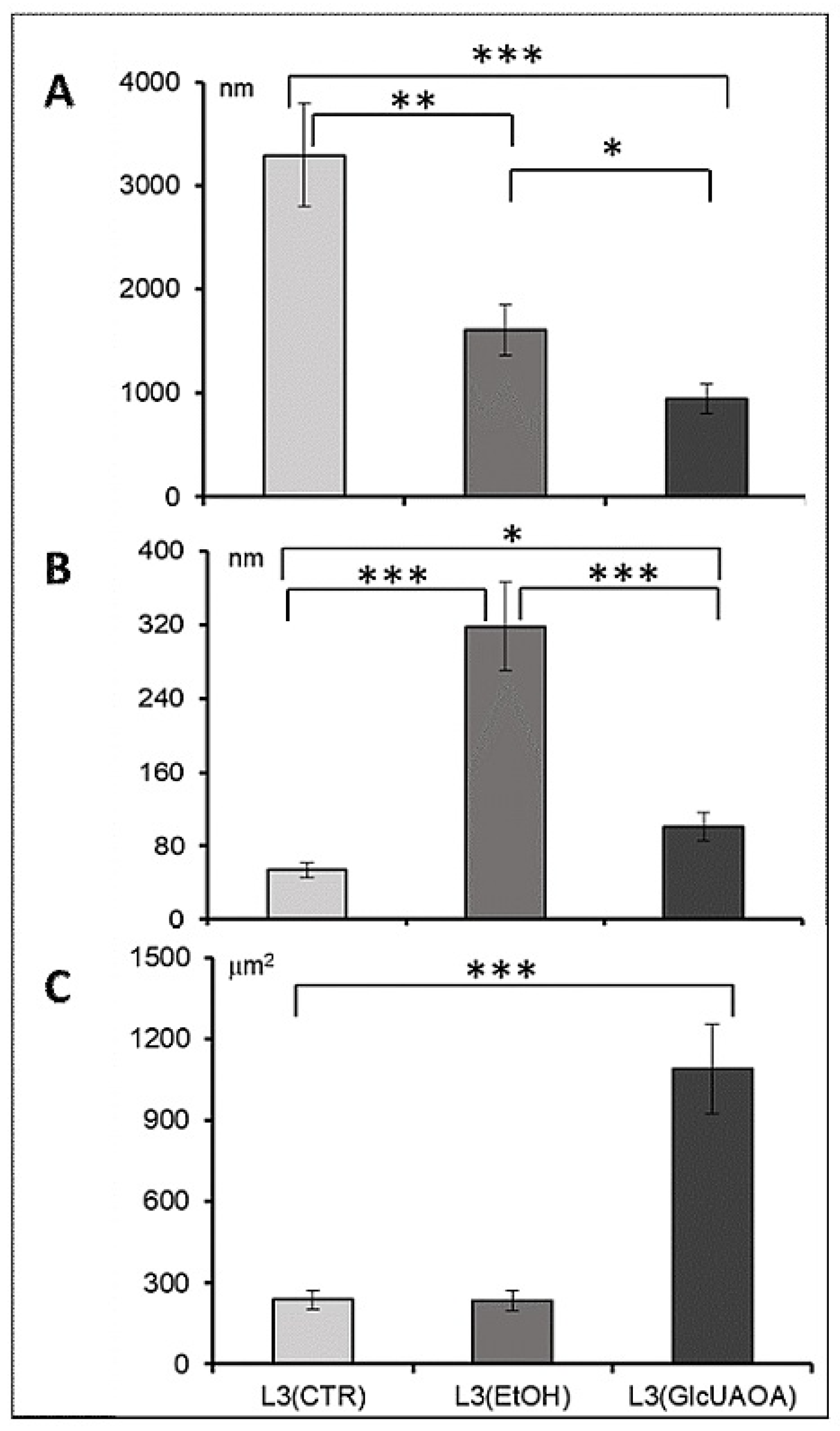
2.1. Changes in the Ultrastructure of L3 Evaluated in TEM
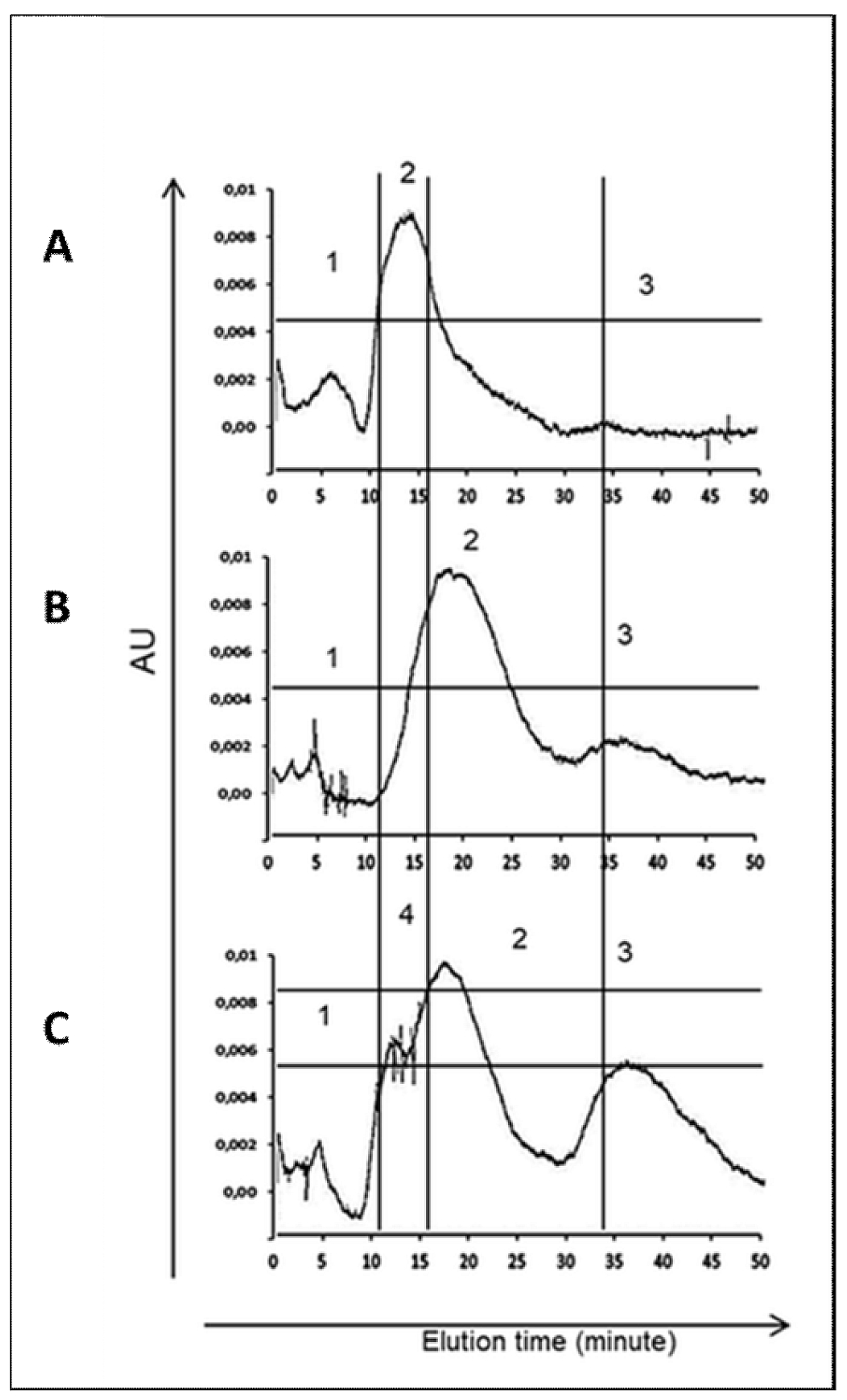
2.2. HPLC Profile of H. polygyrus bakeri L3 Larvae Exposed to C. officinals Saponins
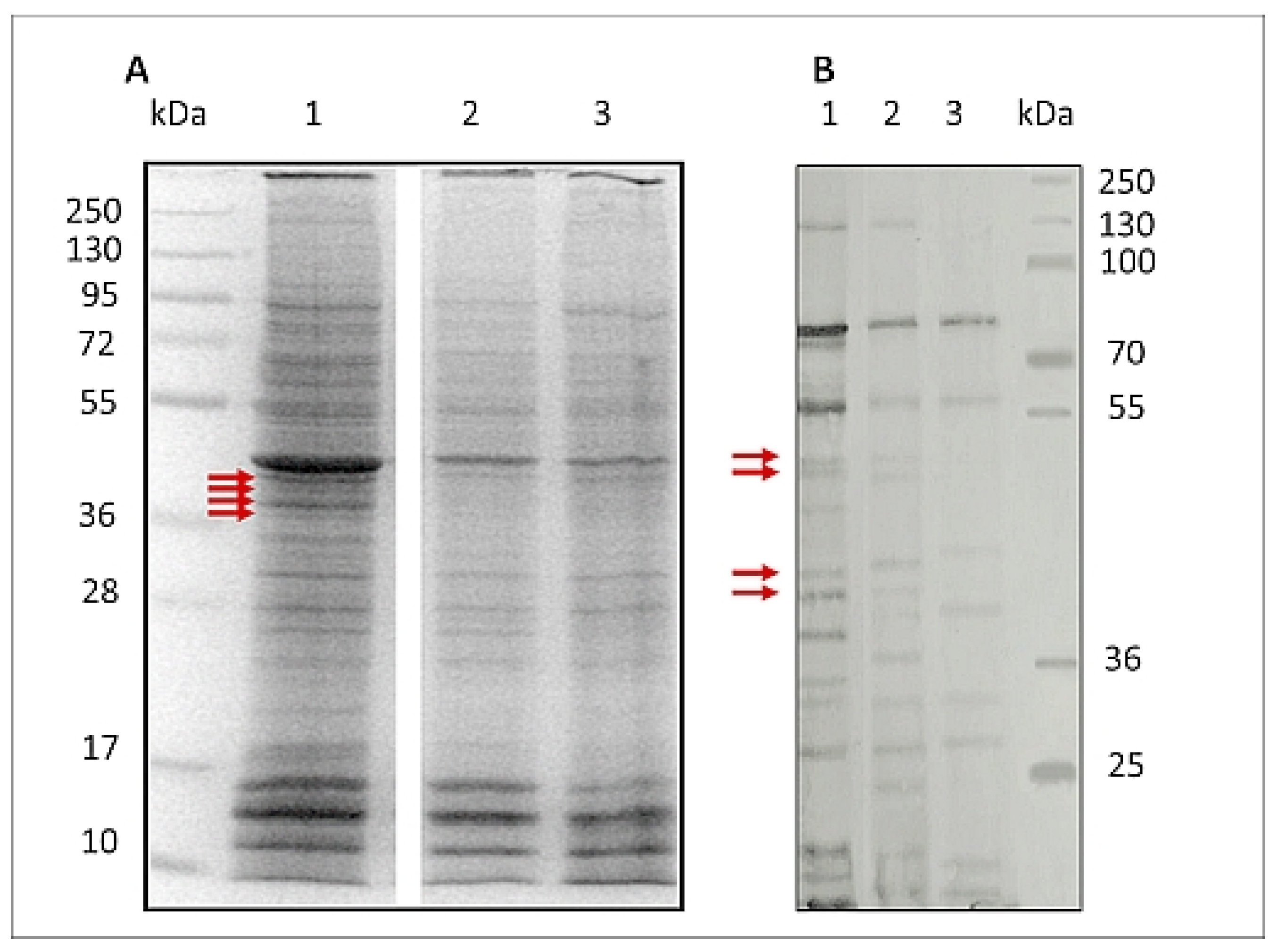
2.3. Marigold GlcUAOA Affects Antigenicity of L3 Proteins
2.4. LC-MS/MS Identification of Antigenic Proteins of L3 H. polygyrus bakeri
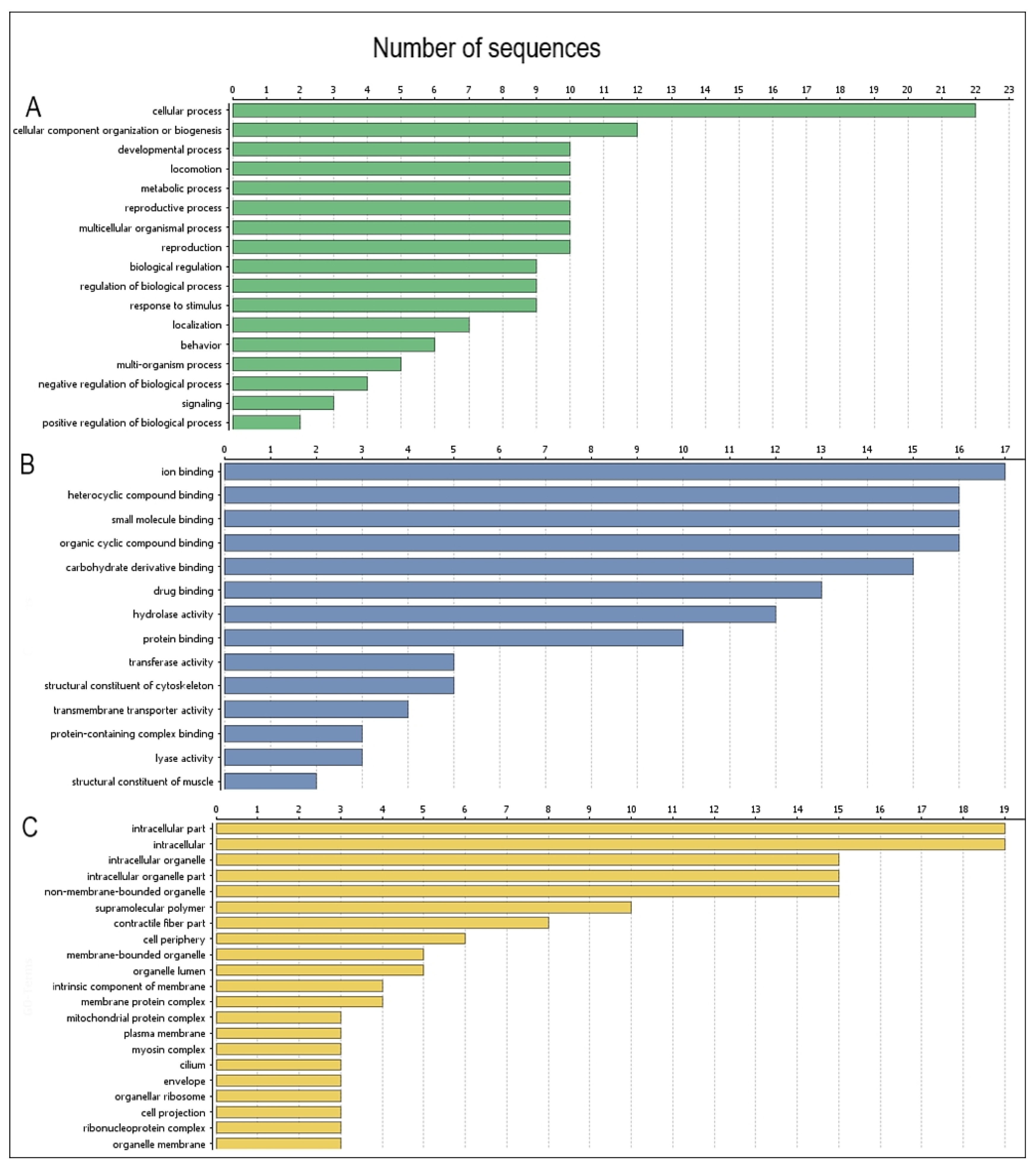
2.5. Functional Categories of Potentially Antigenic Proteins of L3 H. polygyrus bakeri by Gene Ontology Analysis
3. Discussion
4. Materials and Methods
4.1. Separation of Oleanolic Acid Glucuronides
4.2. Exposure of H. polygyrus bakeri to Oleanolic Acid Glucuronides
4.3. Evaluation of L3 by Transmission Electron Microscopy
4.4. Protein Sample Preparation and HPLC Profile
4.5. Preparation of L3 Protein Samples
4.6. Recognition of H. polygyrus bakeri L3 Immunogenic Proteins
4.7. LC-MS/MS Identification
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kerboeuf, D.; Riou, M.; Guegnard, F. Flavonids and related compounds in parasitic disease control. Mini Rev. Med. Chem. 2008, 8, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Quijada, J.; Fryganas, C.; Ropiak, H.M.; Ramsay, A.; Mueller-Harvey, I.; Hoste, H. Anthelmintic activities against Haemonchus contortus or Trichostrongylus colubriformis from small ruminants are influenced by structural features of condensed tannins. Agric. Food Chem. 2015, 63, 6346–6354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrckova, G.; Velebny, S. Pharmacological potential of selected natural compounds in the control of parasitic disease. In SpringerBriefs in Pharmaceutical Science and Drug Development; Springer: Heidelberg, Germany, 2013; pp. 101–125. [Google Scholar] [CrossRef]
- Maestrini, M.; Tava, A.; Mancini, S.; Tedesco, D.; Perrucci, S. In vitro anthelmintic activity of saponins from Medicago spp. against sheep gastrointestinal nematodes. Molecules 2020, 25, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klongsiriwet, C.; Quijada, J.; Williams, A.R.; Mueller-Harvey, I.; Williamson, E.M.; Hoste, H. Synergistic inhibition of Haemonchus contortus exsheathment by flavonoid monomers and condensed tannins. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 127–134. [Google Scholar] [CrossRef]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 2006, 22, 253–261. [Google Scholar] [CrossRef]
- Isah, M.; Ibrahim, M.; Mohammed, A.; Aliyu, A.; Masola, B.; Coetzer, T. A systematic review of pentacyclic triterpenes and their derivatives as chemotherapeutic agents against tropical parasitic diseases. Parasitology 2016, 143, 1219–1231. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Szakiel, A.; Ruszkowski, D.; Janiszowska, W. Saponins in Calendula officinalis L.—Structure, biosynthesis, transport and biological activity. Phytochem. Rev. 2005, 4, 151–158. [Google Scholar] [CrossRef]
- Reynolds, L.A.; Filbey, K.J.; Maizels, R.M. Immunity to the model intestinal helminth parasite Heligmosomoids polygyrus. Semin. Immunopathol. 2012, 34, 829–846. [Google Scholar] [CrossRef] [Green Version]
- Page, A.P.; Stepek, G.; Winter, A.D.; Pertab, D. Enzymology of the nematode cuticle: A potential drug target? Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Nollen, E.A.; Morimoto, R.I. Chaperoning signaling pathways: Molecular chaperones as stress-sensing ‘heat shock’ proteins. J. Cell Sci. 2002, 115, 2809–2816. [Google Scholar] [PubMed]
- Burnell, A.M.; Houthoofd, K.; O’Hanlon, K.; Vanfleteren, J.R. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Exp. Gerontol. 2005, 40, 850–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stear, M.J.; Doligalska, M.; Donskow-Schmelter, K. Alternatives to anthelmintics for the control of nematodes in livestock. Parasitology 2007, 134, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kerboeuf, D.; Guégnard, F. Anthelmintics are substrates and activators of nematode P glycoprotein. Antimicrob. Agents Chemother. 2011, 55, 2224–2232. [Google Scholar] [CrossRef] [Green Version]
- Issouf, M.; Guégnard, F.; Koch, C.; Le Vern, Y.; Blanchard-Letort, A.; Che, H.; Beech, R.N.; Kerboeuf, D.; Neveu, C. Haemonchus contortus P-glycoproteins interact with host eosinophil granules: A novel insight into the role of ABC transporters in host-parasite interaction. PLoS ONE 2014, 9, e87802. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, E.; Riou, M.; Kerboeuf, D.; Beugnet, F.; Chaminade, P.; Loiseau, P.M. Membrane lipidomics for the discovery of new antiparasitic drug targets. Trends Parasitol. 2011, 27, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Ey, P.L. Heigmosomoides polygyrus: Excretory/secretory antigens released in vitro by exsheathed third-stage larvae. Exp. Parasitol. 1988, 67, 210–220. [Google Scholar] [CrossRef]
- Doligalska, M.; Joźwicka, K.; Laskowska, M.; Donskow-Łysoniewska, K.; Pączkowski, C.; Janiszowska, W. Changes in Heligmosomoids polygyrus glycoprotein pattern by saponins impact the BALB/c mice immune response. Exp. Parasitol. 2013, 135, 524–531. [Google Scholar] [CrossRef]
- Hewitson, J.P.; Grainger, J.R.; Maizels, R.M. Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Mol. Biochem. Parasitol. 2009, 167, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, J.P.; Ivens, A.C.; Harcus, Y.; Filbey, K.J.; McSorley, H.J.; Murray, J.; Bridgett, S.; Ashford, D.; Dowle, A.A.; Maizels, R.M. Secretion of protective antigens by tissue-stage nematode larvae revealed by proteomic analysis and vaccination-induced sterile immunity. PLoS Pathog. 2013, 9, e1003492. [Google Scholar] [CrossRef] [Green Version]
- Hewitson, J.P.; Filbey, K.J.; Grainger, J.R.; Dowle, A.A.; Pearson, M.; Murray, J.; Harcus, Y.; Maizels, R.M. Heigmosomoides polygyrus elicits a dominant nonprotective antibody response directed against restricted glycan and peptide epitopes. J. Immunol. 2011, 187, 4764–4777. [Google Scholar] [CrossRef]
- Donskow-Łysoniewska, K.; Bien, J.; Brodaczewska, K.; Krawczak, K.; Doligalska, M. Colitis promotes adaptation of an intestinal nematode: A Heigmosomoides polygyrus mouse model system. PLoS ONE 2013, 8, e78034. [Google Scholar] [CrossRef] [PubMed]
- Donskow-Łysoniewska, K.; Krawczak, K.; Bocian, K.; Doligalska, M. The effects of intestinal nematode L4 stage on mouse experimental autoimmune encephalomyelitis. Arch. Immun. Ther. Exp. 2018, 66, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szakiel, A.; Ruszkowski, D.; Grudniak, A.; Kurek, A.; Wolska, K.I.; Doligalska, M.; Janiszowska, W. Antibacterial and antiparasitic activity of oleanolic acid and its glycosides isolated from marigold (Calendula officinalis). Planta Med. 2008, 74, 1709–1715. [Google Scholar] [CrossRef]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef]
- Johnstone, I.L. The cuticle of the nematode Caenorhabditis elegans: A complex collagen structure. Bioessays 1994, 16, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.H.; Bull, K.; Glautier, S.; Hopper, N.A.; Holden-Dye, L.; O’Connor, V. The concentration-dependent effects of ethanol on Caenorhabditis elegans behaviour. Pharm. J. 2007, 7, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Gopinath, A.; Reddy, S.M.; Madhan, B.; Shanmguam, G.; Rao, J.R. Effect of aqueous ethanol on the triple helical structure of collagen. Eur. Biophys. J. 2014, 43, 643–652. [Google Scholar] [CrossRef]
- Baumann, E.; Stoya, G.; Völkner, A.; Richter, W.; Lemke, C.; Linss, W. Hemolysis of human erythrocytes with saponin affects the membrane structure. Acta Histochem. 2000, 102, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Riou, M.; Guégnard, F.; Le Vern, Y.; Grasseau, I.; Koch, C.; Blesbois, E.; Kerboeuf, D. Effects of cholesterol content on activity of P-glycoproteins and membrane physical state, and consequences for anthelmintic resistance in the nematode Haemonchus contortus. Parasite 2020, 27, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harder, A. The biochemistry of Haemonchus contortus and other parasitic nematodes. Adv. Parasitol. 2016, 93, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Luz, A.L.; Lagido, C.; Hirschey, M.D.; Meyer, J.N. In vivo determination of mitochondrial function using luciferase-expressing Caenorhabditis elegans: Contribution of oxidative phosphorylation, glycolysis, and fatty acid oxidation to toxicant-induced dysfunction. Curr. Protoc. Toxicol. 2016, 69, 25.8.1–25.8.22. [Google Scholar] [CrossRef] [Green Version]
- Mathers, K.E.; James, F.; Staples, J.F. Saponin-permeabilization is not a viable alternative to isolated mitochondria for assessing oxidative metabolism in hibernation. Biol. Open 2015, 4, 858–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, P.; Singh, D.; Singh, M.M. Anti-Trichomonas activity of Sapindus saponins, a candidate for development as microbicidal contraceptive. J. Antimicrob. Chemother. 2008, 62, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.J.; Behm, C.A. Minireview: The role of the vacuolar ATPase in nematodes. Exp. Parasitol. 2012, 132, 47–55. [Google Scholar] [CrossRef]
- Hinton, A.; Bond, S.; Forgac, M. V-ATPase functions in normal and disease processes. Pflugers Arch. Eur. J. Physiol. 2009, 457, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, H.; Niesler, N.; Trautner, A.; Sama, S.; Jerz, G.; Panjideh, H.; Weng, A. Glycosylated triterpenoids as endosomal escape enhancers in targeted tumor therapies. Biomedicines 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.Q.; McNally, E.M. The dystrophin complex: Structure, function and implications for therapy. Compr. Physiol. 2015, 5, 1223–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, S. Regulation of structure and function of sarcomeric actin filaments in striated muscle of the nematode Caenorhabditis elegans. Anat. Rec. 2014, 297, 1548–1559. [Google Scholar] [CrossRef] [Green Version]
- Kwartler, C.S.; Chen, J.; Thakur, D.; Li, S.; Baskin, K.; Wang, S.; Wang, Z.V.; Walker, L.; Hill, J.A.; Epstein, H.F.; et al. Overexpression of smooth muscle myosin heavy chain leads to activation of the unfolded protein response and autophagic turnover of thick filament-associated proteins in vascular smooth muscle cells. J. Biol. Chem. 2014, 289, 14075–14088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, S. Vertebrate tropomyosin: Distribution, properties and function. J. Muscle Res. Cell Motil. 2001, 22, 5–49. [Google Scholar] [CrossRef]
- Nakada, T.; Nagano, I.; Wu, Z.; Takahashi, Y. Molecular cloning and expression of the full-length tropomyosin gene from Trichinella spiralis. J. Helminthol. 2003, 77, 57–63. [Google Scholar] [CrossRef]
- Ehsan, M.; Haseeb, M.; Hu, R.; Ali, H.; Memon, M.A.; Yan, R.; Xu, L.; Song, X.; Zhu, X.; Li, X. Tropomyosin: An excretory/secretory protein from Haemonchus contortus mediates the immuno-suppressive potential of goat peripheral blood mononuclear cells in vitro. Vaccines 2020, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, M.A.; Memon, M.A.; Jamil, T.; Naqvi, S.Z.; Aimulajiang, K.; Gadahi, J.A.; Xu, L.; Song, X.; Li, X.; Yan, R. Galectin domain containing protein from Haemonchus contortus modulates the immune functions of goat PBMCs and regulates CD4+ T-helper cells in vitro. Biomolecules 2020, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Ma, G.; Ang, C.S.; Korhonen, P.K.; Xu, R.; Nie, S.; Koehler, A.V.; Simpson, R.J.; Greening, D.W.; Reid, G.E.; et al. Somatic proteome of Haemonchus contortus. Int. J. Parasitol. 2019, 49, 311–320. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.S.; Wieseman, M.; Tropea, J.; Kaslow, D.; Abraham, D.; Lustigman, S.; Tuan, R.; Guderian, R.H.; Nutman, T.B. Onchocerca volvulus glycolytic enzyme fructose-1,6-bisphosphate aldolase as a target for a protective immune response in humans. Infect. Immun. 2002, 70, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Xu, M.J.; Nisbet, A.J.; Huang, C.Q.; Lin, R.Q.; Yuan, Z.G.; Song, H.Q.; Zhu, X.Q. Ascaris suum: RNAi mediated silencing of enolase gene expression in infective larvae. Exp. Parasitol. 2011, 127, 142–146. [Google Scholar] [CrossRef]
- Ayón-Núñez, D.A.; Fragoso, G.; Bobes, R.J.; Laclette, J.P. Plasminogen-binding proteins as an evasion mechanism of the host’s innate immunity in infectious diseases. Biosci. Rep. 2018, 38, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, M.B.; Ferguson, D.L. Haemonchus contortus: Enzymes III. Glutamate dehydrogenase. Exp. Parasitol. 1973, 34, 100–110. [Google Scholar] [CrossRef]
- Plaitakis, A.; Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Spanaki, C. The glutamate dehydrogenase pathway and its roles in cell and tissue biology in health and disease. Biology 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wloga, D.; Joachimiak, E.; Fabczak, H. Tubulin post-translational modifications and microtubule dynamics. Int. J. Mol. Sci. 2017, 18, 2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yutin, N.; Koonin, E.V. Archaeal origin of tubulin. Biol. Direct 2012, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felth, J. Studies of cytotoxic compounds of natural origin and their mechanisms of action. Acta Univ. Ups. 2011, 141, 56. [Google Scholar]
- Winterrowd, C.A.; Pomroy, W.E.; Sangster, N.C.; Johnson, S.S.; Geary, T.G. Benzimidazole-resistant β- tubulin alleles in a population of parasitic nematodes (Cooperia oncophora) of cattle. Vet. Parasitol. 2003, 117, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.I.; Wasmuth, J.D.; Beech, R.; Laing, R.; Hunt, M.; Naghra, H.; Cotton, J.A.; Berriman, M.; Britton, C.; Gilleard, J.S. Characterization and comparative analysis of the complete Haemonchus contortus β-tubulin gene family and implications for benzimidazole resistance in strongylid nematodes. Int. J. Parasitol. 2013, 43, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Goranova, T.E.; Bozhanov, S.S.; Lozanov, V.S.; Mitev, V.I.; Kaneva, R.P.; Georgieva, E.I. Changes in gene expression of CXCR4, CCR7 and BCL2 after treatment of breast cancer cells with saponin extract from Tribulus terrestris. Neoplasma 2015, 62, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, S.W. Antiparasitic drugs. In Small Animal Clinical Pharmacology, 2nd ed.; Saunders Ltd.: Nottingham, UK, 2008. [Google Scholar]
- Jordan, A.; Hadfield, J.A.; Lawrence, N.J.; McGown, A.T. Tubulin as a target for anticancer drugs: Agents which interact with the mitotic spindle. Med. Res. Rev. 1998, 18, 259–296. [Google Scholar] [CrossRef]
- Downing, K.H. Structural basis for the interaction of tubulin with proteins and drugs that affect microtubule dynamics. Ann. Rev. Cell Dev. Biol. 2000, 16, 89–111. [Google Scholar] [CrossRef]
- Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wei, S.Y.; Xu, B.; Guo, W.; Liu, D.L.; Cui, J.R.; Yao, X.S. Pro-apoptotic and microtubule-disassembly effects of ardisiacrispin (A+B), triterpenoid saponins from Ardisia crenata on human hepatoma Bel-7402 cells. J. Asian Nat. Prod. Res. 2008, 10, 739–746. [Google Scholar] [CrossRef]
- Rowinsky, E.K.; Donehower, R.C. Antimicrotubule agents. In Cancer Chemotherapy and Biotherapy; Chabner, B.A., Longo, D.L., Eds.; Lippincott, Williams & Wilkins: Philadelphia, PA, USA, 2001; pp. 329–372. [Google Scholar]
- Fang, Y.; Ferrie, A.M.; Li, G. Probing cytoskeleton modulation by optical biosensors. FEBS Lett. 2005, 579, 4175–4180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.R.; Fang, W.S. Pentacyclic triterpenoids and their saponins with apoptosis-inducing an activity. Curr. Top. Med. Chem. 2009, 9, 1581–1596. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Medina, E. A new extract of the plant Calendula officinalis produces a dual in vitro effect: Cytotoxic anti-tumor activity and lymphocyte activation. BMC Cancer 2006, 6, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunet, S.; Fourquaux, I.; Hoste, H. Ultrastructural changes in the third-stage, infective larvae of ruminant nematodes treated with sainfoin (Onobrychis viciifolia) extract. Parasitol. Int. 2011, 60, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.L.; Ji, P.F.; Kong, P.; Wang, Z.; Xiang, J.H.F. Arginine kinase from Litopenaeus vannamei: Cloning, expression and catalytic properties. Fish Shellfish Immunol. 2009, 26, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Hewitson, J.P.; Harcus, Y.; Murray, J.; Van Agtmaal, M.; Filbey, K.J.; Grainger, J.R.; Bridgett, S.; Blaxter, M.L.; Ashton, P.D.; Ashford, D.A.; et al. Proteomic analysis of secretory products from the model gastrointestinal nematode Heligmosomoids polygyrus reveals dominance of venom allergen-like (VAL) proteins. J. Proteom. 2011, 74, 1573–1594. [Google Scholar] [CrossRef] [Green Version]
- Ehsan, M.; Gao, W.X.; Gadahi, J.A.; Lu, M.M.; Liu, X.C.; Wang, Y.J.; Yan, R.F.; Xu, L.X.; Song, X.K.; Li, X.R. Arginine kinase from Haemonchus contortus decreased the proliferation and increased the apoptosis of goat PBMCs in vitro. Parasit. Vectors 2017, 10, 311. [Google Scholar] [CrossRef] [Green Version]
- Doligalska, M.; Jóźwicka, K.; Kiersnowska, M.; Mroczek, A.; Pączkowski, C.; Janiszowska, W. Triterpenoid saponins affect the function of P-glycoprotein and reduce the survival of the free-living stages of Heligmosomoids bakeri. Vet. Parasitol. 2011, 179, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Wahid, F.N.; Behnke, J.M. Stimuli for acquired resistance to Heligmosomoids polygyrus from intestinal tissue resident L3 and L4 larvae. Int. J. Parasitol. 1992, 22, 699–710. [Google Scholar] [CrossRef] [Green Version]
- Sulston, J.; Hodgkin, J. Methods. In The Nematode Caenorhabditis Elegans; Wood, W.B., Ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1988; pp. 587–606. [Google Scholar]
- Luft, J.H. Improvements in epoxy resin embedding methods. J. Biophys. Biochem. Cytol. 1961, 9, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.S. The use of lead citrate at high pH as electron-opaque stain for electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Doligalska, M.; Jóźwicka, K.; Donskow-Łysoniewska, K.; Kalinowska, M. The antiparasitic activity of avenacosides against intestinal nematodes. Vet. Parasitol. 2017, 241, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band (kDa) | Accession (Gi Number) | Protein Name | Species | Score | Mass (kDa) | emPAI | Coverage % | Number of Significant Peptide Matches | Uniprot ID | Description | Length (aa) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 and 2 (50.8-49.1) | gi|568290660 | ATP synthase F1, beta subunit | Necator americanus | 1325 | 58,151 | 1.57 | 0.45 | 28 | XP_013300832.1 | ATP synthase subunit beta, mitochondrial | 541 |
| gi|160415837 | Beta-tubulin isotype 1 | Cylicocyclus nassatus | 671 | 50,444 | 0.95 | 0.27 | 14 | ABX39216.1 | Beta-tubulin isotype 1 | 448 | |
| gi|560120094 | Myosin and Myosin head and Myosin tail domain containing protein | Haemonchus contortus | 644 | 226,702 | 0.28 | 0.09 | 15 | CDJ95285.1 | Myosin head | 1963 | |
| gi|47606682 | Paramyosin isoform 1 | Dictyocaulus viviparus | 620 | 100,878 | 0.65 | 0.22 | 13 | AAT36324.1 | CRE-UNC-15 protein | 876 | |
| gi|597882604 | Hypothetical protein Y032_0003g1332 | Ancylostoma ceylanicum | 492 | 50,904 | 0.79 | 0.3 | 9 | EYC31965.1 | V-type ATPase, B subunit | 458 | |
| gi|371486394 | Alpha tubulin | Ostertagia ostertagi | 465 | 5053 | 0.79 | 0.24 | 11 | AEX31242.1 | Tubulin/FtsZ family, GTPase domain protein | 446 | |
| gi|568269548 | Carboxyl transferase domain protein | Necator americanus | 321 | 58,645 | 0.24 | 0.11 | 6 | ETN70891 | Carboxyl transferase domain protein | 535 | |
| gi|9438176 | Heat shock 70 protein | Parastrongyloides trichosuri | 318 | 70,452 | 0.35 | 0.17 | 6 | AAF87583.1 | Heat shock protein 70 | 644 | |
| gi|162280611 | Actin variant 1 | Dictyocaulus viviparus | 274 | 42,129 | 0.65 | 0.29 | 5 | ABX82966.1 | Actin | 376 | |
| gi|253721983 | Glutamate dehydrogenase, partial | Haemonchus contortus | 210 | 59,444 | 0.33 | 0.09 | 5 | ACT34055.1 | Glutamate dehydrogenase | 532 | |
| 3 (42.7) | gi|162280611 | Actin variant 1 | Dictyocaulus viviparus | 2471 | 42,129 | 2.31 | 0.46 | 57 | ABX82966.1 | Actin | 376 |
| gi|560127002 | Lipoma HMGIC fusion partner protein and Heat shock protein 70 domain containing protein | Haemonchus contortus | 771 | 9216 | 0.66 | 0.19 | 17 | CDJ88360.1 | Putative chaperone protein DnaK | 829 | |
| gi|301015486 | Enolase | Haemonchus contortus | 497 | 47,273 | 1.04 | 0.3 | 9 | ADK47524.1 | Phosphopyruvate hydratase | 434 | |
| gi|597877220 | Hypothetical protein Y032_0010g849 | Ancylostoma ceylanicum | 374 | 62,832 | 0.6 | 0.11 | 9 | EYC26588.1 | Protein unc-87 | 558 | |
| gi|47606682 | Paramyosin isoform 1 | Dictyocaulus viviparus | 310 | 100,878 | 0.18 | 0.07 | 6 | AAT36324.1 | CRE-UNC-15 protein | 876 | |
| gi|157326537 | Tropomyosin | Heigmosomoides polygyrus | 291 | 3313 | 1.14 | 0.18 | 6 | ABV44405.1 | Tropomyosin | 284 | |
| gi|568290660 | ATP synthase F1, beta subunit | Necator americanus | 217 | 58,151 | 0.34 | 0.19 | 4 | XP_013300832.1 | ATP synthase subunit beta, mitochondrial | 541 | |
| 4 (40.7) | gi|325516326 | Disorganized muscle protein 1 | Haemonchus contortus | 414 | 35,825 | 0.26 | 0.14 | 5 | ADZ24723.1 | Disorganized muscle protein 1 | 321 |
| gi|162280611 | Actin variant 1 | Dictyocaulus viviparus | 411 | 42,129 | 0.82 | 0.37 | 10 | ABX82966.1 | Actin | 376 | |
| gi|560124746 | Galectin domain containing protein | Haemonchus contortus | 333 | 40,642 | 0.23 | 0.1 | 9 | CDJ90629.1 | Galactoside-binding lectin | 356 | |
| gi|541045947 | Fructose-bisphosphate aldolase 2 | Ascaris suum | 316 | 45,469 | 0.1 | 0.06 | 6 | ADY45824.1 | Fructose-bisphosphate aldolase class-I | 417 | |
| gi|597874384 | Hypothetical protein Y032_0015g2816 | Ancylostoma ceylanicum | 295 | 40,719 | 0.36 | 0.11 | 7 | EYC23757.1 | Arginine kinase | 359 | |
| gi|568290660 | ATP synthase F1, beta subunit | Necator americanus | 255 | 58,151 | 0.55 | 0.24 | 6 | XP_013300832.1 | ATP synthase subunit beta, mitochondrial | 541 | |
| gi|157381686 | Heat shock protein 70 | Angiostrongylus vasorum | 253 | 5614 | 0.35 | 0.13 | 5 | ABV46675.1 | Putative chaperone protein DnaK | 507 | |
| gi|597834320 | Hypothetical protein Y032_0324g2520 | Ancylostoma ceylanicum | 228 | 30,314 | 0.15 | 0.09 | 4 | EYB84003.1 | Fructose-bisphosphate aldolase class-I | 282 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doligalska, M.; Jóźwicka, K.; Szewczak, L.; Nowakowska, J.; Brodaczewska, K.; Goździk, K.; Pączkowski, C.; Szakiel, A. Calendula officinalis Triterpenoid Saponins Impact the Immune Recognition of Proteins in Parasitic Nematodes. Pathogens 2021, 10, 296. https://doi.org/10.3390/pathogens10030296
Doligalska M, Jóźwicka K, Szewczak L, Nowakowska J, Brodaczewska K, Goździk K, Pączkowski C, Szakiel A. Calendula officinalis Triterpenoid Saponins Impact the Immune Recognition of Proteins in Parasitic Nematodes. Pathogens. 2021; 10(3):296. https://doi.org/10.3390/pathogens10030296
Chicago/Turabian StyleDoligalska, Maria, Kinga Jóźwicka, Ludmiła Szewczak, Julita Nowakowska, Klaudia Brodaczewska, Katarzyna Goździk, Cezary Pączkowski, and Anna Szakiel. 2021. "Calendula officinalis Triterpenoid Saponins Impact the Immune Recognition of Proteins in Parasitic Nematodes" Pathogens 10, no. 3: 296. https://doi.org/10.3390/pathogens10030296
APA StyleDoligalska, M., Jóźwicka, K., Szewczak, L., Nowakowska, J., Brodaczewska, K., Goździk, K., Pączkowski, C., & Szakiel, A. (2021). Calendula officinalis Triterpenoid Saponins Impact the Immune Recognition of Proteins in Parasitic Nematodes. Pathogens, 10(3), 296. https://doi.org/10.3390/pathogens10030296