
Molecular Characterisation of Cryptosporidium spp. in Mozambican Children Younger than 5 Years Enrolled in a Matched Case-Control Study on the Aetiology of Diarrhoeal Disease

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
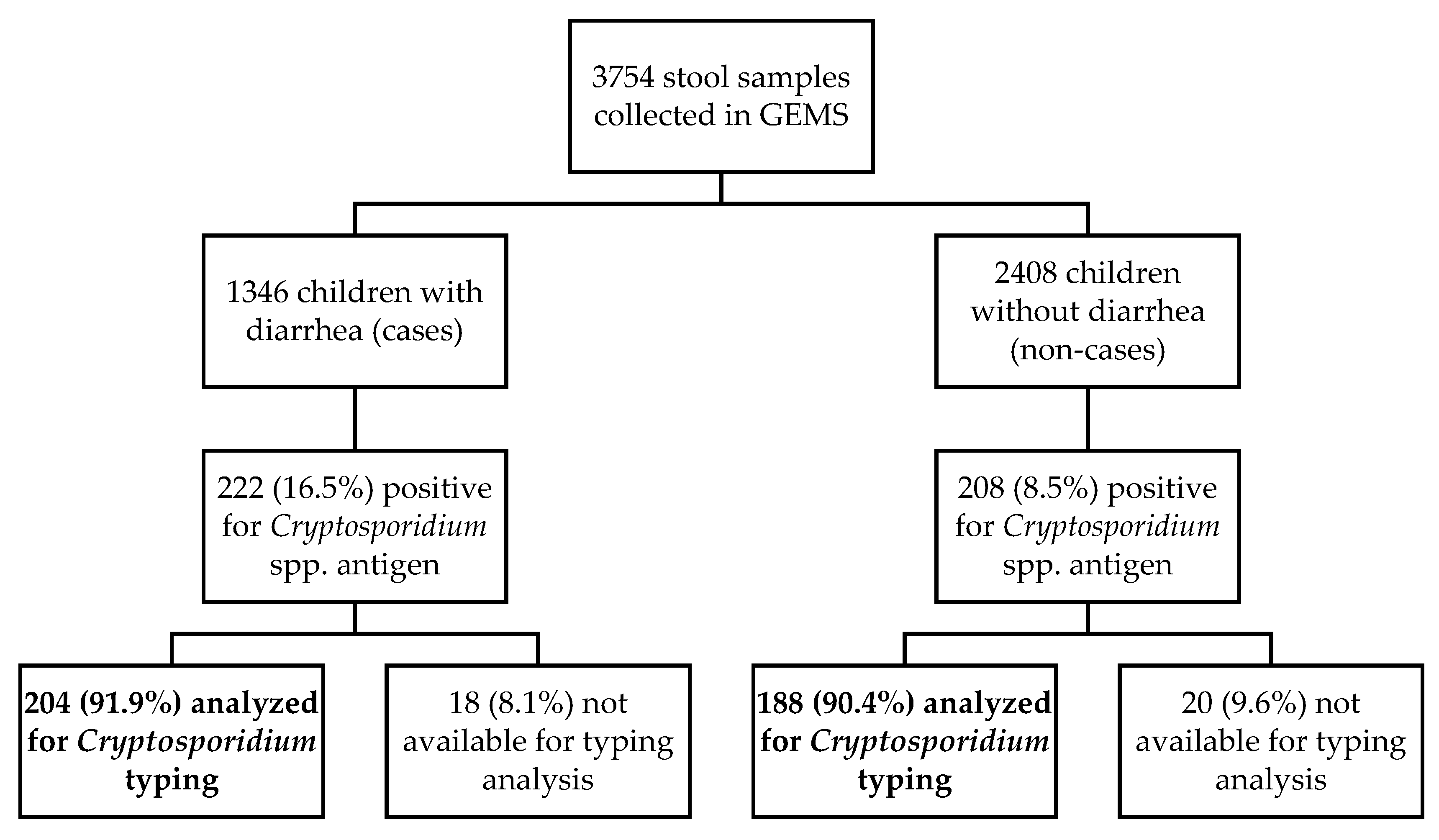
2.1. Initial Screening for the Detection of Cryptosporidium spp. by ELISA Immunoassay
2.2. Confirmation of Cryptosporidium spp. by Nested PCR Methods
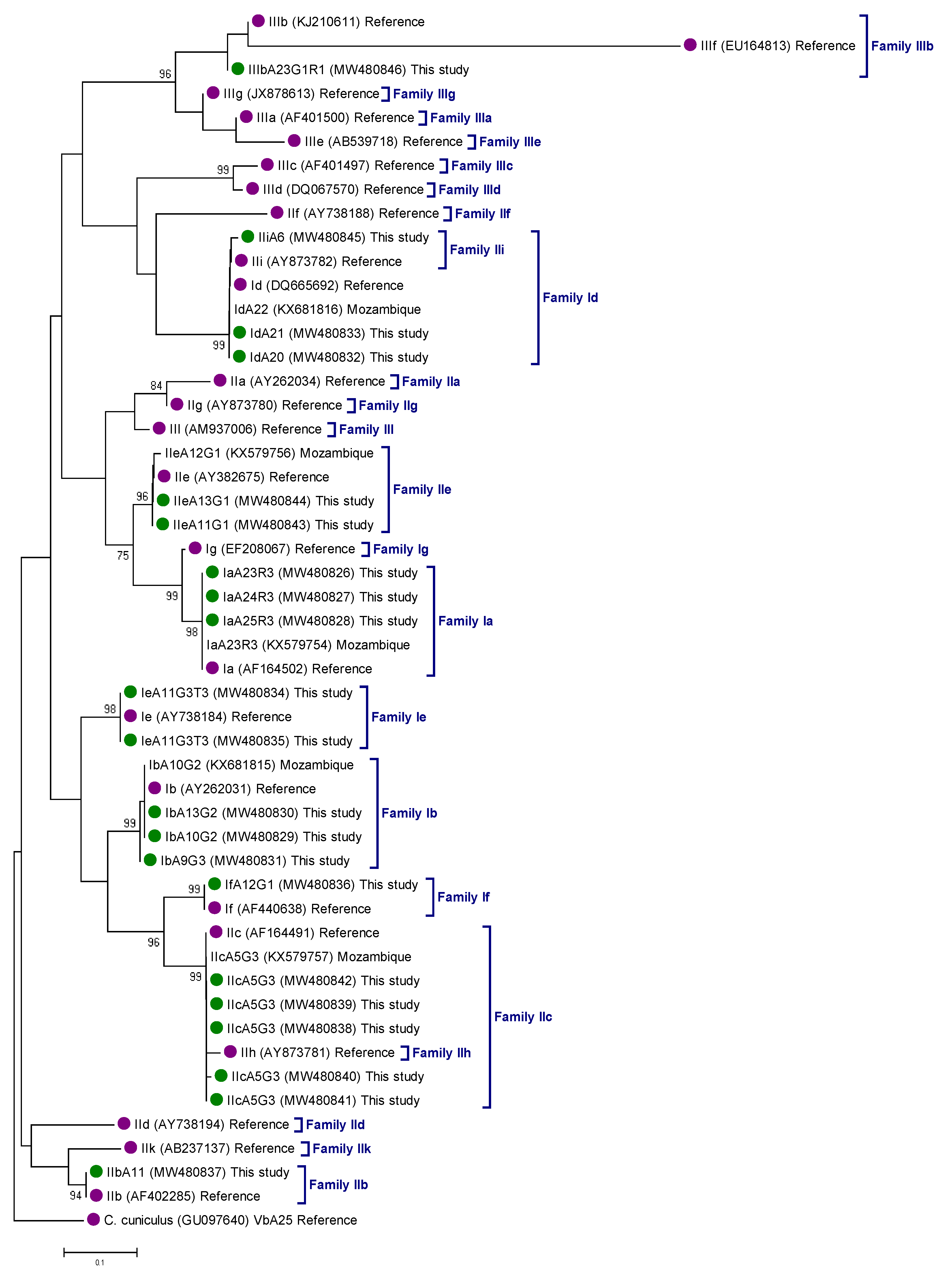
2.3. Genetic Variation within C. hominis and C. parvum
3. Discussion
4. Materials and Methods
4.1. Study Context
4.2. Stool Collection and Initial Testing
4.3. Molecular Study
4.3.1. DNA Extraction and Purification
4.3.2. Molecular Detection and Characterisation of Cryptosporidium spp.
4.4. Data Analysis
4.4.1. Epidemiological Analysis
4.4.2. Sequence and Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2015 Mortality and Causes of Death Collaborators. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- GBD 2015 Child Mortality Collaborators. Global, regional, national, and selected subnational levels of stillbirths, neonatal, infant, and under-5 mortality, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1725–1774. [Google Scholar] [CrossRef] [Green Version]
- GBD Diarrhoeal Diseases Collaborators. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef] [Green Version]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.; Panchalingham, S.; Sow, S.O.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G.; et al. The incidence, aetiology, and adverse clinical consequences of less severe diarrhoeal episodes among infants and children residing in low-income and middle-income countries: A 12-month case-control study as a follow-on to the Global Enteric Multicenter Study (GEMS). Lancet Glob. Health 2019, 7, e568–e584. [Google Scholar] [PubMed] [Green Version]
- Steiner, K.L.; Ahmed, S.; Gilchrist, C.A.; Burkey, C.; Cook, H.; Ma, J.Z.; Korpe, P.S.; Ahmed, E.; Alam, M.; Kabir, M.; et al. Species of Cryptosporidia causing subclinical infection associated with growth faltering in rural and urban Bangladesh: A birth cohort study. Clin. Infect. Dis. 2018, 67, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. MAL-ED Network Investigators. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef] [Green Version]
- Bouzid, M.; Hunter, P.R.; Chalmers, R.M.; Tyler, K.M. Cryptosporidium pathogenicity and virulence. Clin. Microbiol. Rev. 2013, 26, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Ehmadpour, E.; Safarpour, H.; Xiao, L.; Zarean, M.; Hatam-Nahavandi, K.; Barac, A.; Picot, S.; Rahimi, M.T.; Rubino, S.; Mahami-Oskouei, M.; et al. Cryptosporidiosis in HIV-positive patients and related risk factors: A systematic review and meta-analysis. Parasite 2020, 27, 27. [Google Scholar] [CrossRef]
- Xiao, L. Molecular epidemiology of cryptosporidiosis: An update. Exp. Parasitol. 2010, 124, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasit. Vectors 2017, 10, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium species in humans and animals: Current understanding and research needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Ryan, U.; Xiao, L. Genetic diversity and population structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef]
- Strong, W.B.; Gut, J.; Nelson, R.G. Cloning and sequence analysis of a highly polymorphic Cryptosporidium parvum gene encoding a 60-kilodalton glycoprotein and characterization of its 15- and 45-kilodalton zoite surface antigen products. Infect. Immun. 2000, 68, 4117–4134. [Google Scholar] [CrossRef] [Green Version]
- Dgedge, M.; Novoa, A.; Macassa, G.; Sacarlal, J.; Black, J.; Michaud, C.; Cliff, J. The burden of disease in Maputo City, Mozambique: Registered and autopsied deaths in 1994. Bull. World Health Organ. 2001, 79, 546–552. [Google Scholar] [PubMed]
- Sacarlal, J.; Nhacolo, A.Q.; Sigaúque, B.; Nhalungo, D.A.; Abacassamo, F.; Sacoor, C.N.; Aide, P.; Machevo, S.; Nhampossa, T.; Macete, E.V.; et al. A 10 year study of the cause of death in children under 15 years in Manhiça, Mozambique. BMC Public Health 2009, 9, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacoor, C.; Nhacolo, A.; Nhalungo, D.; Aponte, J.J.; Bassat, Q.; Augusto, O.; Mandomando, I.; Sacarlal, J.; Lauchande, N.; Sigaúque, B.; et al. Profile: Manhiça Health Research Centre (Manhiça HDSS). Int. J. Epidemiol. 2013, 42, 1309–1318. [Google Scholar] [CrossRef]
- Nhampossa, T.; Mandomando, I.; Acacio, S.; Quintó, L.; Vubil, D.; Ruiz, J.; Nhalungo, D.; Sacoor, C.; Nhabanga, A.; Nhacolo, A.; et al. Diarrheal disease in rural Mozambique: Burden, risk factors and etiology of diarrheal disease among children aged 0–59 months seeking care at health facilities. PLoS ONE 2015, 10, e0119824. [Google Scholar] [CrossRef]
- Casmo, V.; Lebbad, M.; Maungate, S.; Lindh, J. Occurrence of Cryptosporidium spp. and Cystoisospora belli among adult patients with diarrhoea in Maputo, Mozambique. Heliyon 2018, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Irisarri-Gutiérrez, M.J.; Mingo, M.H.; de Lucio, A.; Gil, H.; Morales, L.; Seguí, R.; Nacarapa, E.; Muñoz-Antolí, C.; Bornay-Llinares, F.J.; Esteban, J.G.; et al. Association between enteric protozoan parasites and gastrointestinal illness among HIV- and tuberculosis-infected individuals in the Chowke district, southern Mozambique. Acta Trop. 2017, 170, 197–203. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Beser, J.; Axén, C.; Lebbad, M. High applicability of a novel method for gp60-based subtyping of Cryptosporidium meleagridis. J. Clin. Microbiol. 2014, 52, 2311–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slapeta, J. Cryptosporidium species found in cattle: A proposal for a new species. Trends Parasitol. 2006, 22, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Valiathan, R.; Ashman, M.; Asthana, D. Effects of ageing on the immune system: Infants to elderly. Scand. J. Immunol. 2016, 83, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Local Burden of Disease Diarrhoea Collaborators. Mapping geographical inequalities in childhood diarrhoeal morbidity and mortality in low-income and middle-income countries, 2000–2017: Analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 1779–1801. [Google Scholar] [CrossRef]
- Muadica, A.S.; Balasegaram, S.; Beebeejaun, K.; Köster, P.C.; Bailo, B.; Hernández-de-Mingo, M.; Dashti, A.; Dacal, E.; Saugar, J.M.; Fuentes, I.; et al. Risk associations for intestinal parasites in symptomatic and asymptomatic schoolchildren in central Mozambique. Clin. Microbiol. Infect. 2020, in press. [Google Scholar]
- Delahoy, M.J.; Omore, R.; Ayers, T.L.; Schilling, K.A.; Blackstock, A.J.; Ochieng, J.B.; Moke, F.; Jaron, P.; Awuor, A.; Okonji, C.; et al. Clinical, environmental, and behavioral characteristics associated with Cryptosporidium infection among children with moderate-to-severe diarrhea in rural western Kenya, 2008-2012: The Global Enteric Multicenter Study (GEMS). PLoS Negl. Trop. Dis. 2018, 12, e0006640. [Google Scholar] [CrossRef] [Green Version]
- Acácio, S.; Mandomando, I.; Nhampossa, T.; Quintó, L.; Vubil, D.; Sacoor, C.; Kotloff, K.; Farag, T.; Nasrin, D.; Macete, E.; et al. Risk factors for death among children 0–59 months of age with moderate-to-severe diarrhea in Manhiça district, southern Mozambique. BMC Infect. Dis. 2019, 19, 322. [Google Scholar] [CrossRef]
- Tellevik, M.G.; Moyo, S.J.; Blomberg, B.; Hjøllo, T.; Maselle, S.Y.; Langeland, N.; Hanevik, K. Prevalence of Cryptosporidium parvum/hominis, Entamoeba histolytica and Giardia lamblia among young children with and without diarrhea in Dar es Salaam, Tanzania. PLoS Negl. Trop. Dis. 2015, 9, e0004125. [Google Scholar] [CrossRef] [Green Version]
- Tumwine, J.K.; Kekitiinwa, A.; Nabukeera, N.; Akiyoshi, D.E.; Rich, S.M.; Widmer, G.; Feng, X.; Tzipori, S. Cryptosporidium parvum in children with diarrhea in Mulago Hospital, Kampala, Uganda. Am. J. Trop. Med. Hyg. 2003, 68, 710–715. [Google Scholar] [CrossRef]
- Acácio, S.; Nhampossa, T.; Quintó, L.; Vubil, D.; Sacoor, C.; Kotloff, K.; Farag, T.; Dilruba, N.; Macete, E.; Levine, M.M.; et al. The role of HIV infection in the etiology and epidemiology of diarrheal disease among children aged 0–59 months in Manhiça District, Rural Mozambique. Int. J. Infect. Dis. 2018, 73, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Pavlinac, P.B.; John-Stewart, G.C.; Naulikha, J.M.; Onchiri, F.M.; Denno, D.M.; Odundo, E.A.; Singa, B.O.; Richardson, B.A.; Walson, J.L. High-risk enteric pathogens associated with HIV infection and HIV exposure in Kenyan children with acute diarrhoea. AIDS 2014, 28, 2287–2296. [Google Scholar] [CrossRef] [Green Version]
- Mengist, H.M.; Taye, B.; Tsegaye, A. Intestinal parasitosis in relation to CD4+T cells levels and anemia among HAART initiated and HAART naive pediatric HIV patients in a Model ART center in Addis Ababa, Ethiopia. PLoS ONE 2015, 10, e0117715. [Google Scholar] [CrossRef]
- Sow, S.O.; Muhsen, K.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.H.; Panchalingam, S.; Sur, D.; Zaidi, A.K.; Faruque, A.S.; et al. The burden of Cryptosporidium diarrheal disease among children <24 months of age in moderate/high mortality regions of sub-Saharan Africa and South Asia, utilizing data from the Global Enteric Multicenter Study (GEMS). PLoS Negl. Trop. Dis. 2016, 10, e0004729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernández-de-Mingo, M.; Balasegaram, S.; Carmena, D. Molecular diversity of Giardia duodenalis, Cryptosporidium spp., and Blastocystis sp. in symptomatic and asymptomatic school children in Zambézia province (Mozambique). Pathogens 2021, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Elwin, K.; Hadfield, S.J.; Robinson, G.; Crouch, N.D.; Chalmers, R.M. Cryptosporidium viatorum n. sp. (Apicomplexa: Cryptosporidiidae) among travellers returning to Great Britain from the Indian subcontinent, 2007–2011. Int. J. Parasitol. 2012, 42, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Koehler, A.V.; Wang, T.; Haydon, S.R.; Gasser, R.B. Cryptosporidium viatorum from the native Australian swamp rat Rattus lutreolus—An emerging zoonotic pathogen? Int. J. Parasitol. Parasites Wildl. 2018, 7, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhou, H.; Huang, Y.; Xu, L.; Rao, L.; Wang, S.; Wang, W.; Yi, Y.; Zhou, X.; Wu, Y.; et al. Cryptosporidium spp. in wild rats (Rattus spp.) from the Hainan Province, China: Molecular detection, species/genotype identification and implications for public health. Int. J. Parasitol. Parasites Wildl. 2019, 9, 317–321. [Google Scholar] [CrossRef]
- Mbae, C.; Mulinge, E.; Waruru, A.; Ngugi, B.; Wainaina, J.; Kariuki, S. Genetic diversity of Cryptosporidium in children in an urban informal settlement of Nairobi, Kenya. PLoS ONE 2015, 10, e0142055. [Google Scholar] [CrossRef] [PubMed]
- Akinbo, F.O.; Okaka, C.E.; Omoregie, R.; Dearen, T.; Leon, E.T.; Xiao, L. Molecular characterization of Cryptosporidium spp. in HIV-infected persons in Benin City, Edo State, Nigeria. Fooyin J. Health Sci. 2010, 2, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Lobo, M.L.; Augusto, J.; Antunes, F.; Ceita, J.; Xiao, L.H.; Codices, V.; Matos, O. Cryptosporidium spp., Giardia duodenalis, Enterocytozoon bieneusi and other intestinal parasites in young children in Lobata province, Democratic Republic of São Tomé and Príncipe. PLoS ONE 2014, 9, e97708. [Google Scholar] [CrossRef] [Green Version]
- Abu Samra, N.; Thompson, P.N.; Jori, F.; Frean, J.; Poonsamy, B.; du Plessis, D.; Mogoye, B.; Xiao, L. Genetic characterization of Cryptosporidium spp. in diarrhoeic children from four provinces in South Africa. Zoonoses Public Health 2013, 60, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Molloy, S.F.; Smith, H.V.; Kirwan, P.; Nichols, R.A.; Asaolu, S.O.; Connelly, L.; Holland, C.V. Identification of a high diversity of Cryptosporidium species genotypes and subtypes in a pediatric population in Nigeria. Am. J. Trop. Med. Hyg. 2010, 82, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Wanyiri, J.W.; Kanyi, H.; Maina, S.; Wang, D.E.; Steen, A.; Ngugi, P.; Kamau, T.; Waithera, T.; O’Connor, R.; Gachuhi, K.; et al. Cryptosporidiosis in HIV/AIDS patients in Kenya: Clinical features, epidemiology, molecular characterization and antibody responses. Am. J. Trop. Med. Hyg. 2014, 91, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.B.; Travis, D.; Lonsdorf, E.V.; Lipende, I.; Roellig, D.M.; Collins, A.; Kamenya, S.; Zhang, H.; Xiao, L.; Gillespie, T.R. Epidemiology and molecular characterization of Cryptosporidium spp. in humans, wild primates, and domesticated animals in the Greater Gombe Ecosystem, Tanzania. PLoS Negl. Trop. Dis. 2015, 9, e0003529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamu, H.; Petros, B.; Zhang, G.; Kassa, H.; Amer, S.; Ye, J.; Feng, Y.; Xiao, L. Distribution and clinical manifestations of Cryptosporidium species and subtypes in HIV/AIDS patients in Ethiopia. PLoS Negl. Trop. Dis. 2014, 8, e2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.A.; Montoya, A.; Iborra, A.; Fuentes, I. Identification of Cryptosporidium subtype isolates from HIV-seropositive patients in Equatorial Guinea. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 594–596. [Google Scholar] [CrossRef]
- Eibach, D.; Krumkamp, R.; Al-Emran, H.M.; Sarpong, N.; Hagen, R.M.; Adu-Sarkodie, Y.; Tannich, E.; May, J. Molecular characterization of Cryptosporidium spp. among children in rural Ghana. PLoS Negl. Trop. Dis. 2015, 9, e0003551. [Google Scholar] [CrossRef] [Green Version]
- Areeshi, M.; Dove, W.; Papaventsis, D.; Gatei, W.; Combe, P.; Grosjean, P.; Leatherbarrow, H.; Hart, C.A. Cryptosporidium species causing acute diarrhoea in children in Antananarivo, Madagascar. Ann. Trop. Med. Parasitol. 2008, 102, 309–315. [Google Scholar] [CrossRef]
- Akiyoshi, D.E.; Tumwine, J.K.; Bakeera-Kitaka, S.; Tzipori, S. Subtype analysis of Cryptosporidium isolates from children in Uganda. J. Parasitol. 2006, 92, 1097–1100. [Google Scholar] [CrossRef]
- Adamu, H.; Petros, B.; Hailu, A.; Petry, F. Molecular characterization of Cryptosporidium isolates from humans in Ethiopia. Acta Trop. 2010, 115, 77–83. [Google Scholar] [CrossRef]
- Rahmouni, I.; Essid, R.; Aoun, K.; Bouratbine, A. Glycoprotein 60 diversity in Cryptosporidium parvum causing human and cattle cryptosporidiosis in the rural region of Northern Tunisia. Am. J. Trop. Med. Hyg. 2014, 90, 346–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essid, R.; Menotti, J.; Hanen, C.; Aoun, K.; Bouratbine, A. Genetic diversity of Cryptosporidium isolates from human populations in an urban area of Northern Tunisia. Infect. Genet. Evol. 2018, 58, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Grace Mphephu, M.; Deogratias Ekwanzala, M.; Ndombo Benteke Momba, M. Cryptosporidium species and subtypes in river water and riverbed sediment using next-generation sequencing. Int. J. Parasitol. 2021, in press. [Google Scholar]
- Limaheluw, J.; Medema, G.; Hofstra, N. An exploration of the disease burden due to Cryptosporidium in consumed surface water for sub-Saharan Africa. Int. J. Hyg. Environ. Health 2019, 222, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Blackwelder, W.C.; Nasrin, D.; Nataro, J.P.; Farag, T.H.; van Eijk, A.; Adegbola, R.A.; Alonso, P.L.; Breiman, R.F.; Faruque, A.S.; et al. The Global Enteric Multicenter Study (GEMS) of diarrheal disease in infants and young children in developing countries: Epidemiologic and clinical methods of the case/control study. Clin. Infect. Dis. 2012, 55 (Suppl. 4), S232–S245. [Google Scholar] [CrossRef] [Green Version]
- Alonso, P.L.; Saúte, F.; Aponte, J.J.; Gómez-Olivé, F.X.; Nhacolo, A.; Thomson, R.; Macete, E.; Abacassamo, F.; Ventura, P.J.; Bosch, X.; et al. Manhiça DSS, Mozambique. In Population and Health in Developing Countries; International Development Research Centre: Otawa, ON, Canada, 2002; Volume 1, pp. 189–195. [Google Scholar]
- Levine, M.M.; Kotloff, K.L.; Nataro, J.P.; Muhsen, K. The Global Enteric Multicenter Study (GEMS): Impetus, rationale, and genesis. Clin. Infect. Dis. 2012, 55 (Suppl. 4), S215–S224. [Google Scholar] [CrossRef] [Green Version]
- Panchalingam, S.; Antonio, M.; Hossain, A.; Mandomando, I.; Ochieng, B.; Oundo, J.; Ramamurthy, T.; Tamboura, B.; Zaidi, A.K.; Petri, W.; et al. Diagnostic microbiologic methods in the GEMS-1 case/control study. Clin. Infect. Dis. 2012, 55 (Suppl. 4), S294–S302. [Google Scholar] [CrossRef] [Green Version]
- Feltus, D.C.; Giddings, C.W.; Schneck, B.L.; Monson, T.; Warshauer, D.; McEvoy, J.M. Evidence supporting zoonotic transmission of Cryptosporidium spp. in Wisconsin. J. Clin. Microbiol. 2006, 44, 4303–4308. [Google Scholar] [CrossRef] [Green Version]
- Tiangtip, R.; Jongwutiwes, S. Molecular analysis of Cryptosporidium species isolated from HIV-infected patients in Thailand. Trop. Med. Int. Health 2002, 7, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Chromas Lite 2.1—Software Informer. Available online: http://chromaslite.software.informer.com/2.1/ (accessed on 8 April 2021).
- Agarwala, R.; Barrett, T.; Beck, J.; Benson, D.A.; Bollin, C.; Bolton, E.; Bourexis, D.; Brister, J.R.; Bryant, S.H.; Canese, K.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res 2016, 44, D7–D19. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| 0–11 Months | 12–23 Months | 24–59 Months | ||||
|---|---|---|---|---|---|---|
| Variable | Cases | Non-Cases 1 | Cases | Non-Cases 1 | Cases | Non-Cases 1 |
| n = 111 (%) | n = 99 (%) | n = 72 (%) | n = 68 (%) | n = 21 (%) | n = 21 (%) | |
| MSD 2 | 87 (78.4) | 85 (85.9) | 45 (62.5) | 57 (83.8) | 12 (57.1) | 19 (90.5) |
| LSD 2 | 24 (21.6) | 14 (14.1) | 27 (37.5) | 11 (16.2) | 9 (42.9) | 2 (9.5) |
| Mean age (months) | 7.3 | 7.1 | 15.9 | 16.9 | 31.1 | 31 |
| Sex (male) | 68 (61.3) | 68 (68.7) | 48 (66.7) | 41 (61.2) | 13 (69.9) | 14 (66.7) |
| HIV+ 3 | 10/44 (24.7) | 2/18 (11.1) | 7/37 (18.9) | 2/21 (9.5) | 2/9 (22.2) | 0 (0.0) |
| Undernutrition | 14 (12.6) | 2 (2.0) | 12 (16.7) | 1 (1.5) | 1 (4.8) | 0 (0.0) |
| Co-infections | ||||||
| Rotavirus | 33 (29.7) | 13 (13.1) | 9 (12.5) | 10 (14.7) | 4 (19.1) | 1 (4.7) |
| Shigella spp. | 0 (0.0) | 0 (0.0) | 6 (8.3) | 0 (0.0) | 1 (4.8) | 0 (0.0) |
| All ETECs | 7 (6.3) | 5 (5.1) | 13 (18.1) | 7 (10.3) | 4 (19.1) | 4 (19.1) |
| G. duodenalis | 16 (14.4) | 30 (30.3) | 18 (25.0) | 38 (55.9) | 4 (19.5) | 10 (47.6) |
| E. histolytica4 | 6 (5.5)4 | 8 (8.1) | 7 (9.7) | 5 (7.4) | 2 (9.5) | 4 (19.1) |
| 0–11 Months | 12–23 Months | 24–59 Months | |||||||
|---|---|---|---|---|---|---|---|---|---|
| PCR Results | Cases | Non-Cases | p | Cases | Non-Cases | p | Cases | Non-Cases | p |
| n = 112 (%) | n = 101 (%) | n = 73 (%) | n = 67 (%) | n = 21 (%) | n = 22 (%) | ||||
| gp60 | 49 (44.1) | 24 (24.4) | 0.003 | 37 (51.4) | 30 (44.1) | 0.389 | 3 (14.3) | 3 (14.3) | 1 |
| (n = 146) | |||||||||
| ssu rRNA1 | 12/62 (19.4) | 10/75 (13.3) | 0.339 | 9/35 (25.7) | 8/38 (21.1) | 0.638 | 4/18 (22.2) | 1/18 (5.6) | 0.148 |
| (n = 44) | |||||||||
| Both | 61 (54.9) | 34 (34.3) | 0.003 | 46 (63.9) | 38 (55.9) | 0.334 | 7 (33.3) | 4 (19.1) | 0.292 |
| (n = 190) | |||||||||
| Species 2 | |||||||||
| C. hominis | 45 (47.4) | 27 (29.4) | 0.011 | 33 (55.9) | 26 (46.4) | 0.308 | 3 (22.2) | 3 (15.0) | 0.687 |
| (n = 138) | |||||||||
| C. parvum | 14 (21.9) | 6 (8.5) | 0.028 | 12 (31.6) | 9 (23.1) | 0.402 | 2 (12.5) | 0 (0.0) | 0.229 |
| (n = 43) | |||||||||
| C. meleagridis | 1 (1.9) | 1 (1.5) | 1 | 1 (3.7) | 3 (9.1) | 0.620 | 1 (6.7) | 1 (5.6) | 1 |
| (n = 8) | |||||||||
| Unknown 3 | 1 (1.9) | 0 (0.0) | 0.440 | 0 (0.0) | 0 (0.0) | NA | 0 (0.0) | 0 (0.0) | NA |
| (n = 1) | |||||||||
| Locus | Species | Isolates | Family | Subtype | Reference | Stretch | Single Nucleotide Polymorphisms | GenBank ID |
|---|---|---|---|---|---|---|---|---|
| gp60 | C. hominis | 37 | Ia | IaA23R3 | KX579755 | 3–805 | None | MW480826 |
| 1 | IaA24R3 | KX579755 | 1–793 | 84_86InsTCA | MW480827 | |||
| 3 | IaA25R3 | JF927194 | 18–838 | None | MW480828 | |||
| 7 | Ib | IbA10G2 | AY262031 | 22–857 | None | MW480829 | ||
| 15 | IbA13G2 | MT053132 | 13–896 | G85A | MW480830 | |||
| 2 | IbA9G3 | DQ665688 | 14–825 | None | MW480831 | |||
| 1 | Id | IdA20 | JX088404 | 48–904 | None | MW480832 | ||
| 1 | IdA21 | MN904672 | 47–910 | None | MW480833 | |||
| 39 | Ie | IeA11G3T3 | AY738184 | 19–923 | None | MW480834 | ||
| 1 | IeA11G3T3 | AY738184 | 48–923 | T284Y, A662R | MW480835 | |||
| 10 | If | IfA12G1 | EU161655 | 1–870 | None | MW480836 | ||
| C. parvum | 1 | IIb | IIbA11 | AY166805 | 1–782 | 51_59DelTCATCATCA | MW480837 | |
| 11 | IIc | IIcA5G3 | GU214365 | 31–851 | None | MW480838 | ||
| 7 | IIcA5G3 | GU214365 | 29–854 | 38 SNPs 2 | MW480839 | |||
| 5 | IIcA5G3 | GU214365 | 29–853 | 40 SNPs 2 | MW480840 | |||
| 1 | IIcA5G3 | GU214365 | 50–853 | C110T | MW480841 | |||
| 1 | IIcA5G3 | GU214365 | 50–853 | 40 SNPs 2 | MW480842 | |||
| 1 | IIe | IIeA11G1 | MN904721 | 1–813 | None | MW480843 | ||
| 1 | IIeA13G1 | KU852716 | 7–795 | None | MW480844 | |||
| 1 | IIi | IIiA6-like | AY873782 | 26–932 | 85 SNPs 2 | MW480845 | ||
| C. meleagridis1 | 4 | IIIb | IIIbA23G1R1 | MK331716 | 1–714 | None | MW480846 | |
| ssu rRNA | C. hominis | 17 | – | – | AF108865 | 529–954 | None | MW487256 |
| 1 | – | – | AF108865 | 587–965 | A892R | MW487257 | ||
| 2 | – | – | AF108865 | 591–969 | T795Y, A892R | MW487258 | ||
| 1 | – | – | AF108865 | 640–956 | 697delT, T795Y, A892R | MW487259 | ||
| C. parvum | 1 | – | – | AF112571 | 565–956 | A646G, T649G, 686_689DelTAAT, A691T | MW487260 | |
| 3 | – | – | AF112571 | 526–1039 | A646G, T649G, 686_689DelTAAT, T693A | MW487261 | ||
| 1 | – | – | AF112571 | 524–1039 | A646G, 647_649DelATT, T663C, 686_689DelTAAT, C795T | MW487262 | ||
| 5 | – | – | AF112571 | 539–1031 | A646G, T649G, 686_689DelTAAT, T693A, C795T | MW487263 | ||
| 1 | – | – | AF112571 | 526–965 | A646G, T649G, 686_689DelTAAT, T693A, C795Y | MW487264 | ||
| 3 | – | – | AF112571 | 539–954 | A646G, T649G, 686_689DelTAAT, A691T, C795Y, A892R | MW487265 | ||
| Unknown | 1 | – | – | – | – | – | – | |
| C. meleagridis | 8 | – | – | AF112574 | 524–1034 | None | MW487266 |
| C. hominis | C. parvum | C. meleagridis | ||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Total | Ia | Ib | Id | Ie | If | IIc | IIIb |
| n = 90 (%) | n = 25 (%) | n = 11 (%) | n = 2 (%) | n = 28 (%) | n = 6 (%) | n = 17 (%) | n = 1 (%) | |
| Diarrhoea | ||||||||
| MSD | 60 (66.7) | 11 (44.0) | 7 (63.6) | 1 (50.0) | 26 (92.9) | 5 (83.3) | 9 (52.9) | 1 (100) |
| LSD | 30 (33.3) | 14 (56.0) | 4 (36.4) | 1 (50.0) | 2 (7.1) | 1 (16.7) | 8 (47.1) | 0 (0) |
| Age (months) | ||||||||
| 0–11 | 49 (54.4) | 16 (64.0) | 8 (72.7) | 1 (50.0) | 14 (50) | 2 (33.3) | 8 (47.1) | 0 (0) |
| 12–23 | 37 (41.1) | 7 (28.0) | 3 (27.3) | 1 (50.0) | 13 (46.4) | 4 (66.7) | 9 (52.9) | 0 (0) |
| 24–59 | 4 (4.4) | 2 (8.0) | 0 (0.0) | 0 (0.0) | 1 (3.6) | 0 (0.0) | 0 (0.0) | 1 (100) |
| Co-infections | ||||||||
| HIV+ 1 | 9/42 (21.4) | 5/17 (29.4) | 1/4 (25) | 0/1 (0.0) | 1/5 (20) | 0/1 (0.0) | 2/14 (14.3) | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messa, A., Jr.; Köster, P.C.; Garrine, M.; Nhampossa, T.; Massora, S.; Cossa, A.; Bassat, Q.; Kotloff, K.; Levine, M.M.; Alonso, P.L.; et al. Molecular Characterisation of Cryptosporidium spp. in Mozambican Children Younger than 5 Years Enrolled in a Matched Case-Control Study on the Aetiology of Diarrhoeal Disease. Pathogens 2021, 10, 452. https://doi.org/10.3390/pathogens10040452
Messa A Jr., Köster PC, Garrine M, Nhampossa T, Massora S, Cossa A, Bassat Q, Kotloff K, Levine MM, Alonso PL, et al. Molecular Characterisation of Cryptosporidium spp. in Mozambican Children Younger than 5 Years Enrolled in a Matched Case-Control Study on the Aetiology of Diarrhoeal Disease. Pathogens. 2021; 10(4):452. https://doi.org/10.3390/pathogens10040452
Chicago/Turabian StyleMessa, Augusto, Jr., Pamela C. Köster, Marcelino Garrine, Tacilta Nhampossa, Sérgio Massora, Anélsio Cossa, Quique Bassat, Karen Kotloff, Myron M. Levine, Pedro L. Alonso, and et al. 2021. "Molecular Characterisation of Cryptosporidium spp. in Mozambican Children Younger than 5 Years Enrolled in a Matched Case-Control Study on the Aetiology of Diarrhoeal Disease" Pathogens 10, no. 4: 452. https://doi.org/10.3390/pathogens10040452
APA StyleMessa, A., Jr., Köster, P. C., Garrine, M., Nhampossa, T., Massora, S., Cossa, A., Bassat, Q., Kotloff, K., Levine, M. M., Alonso, P. L., Carmena, D., & Mandomando, I. (2021). Molecular Characterisation of Cryptosporidium spp. in Mozambican Children Younger than 5 Years Enrolled in a Matched Case-Control Study on the Aetiology of Diarrhoeal Disease. Pathogens, 10(4), 452. https://doi.org/10.3390/pathogens10040452