
Evaluation of “Caterina assay”: An Alternative Tool to the Commercialized Kits Used for Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Identification

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
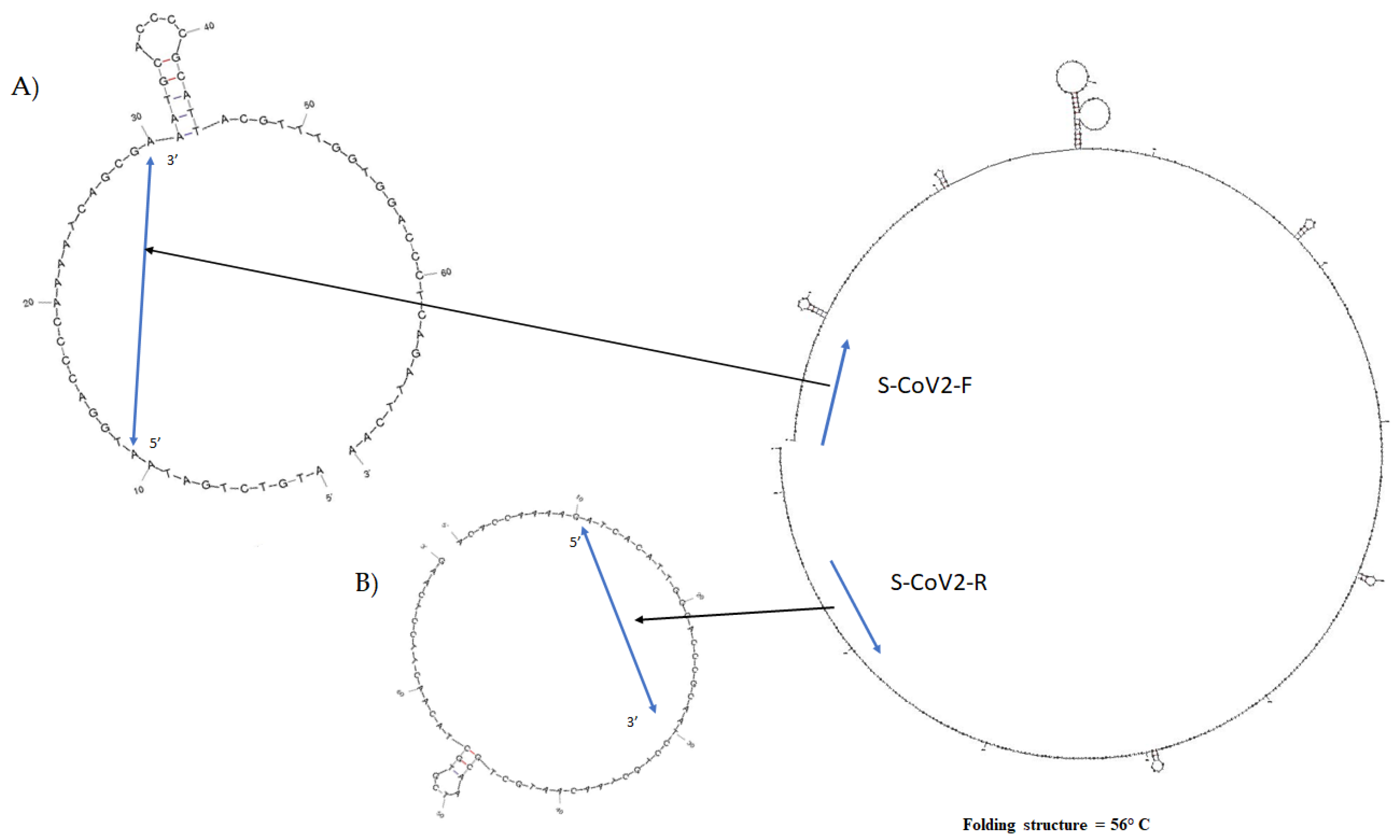
2.1. SARS-CoV-2 rRT-PCR In Silico Design
2.2. Positive Control
2.3. RNA Extraction
2.4. RNA Extraction Internal Controls
2.5. Real-Time rt-PCR Quantitative Conditions
2.6. cDNA Sequencing
2.7. Clinical Samples
2.8. Statistical Analysis
3. Results
3.1. Primer Design
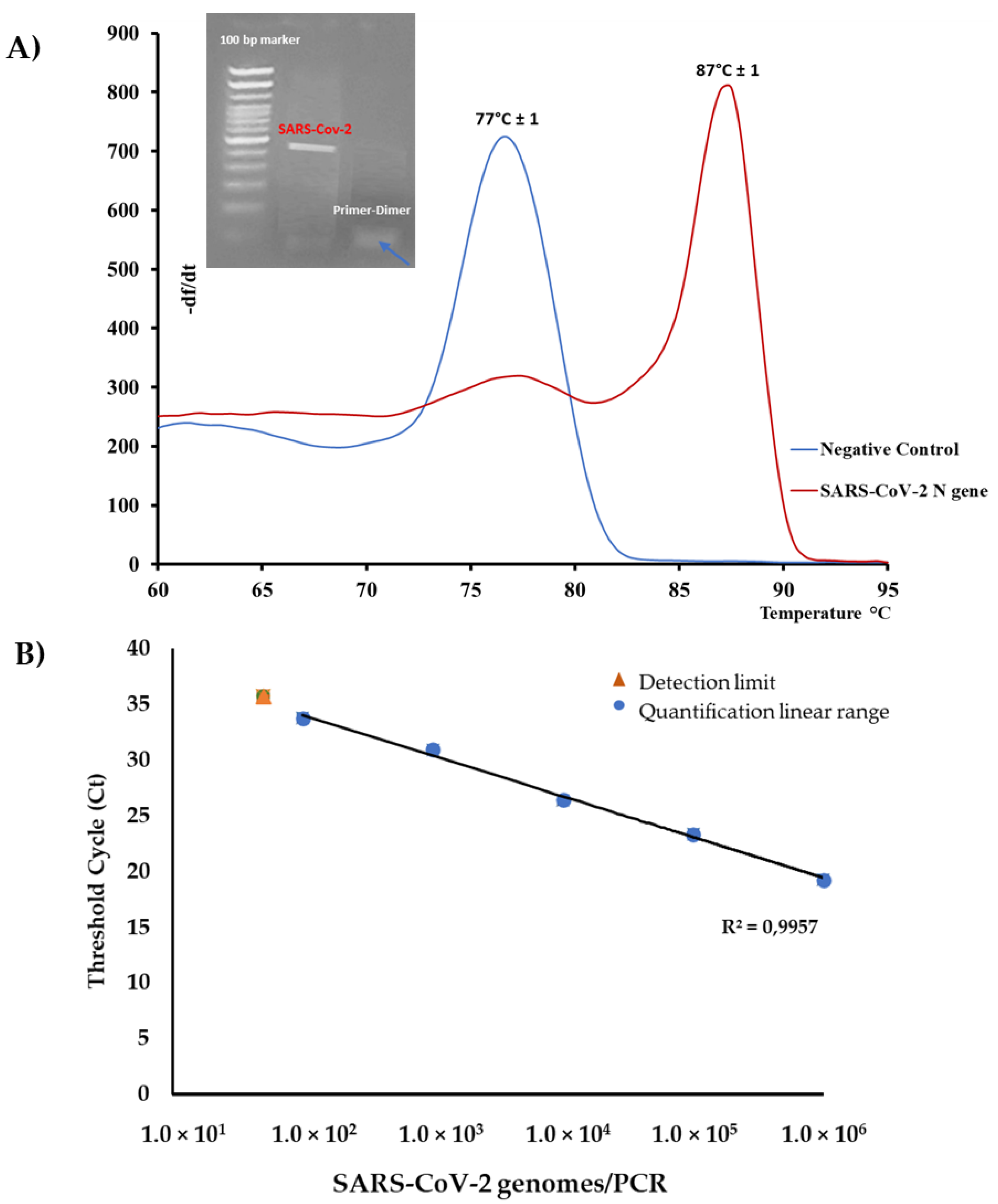
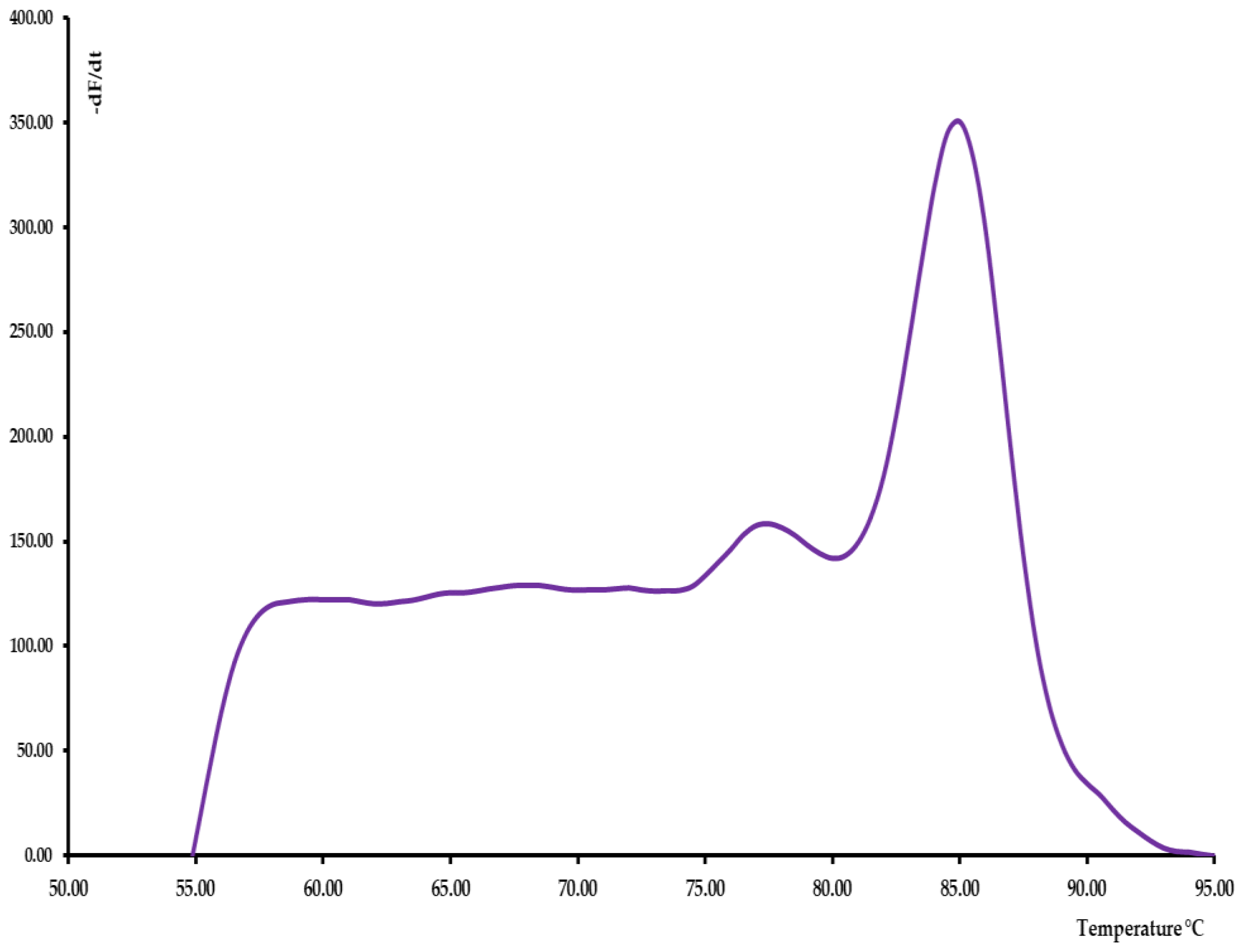
3.2. SYBR®Green rRT-PCR Melting Curve Analysis
3.3. Sensitivity of the Method
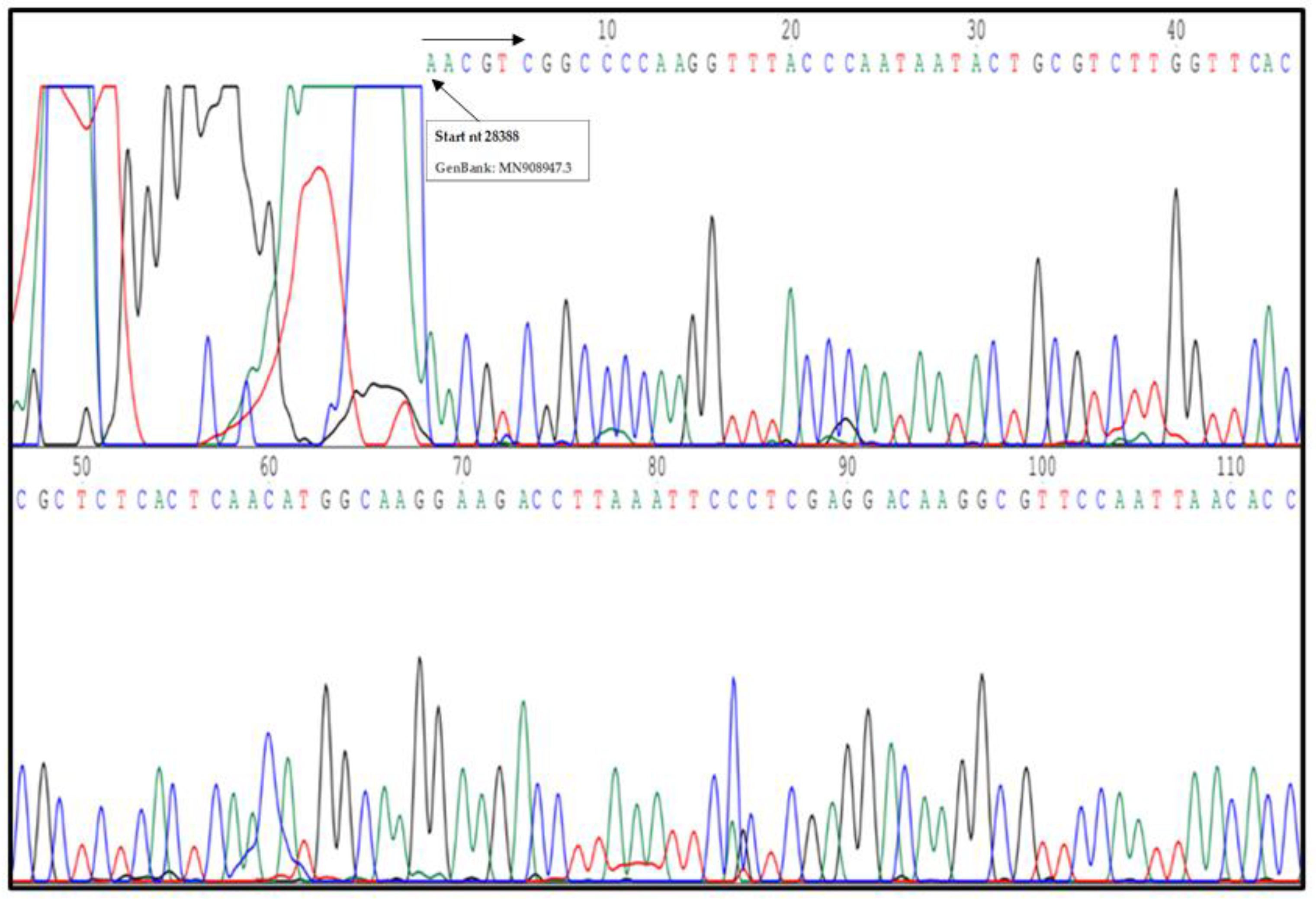
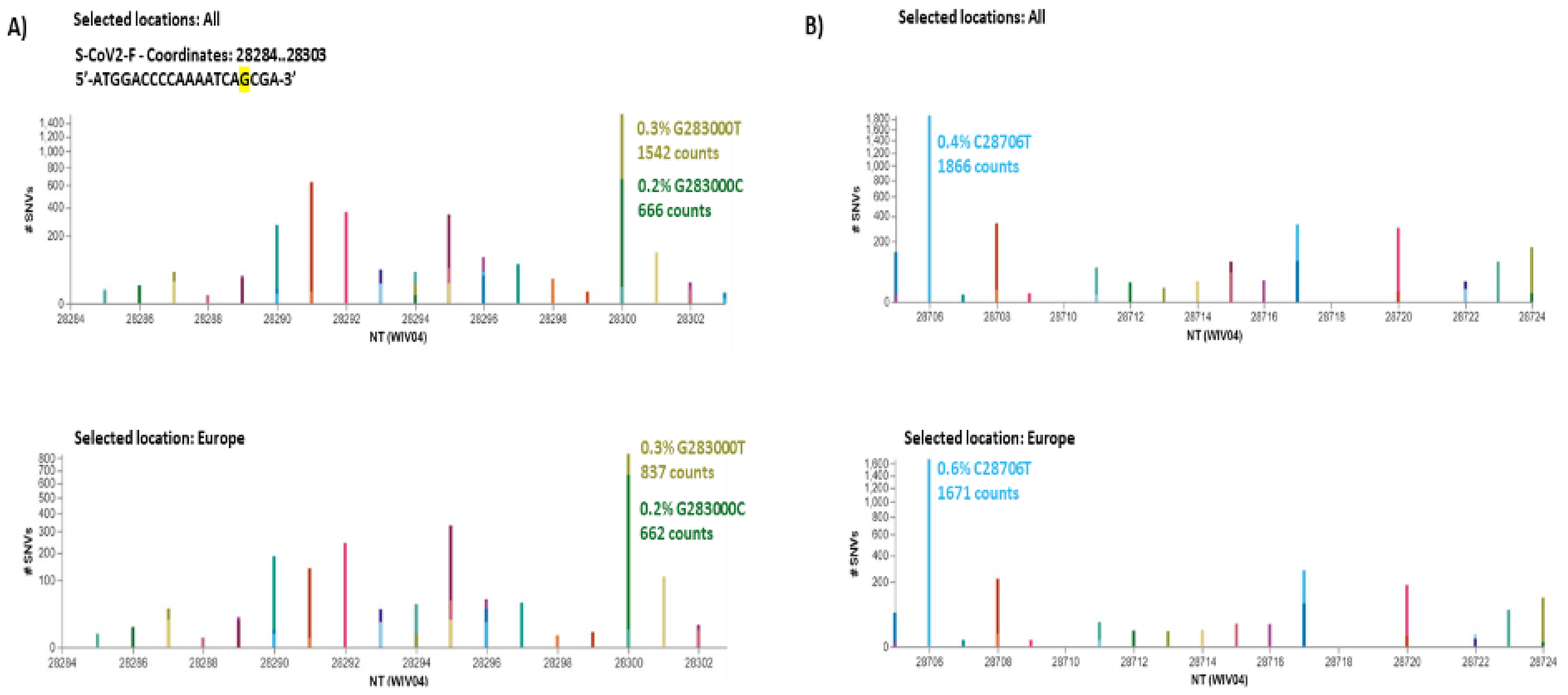
3.4. SARS-CoV-2 N Protein Identification by Sequencing
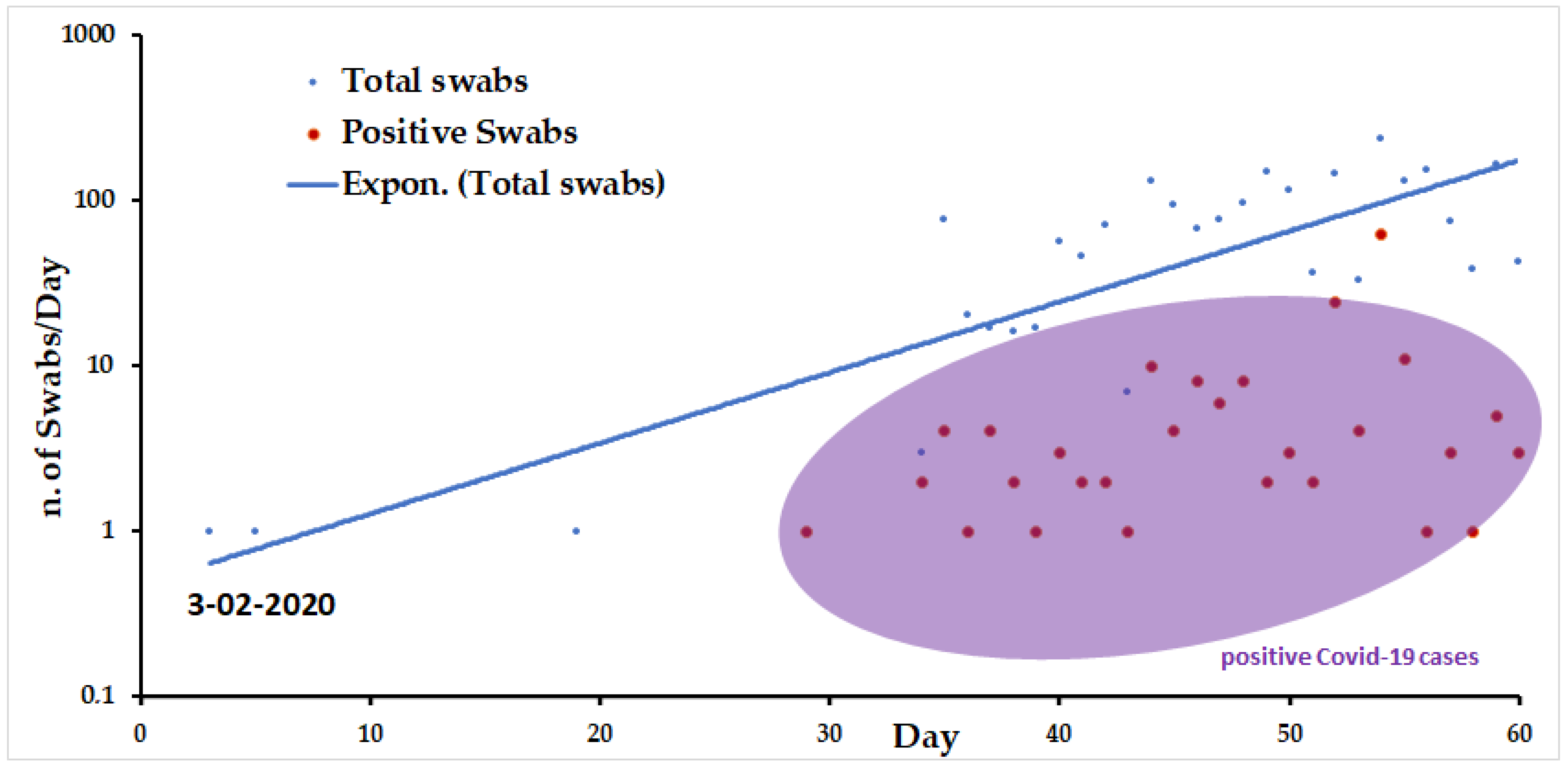
3.5. Detection of SARS-CoV-2 on Clinical Samples from a Court of Sardinian Patients
3.6. PCR Validation Method: Comparison of Commercial Kit and the Homemade Procedure
4. Discussion
5. Conclusions
Recommendation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Carta, M.G.; Orrù, G.; Scano, A.; Coghe, F.; Nunnari, G.; Facchini, G.; Numis, F.G.; Berretta, M. In the face of the SARS-CoV-2 outbreak, do people suffering from oncological disease need specific attention? Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3434–3436. [Google Scholar] [CrossRef] [PubMed]
- Saban, O.; Levy, J.; Chowers, I. Risk of SARS-CoV-2 transmission to medical staff and patients from an exposure to a COVID-19-positive ophthalmologist. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 2271–2274. [Google Scholar] [CrossRef]
- Ferrando, M.L.; Coghe, F.; Scano, A.; Carta, M.G.; Orrù, G. Co-infection of Streptococcus pneumoniae in Respiratory Infections Caused by SARS-CoV-2. Biointerface Res. Appl. Chem. 2021, 11, 12170–12177. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Godri Pollitt, K.J.; Peccia, J.; Ko, A.I.; Kaminski, N.; Dela Cruz, C.S.; Nebert, D.W.; Reichardt, J.K.V.; Thompson, D.C.; Vasiliou, V. COVID-19 vulnerability: The potential impact of genetic susceptibility and airborne transmission. Hum. Genom. 2020, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- WHO Weekly Epidemiological Update—29 December 2020. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update (accessed on 29 December 2020).
- Lu, H.; Stratton, C.W.; Tang, Y. The Wuhan SARS-CoV-2—What’s next for China. J. Med. Virol. 2020, 92, 546–547. [Google Scholar] [CrossRef] [Green Version]
- Galli, C.; Plebani, M. Clinical laboratory and SARS-CoV-2 infection: Where do we stand? Clin. Chem. Lab. Med. 2020, 58, 1139–1141. [Google Scholar] [CrossRef] [Green Version]
- Mathuria, J.P.; Yadav, R. Rajkumar Laboratory diagnosis of SARS-CoV-2—A review of current methods. J. Infect. Public Health 2020, 13, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.M.; Abramson, R.D.; Watson, R.; Gelfand, D.H. Detection of specific polymerase chain reaction product by utilizing the 5′----3′ exonuclease activity of Thermus aquaticus DNA polymerase. Proc. Natl. Acad. Sci. USA 1991, 88, 7276–7280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention CDC 2019-Novel Coronavirus (2019-nCoV) Real-Time RT-PCR Diagnostic Panel. CDC-006-00005; 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/lab/rt-pcr-panel-primer-probes.html (accessed on 21 January 2020).
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corman, V.; Bleicker, T.; Brunink, S.; Drosten, C. Diagnostic detection of Wuhan coronavirus 2019 by real-time RT-PCR. Public Health Engl. 2020, 25, 2000045. [Google Scholar] [CrossRef] [Green Version]
- Moore, N.M.; Li, H.; Schejbal, D.; Lindsley, J.; Hayden, M.K. Comparison of Two Commercial Molecular Tests and a Laboratory-Developed Modification of the CDC 2019-nCoV Reverse Transcriptase PCR Assay for the Detection of SARS-CoV-2. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Iglói, Z.; Leven, M.; Abdel-Karem Abou-Nouar, Z.; Weller, B.; Matheeussen, V.; Coppens, J.; Koopmans, M.; Molenkamp, R. Comparison of commercial realtime reverse transcription PCR assays for the detection of SARS-CoV-2. J. Clin. Virol. 2020, 129, 104510. [Google Scholar] [CrossRef]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carta, M.G.; Scano, A.; Lindert, J.; Bonanno, S.; Rinaldi, L.; Fais, S.; Orrù, G. Association between the spread of COVID-19 and weather-climatic parameters. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8226–8231. [Google Scholar] [CrossRef]
- Arcadu, B.; Orrù, M.; Piga, R.; Orrù, G. Designing of sequencing assay assisted by capillary electrophoresis based on DNA folding analysis: An application to the VCAM1 gene. Electrophoresis 2012, 33, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Orrù, G.; Ferrando, M.L.; Meloni, M.; Liciardi, M.; Savini, G.; De Santis, P. Rapid detection and quantitation of Bluetongue virus (BTV) using a Molecular Beacon fluorescent probe assay. J. Virol. Methods 2006, 137, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Sumner, K.; Swensen, J.J.; Procter, M.; Jama, M.; Wooderchak-Donahue, W.; Lewis, T.; Fong, M.; Hubley, L.; Schwarz, M.; Ha, Y.; et al. Noncontinuously Binding Loop-Out Primers for Avoiding Problematic DNA Sequences in PCR and Sanger Sequencing. J. Mol. Diagn. 2014, 16, 477–480. [Google Scholar] [CrossRef] [PubMed]
- WHO Announces COVID-19 Outbreak a Pandemic. Available online: https://www.euro.who.int/en/health-topics/health-emergencies/coronavirus-covid-19/news/news/2020/3/who-announces-covid-19-outbreak-a-pandemic (accessed on 19 February 2021).
- Petrillo, S.; Carrà, G.; Bottino, P.; Zanotto, E.; De Santis, M.C.; Margaria, J.P.; Giorgio, A.; Mandili, G.; Martini, M.; Cavallo, R.; et al. A Novel Multiplex qRT-PCR Assay to Detect SARS-CoV-2 Infection: High Sensitivity and Increased Testing Capacity. Microorganisms 2020, 8, 1064. [Google Scholar] [CrossRef] [PubMed]
- Zhen, W.; Berry, G.J. Development of a New Multiplex Real-Time RT-PCR Assay for Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Detection. J. Mol. Diagn. 2020, 22, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Kashir, J.; Yaqinuddin, A. Loop mediated isothermal amplification (LAMP) assays as a rapid diagnostic for COVID-19. Med. Hypotheses 2020, 141, 109786. [Google Scholar] [CrossRef]
- Zhang, Y.-Z. Novel 2019 coronavirus genome. Virological. Org. 2020. Available online: https://virological.org/t/initial-assessment-of-the-ability-of-published-coronavirus-primers-sets-to-detect-the-wuhan-coronavirus/321 (accessed on 21 January 2020).
- Li, D.; Zhang, J.; Li, J. Primer design for quantitative real-time PCR for the emerging Coronavirus SARS-CoV-2. Theranostics 2020, 10, 7150–7162. [Google Scholar] [CrossRef]
- Dorlass, E.G.; Monteiro, C.O.; Viana, A.O.; Soares, C.P.; Machado, R.R.G.; Thomazelli, L.M.; Araujo, D.B.; Leal, F.B.; Candido, E.D.; Telezynski, B.L.; et al. Lower cost alternatives for molecular diagnosis of COVID-19: Conventional RT-PCR and SYBR Green-based RT-qPCR. Braz. J. Microbiol. 2020, 51, 1117–1123. [Google Scholar] [CrossRef] [PubMed]
- Guglielmotti, M.B.; Olmedo, D.G.; Cabrini, R.L. Research on implants and osseointegration. Periodontology 2000, 79, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.W.; Pan, Y.; Cheng, S.M.S.; Hui, K.P.Y.; Krishnan, P.; Liu, Y.; Ng, D.Y.M.; Wan, C.K.C.; Yang, P.; Wang, Q.; et al. Molecular Diagnosis of a Novel Coronavirus (2019-nCoV) Causing an Outbreak of Pneumonia. Clin. Chem. 2020, 66, 549–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Qavi, A.J.; Hachim, A.; Kavian, N.; Cole, A.R.; Moyle, A.B.; Wagner, N.D.; Sweeney-Gibbons, J.; Rohrs, H.W.; Gross, M.L.; et al. Characterization of SARS-CoV-2 N protein reveals multiple functional consequences of the C-terminal domain. bioRxiv 2020. [Google Scholar] [CrossRef]
- Surjit, M.; Lal, S.K. The SARS-CoV nucleocapsid protein: A protein with multifarious activities. Infect. Genet. Evol. 2008, 8, 397–405. [Google Scholar] [CrossRef]
- Moreira, R.A.; Guzman, H.V.; Boopathi, S.; Baker, J.L.; Poma, A.B. Characterization of Structural and Energetic Differences between Conformations of the SARS-CoV-2 Spike Protein. Materials 2020, 13, 5362. [Google Scholar] [CrossRef]
- Vilar, S.; Isom, D.G. One Year of SARS-CoV-2: How Much Has the Virus Changed? Biology 2021, 10, 91. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.T.; Altschuler, K.; Zhan, S.H.; Chan, Y.A.; Deverman, B.E. COVID-19 CG: Tracking SARS-CoV-2 mutations by locations and dates of interest. bioRxiv 2020. [Google Scholar] [CrossRef]
- CDC, Centers for Disease Control and Emerging SARS-CoV-2 Variants. Available online: https://www.cdc.gov/coronavirus/2019-ncov/more/science-and-research/scientific-brief-emerging-variants.html (accessed on 20 February 2021).
- Carta, M.G.; Romano, F.; Orrù, G. The True Challenges of the Covid-19 Epidemics: The Need for Essential Levels of Care for All. Open Respir. Med. J. 2020. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 Genesig Real-Time PCR AssayTM | DA An Gene (2019-nCoV) RNATM | |||||
|---|---|---|---|---|---|---|
| Caterina Assay | Detected | No Detected | Kappa (±95% CI) * | Detected | No Detected | Kappa (±95% CI) * |
| Detected | 110 | 3 | 0.971 (0.939–1.0) | 112 | 1 | 0.990 (0.972–1.0) |
| No Detected | 0 | 97 | 0 | 97 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orrù, G.; Scano, A.; Fais, S.; Loddo, M.; Carta, M.G.; Steri, G.C.; Santus, S.; Cappai, R.; Ferrando, M.L.; Coghe, F. Evaluation of “Caterina assay”: An Alternative Tool to the Commercialized Kits Used for Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Identification. Pathogens 2021, 10, 325. https://doi.org/10.3390/pathogens10030325
Orrù G, Scano A, Fais S, Loddo M, Carta MG, Steri GC, Santus S, Cappai R, Ferrando ML, Coghe F. Evaluation of “Caterina assay”: An Alternative Tool to the Commercialized Kits Used for Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Identification. Pathogens. 2021; 10(3):325. https://doi.org/10.3390/pathogens10030325
Chicago/Turabian StyleOrrù, Germano, Alessandra Scano, Sara Fais, Miriam Loddo, Mauro Giovanni Carta, Giorgio Carlo Steri, Simonetta Santus, Riccardo Cappai, Maria Laura Ferrando, and Ferdinando Coghe. 2021. "Evaluation of “Caterina assay”: An Alternative Tool to the Commercialized Kits Used for Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Identification" Pathogens 10, no. 3: 325. https://doi.org/10.3390/pathogens10030325
APA StyleOrrù, G., Scano, A., Fais, S., Loddo, M., Carta, M. G., Steri, G. C., Santus, S., Cappai, R., Ferrando, M. L., & Coghe, F. (2021). Evaluation of “Caterina assay”: An Alternative Tool to the Commercialized Kits Used for Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Identification. Pathogens, 10(3), 325. https://doi.org/10.3390/pathogens10030325