
To Treat or Not to Treat Bees? Handy VarLoad: A Predictive Model for Varroa destructor Load


 and
and
Abstract
:1. Introduction
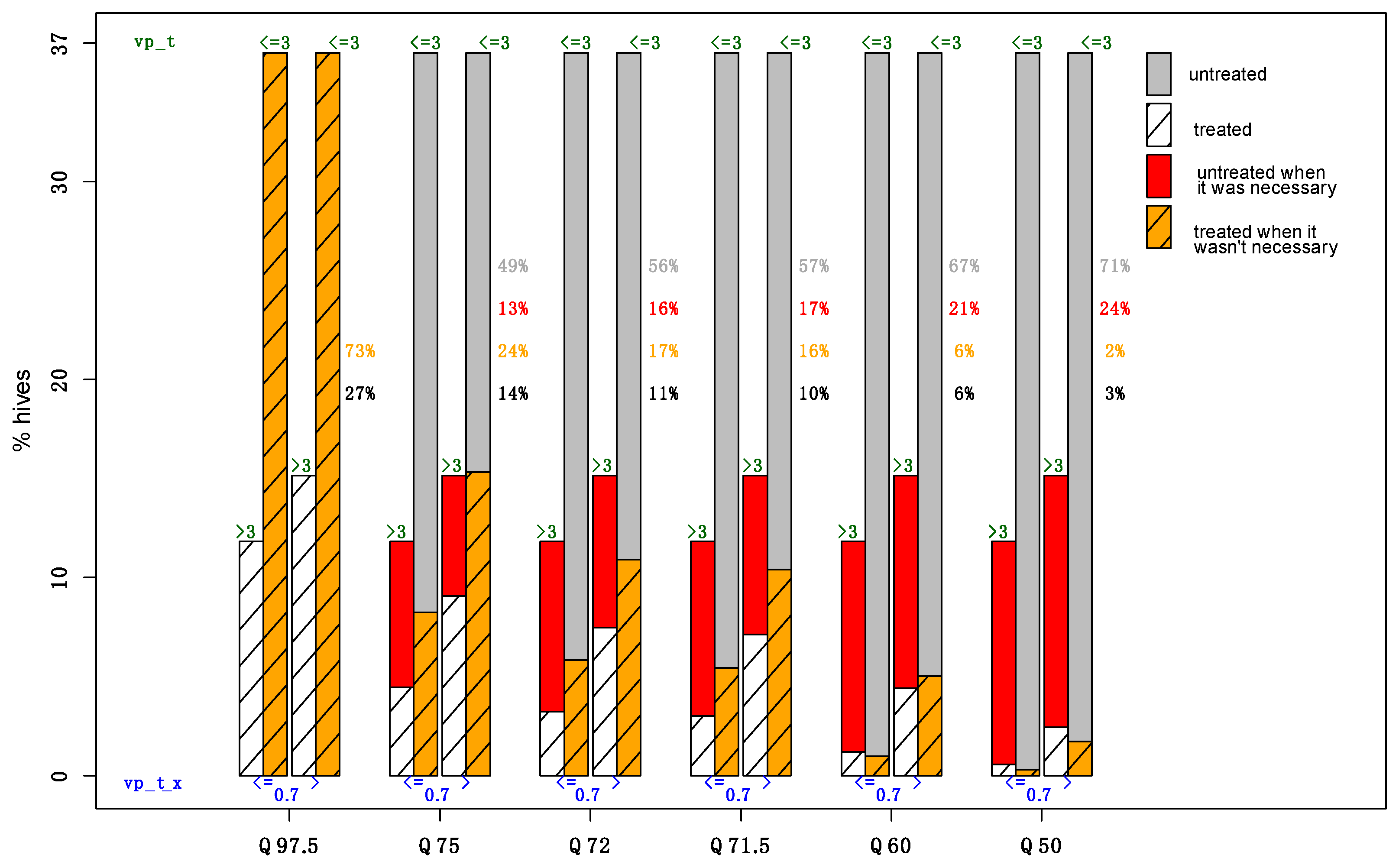
2. Results
2.1. Variable Selection (for Variable Definitions, See Materials and Methods, Statistical Analysis)
Vpt = 0 with ν probability
σ = exp (β0 + β1Vbt−x + β2Dt + Ap)
ν = logit−1 (γ0 + γ1Vbt−x + γ2Cpt−x + γ3Dt + Ap)
σ = exp (β0 + Ap)
ν = logit−1 (γ0 + γ1Vbt−x + γ2Vpt−x)
2.2. Goodness of Fit and Prediction Evaluation
2.2.1. Parameter Uncertainty
2.2.2. Prediction Quality
3. Discussion
3.1. Selected Variables
3.2. Beekeepers’ Interest
3.3. Limits and Prospects of the Model
4. Materials and Methods
4.1. Data Sampling
4.2. Statistical Methods
4.2.1. Distribution Adjustment on “dataset1”
4.2.2. Goodness of Fit and Prediction Including “dataset2”
- The actual coverage of 95%, 70%, and 50% confidence intervals of Vpt (denoted by CI95%, CI70%, and CI50%), providing the proportion of times that the true value of Vpt is contained within the CI;
- The use of different predicted quantiles of Vpt (namely, Q97,5%, Q85%, Q75%, and Q50%) to evaluate the risk that the actual Vpt exceeds the problematic threshold of 3 Varroa mites for 100 bees.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, 96–119. [Google Scholar] [CrossRef]
- Denmark, H.A.; Cromroy, H.L.; Cutts, L. Varroa Mite. Varroa Jacobsoni Oudemans: (Acari: Varroidae); Fla. Department Agric. & Consumer Serv., Division of Plant Industry: Gainesville, FL, USA, 1991.
- Baxter, J.; Eischen, F.; Pettis, J.; Wilson, W.T.; Shimanuki, H. Detection of fluvalinate-resistant Varroa mites in US honey bees. Am. Bee J. 1998, 138, 291. [Google Scholar]
- Elzen, P.J.; Westervelt, D. Detection of coumaphos resistance in Varroa destructor in Florida. Am. Bee J. 2002, 142, 291–292. [Google Scholar]
- Pettis, J.S. A scientific note on Varroa destructor resistance to coumaphos in the United States. Apidologie 2004, 35, 91–92. [Google Scholar] [CrossRef] [Green Version]
- De Jong, D.; De Jong, P.H.; Goncalves, L.S. Weight loss and other damage to developing worker honeybees from infestation with V. jacobsoni. J. Apic. Res. 1982, 21, 165–216. [Google Scholar] [CrossRef]
- Schneider, P.; Drescher, W. Einfluss der Parasitierung durch die Milbe Varroa Jacobsoni aus das Schlupfgewicht, die Gewichtsentwicklung, die Entwicklung der Hypopharynxdrusen und die Lebensdauer von Apis mellifera. Apidologie 1987, 18, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef] [Green Version]
- Emsen, B.; Guzman-Novoa, E.; Kelly, P.G. Honey production of honey bee (Hymenoptera: Apidae) colonies with hight and low Varroa destructor (Acari: Varroidae) infestation rates in eastern Canada. Can. Entomol. 2014, 146, 236–240. [Google Scholar] [CrossRef]
- Kretzschmar, A.; Maisonnasse, A.; Dussaubat, C.; Cousin, M.; Vidau, C. Performances des colonies vues par les observatoires de ruchers. Innov. Agron. 2016, 53, 81–93. [Google Scholar]
- Arechavaleta-Velasco, M.E.; Guzman-Novoa, E. Relative effect of four characteristics that restrain the population growth of the mite Varroa destructor in honey bee (Apis mellifera L.) colonies. Apidologie 2001, 32, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.W.; Harbo, J.R.; Villa, J.D.; Danka, R.G. Variable population growth of Varroa destructor (Mesostigmata: Varroidae) in colonies of honey bees hymenoptera: Apidae) during a 10-year period. Environ. Entomol. 2003, 32, 1305–1312. [Google Scholar] [CrossRef]
- Lodesani, M.; Crailsheim, C.; Moritz, R.F.A. Effect of some characters on the population growth of mite Varroa jacobsoni in Apis mellifera L. colonies and results of a bi-directional selection. J. Appl. Entomol. 2002, 126, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, D.; Smith, G.C. A model of the mite parasite, Varroa destructor, on honeybees (Apis mellifera) to investigate parameters important to mite population growth. Ecol. Model. 2002, 148, 263–275. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Curry, R. A mathematical model of Varroa mite (Varroa destructor Anderson and Trueman) and honeybee (Apis mellifera L.) population dynamics. Int. J. Acarol. 2004, 30, 259–274. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Roth, S.A.; Loper, G.L.; Erickson, E.H., Jr. BEEPOP: A honeybee population dynamics simulation model. Ecol. Model. 1989, 45, 133–150. [Google Scholar] [CrossRef]
- Becher, M.A.; Grimm, V.; Thorbek, P.; Horn, J.; Kennedy, P.J.; Osborne, J.L. BEEHAVE: A systems model of honeybee colony dynamics and foraging to explore multifactorial causes of colony failure. J. Appl. Ecol. 2014, 51, 470–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jay, S.C. The development of honeybees in their cells. J. Apic. Res. 1963, 2, 117–134. [Google Scholar] [CrossRef]
- De Jong, D.; De Jong, P.H. Longevity of Africanized honey bees (Hymenoptera: Apidae) infested by Varroa jacobsoni (Parasitiformes: Varroidae). J. Econ. Entomol. 1983, 76, 766–768. [Google Scholar] [CrossRef]
- Kovac, H.; Crailsheim, K. Lifespan of Apis mellifera carnica Pollm. infested by Varroa jacobsoni Oud. In relation to season and extent of infestation. J. Apic. Res. 1988, 27, 230–238. [Google Scholar] [CrossRef]
- Fries, I.; Camazine, S.; Sneyd, J. Population dynamics of Varroa jacobsoni: A model and a review. Bee World 1994, 75, 4–28. [Google Scholar] [CrossRef]
- Donzé, G.; Herrmann, M.; Bachofen, B.; Guerin, P.M. Effect of mating frequency and brood cell infestation rate on the reproductive success of the honeybee parasite Varroa jacobsoni. Ecol. Entomol. 1996, 21, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Kemp, D. Average number of reproductive cycles performed by Varroa jacobsoni in honey bee (Apis mellifera) colonies. J. Apic. Res. 1997, 36, 113–123. [Google Scholar] [CrossRef]
- Calis, J.N.M.; Fries, I.; Ryrie, S.C. Population modeling of Varroa jacobsoni Oud. Apidologie 1999, 30, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J. A population model for the ectoparasitic mite Varroa jacobsoni in honey bee (Apis mellifera) colonies. Ecol. Model. 1998, 109, 267–281. [Google Scholar] [CrossRef]
- Martin, S.J. The role of Varroa and viral pathogens in the collapse of honey bee colonies: A modeling approach. J. Appl. Entomol. 2001, 38, 1082–1093. [Google Scholar]
- Kurze, C.; Routtu, J.; Moritz, R.F. Parasite resistance and tolerance in honeybees at the individual and social level. Zoology 2016, 119, 290–297. [Google Scholar] [CrossRef]
- Camazine, S. Differential reproduction of the mite Varroa jacobsoni (Mesostigmata: Varroidae), on Africanized and European honey bees (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 1986, 79, 801–803. [Google Scholar] [CrossRef]
- Bogdavov, S.; Charrière, J.-D.; Imdorf, A.; Kilchenmann, V.; Fluri, P. Determination of residues in honey after treatments with formic and oxalic acid under field conditions. Apidologie 2002, 33, 399–409. [Google Scholar] [CrossRef]
- Maisonnasse, A.; Frontero, L.; Kretzschmar, A. Mesurer le taux de VP/100ab (Varroa phorétique pour 100 abeilles) dans les ruchers pour optimiser la gestion et la production. In Proceedings of the 5ème Journée de la Recherche Apicole, Paris, France, 8–9 February 2017. [Google Scholar]
- Sumpter, D.J.T.; Broomhead, D.S. Relating individual behaviour to population dynamics. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Eguaras, M.; Marcangeli, J.; Oppedisano, M.; Fernández, N. Seasonal changes in Varroa jacobsoni reproduction in temperate climates of Argentina. Bee Sci. 1994, 3, 120–123. [Google Scholar]
- Garcia-Fernandez, P.; Rodriguez, R.B.; Orantesbermejo, F.J. Influence of climate on the evolution of the population-dynamics of the Varroa mite on honeybees in the south of Spain. Apidologie 1995, 26, 371–380. [Google Scholar]
- Kraus, B.; Velthuis, H.H.W. High humidity in the honey bee (Apis mellifera L.) brood nest limits reproduction of the parasitic mite Varroa jacobsoni Oud. Naturwissenschaften 1997, 84, 217–218. [Google Scholar] [CrossRef] [Green Version]
- Currie, R.W.; Tahmasbi, G.H. The ability of high-and low-grooming lines of honey bees to remove the parasitic mite Varroa destructor is affected by environmental conditions. Can. J. Zool. 2008, 86, 1059–1067. [Google Scholar] [CrossRef]
- Mondet, F.; Maisonnasse, A.; Kretzschmar, A.; Alaux, C.; Vallon, J.; Basso, B.; Dangleant, A.; Le Conte, Y. Varroa: Son impact, les méthodes d’évaluation de l’infestation et les moyens de lutte. Innov. Agron. 2016, 53, 63–80. [Google Scholar]
- Hernandez, J.; Maisonnasse, A.; Cousin, M.; Beri, C.; Le Quintrec, C.; Bouetard, A.; Castex, D.; Decante, D.; Servel, E.; Buchwalder, G.; et al. ColEval: Honeybee COLony Structure EVALuation for Field Surveys. Insects 2020, 11, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. BEEBOOK Volume II. Standard Methods for Varroa Research 4.2.3.1.2.3; University of Bern: Bern, Switzerland, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 25 May 2021).
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape, (with discussion). Appl. Stat. 2005, 54, 507–554. [Google Scholar] [CrossRef] [Green Version]
- Rigby, R.A.; Stasinopoulos, D.M. The GAMLSS project: A flexible approach to statistical modelling. In New Trends in Statistical Modelling, Proceedings of the 16th International Workshop on Statistical Modelling, Odense, Denmark, 2–6 July 2001; Klein, B., Korsholm, L., Eds.; Odense University Press: Odense, Denmark, 2001; pp. 249–256. [Google Scholar]
- Hurvich, C.M.; Tsai, C.-L. Regression and Time Series Model Selection in Small Samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Aho, K.; Derryberry, D.; Peterson, T. Model selection for ecologists: The worldviews of AIC and BIC. Ecology 2014, 95, 631–636. [Google Scholar] [CrossRef]


{kind=link}
| Adjustment for x = 1 | Adjustment for x = 3 | |
|---|---|---|
| Model | AICc | AICc |
| phoretic Varroa | −2477.5 | −268.3 |
| capped brood cells | −2320.6 | −261.8 |
| varbrood (**) | −2540.1 | −296.9 |
| date | −2413.0 | −260.9 |
| phoretic Varroa + capped brood cells | −2488.0 | −271.1 |
| phoretic Varroa + date | −2561.7 | −266.3 |
| phoretic Varroa + varbrood | −2538.2 | −295.7 |
| capped brood cells + date | −2412.1 | −261.4 |
| capped brood cells + varbrood | −2580.1 | −297.7 |
| date + varbrood | −2618.2 | −294.7 |
| phoretic Varroa + capped brood cells + date | −2564.9 | −269.1 |
| phoretic Varroa + capped brood cells + varbrood | −2582.4 | −296.0 |
| phoretic Varroa + date + varbrood | −2616.2 | −293.8 |
| capped brood cells + date + varbrood (*) | −2645.5 | −295.5 |
| phoretic Varroa + capped brood cells + varbrood + date | −2647.8 | −293.9 |
| (*) and (**) + apiary random effect | −2651.1 | −316.9 |
| Model | Parameter | Covariate | Estimated Coefficient | Lower 95% CI | Upper 95% CI |
|---|---|---|---|---|---|
| A | Mu | Intercept | −5.830 | −6.021 | −5.640 |
| varbrood | 0.025 | 0.021 | 0.028 | ||
| capped brood cells | 0.002 | 0.001 | 0.003 | ||
| date | 0.014 | 0.012 | 0.015 | ||
| Sigma | Intercept | 6.579 | 6.233 | 6.925 | |
| varbrood | −0.023 | −0.030 | −0.016 | ||
| date | −0.018 | −0.021 | −0.015 | ||
| Nu | Intercept | 2.073 | 1.475 | 2.672 | |
| varbrood | −0.063 | −0.087 | −0.039 | ||
| capped brood cells | −0.003 | −0.006 | −0.001 | ||
| date | −0.032 | −0.039 | −0.025 | ||
| B | Mu | Intercept | −3.982 | −4.175 | −3.790 |
| varbrood | 0.023 | 0.019 | 0.027 | ||
| Sigma | Intercept | 4.460 | 4.140 | 4.779 | |
| Nu | Intercept | −0.701 | −1.468 | 0.065 | |
| varbrood | −0.077 | −0.167 | 0.012 | ||
| phoretic Varroa | −3.786 | −10.747 | 3.176 |
| Cross-Validation | |||||||||
| Model A | Model B | Observed Vpt | Model A | Model B | |||||
| Observed Colony Numbers | CI95% | CI70% | CI50% | CI95% | CI70% | CI50% | |||
| 4999 | 2328 | all | 97.6 | 83.6 | 67.7 | 97.3 | 83.1 | 67.8 | |
| 4027 | 1700 | ≤3 | 99.6 | 91.5 | 76.3 | 99.7 | 97.9 | 87.9 | |
| 724 | 526 | >3 and ≤10 | 92.7 | 53.7 | 34.5 | 99.8 | 51.1 | 16 | |
| 248 | 102 | >10 | 80.6 | 42.3 | 24.2 | 45.1 | 2 | 0 | |
| Training Validation | |||||||||
| Model A | Model B | Observed Vpt | Model A | Model B | |||||
| Observed Colony Numbers | CI95% | CI70% | CI50% | CI95% | CI70% | CI50% | |||
| 1438 | 749 | all | 92.6 | 75.3 | 61.8 | 57.8 | 39 | 26 | |
| 1140 | 546 | ≤3 | 95.9 | 82.6 | 69 | 61.2 | 44.1 | 29.9 | |
| 229 | 137 | >3 and ≤10 | 82.1 | 49.8 | 37.6 | 60.6 | 29.9 | 17.5 | |
| 69 | 66 | >10 | 72.5 | 39.1 | 21.7 | 24.2 | 15.2 | 12.1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dechatre, H.; Michel, L.; Soubeyrand, S.; Maisonnasse, A.; Moreau, P.; Poquet, Y.; Pioz, M.; Vidau, C.; Basso, B.; Mondet, F.; et al. To Treat or Not to Treat Bees? Handy VarLoad: A Predictive Model for Varroa destructor Load. Pathogens 2021, 10, 678. https://doi.org/10.3390/pathogens10060678
Dechatre H, Michel L, Soubeyrand S, Maisonnasse A, Moreau P, Poquet Y, Pioz M, Vidau C, Basso B, Mondet F, et al. To Treat or Not to Treat Bees? Handy VarLoad: A Predictive Model for Varroa destructor Load. Pathogens. 2021; 10(6):678. https://doi.org/10.3390/pathogens10060678
Chicago/Turabian StyleDechatre, Hélène, Lucie Michel, Samuel Soubeyrand, Alban Maisonnasse, Pierre Moreau, Yannick Poquet, Maryline Pioz, Cyril Vidau, Benjamin Basso, Fanny Mondet, and et al. 2021. "To Treat or Not to Treat Bees? Handy VarLoad: A Predictive Model for Varroa destructor Load" Pathogens 10, no. 6: 678. https://doi.org/10.3390/pathogens10060678
APA StyleDechatre, H., Michel, L., Soubeyrand, S., Maisonnasse, A., Moreau, P., Poquet, Y., Pioz, M., Vidau, C., Basso, B., Mondet, F., & Kretzschmar, A. (2021). To Treat or Not to Treat Bees? Handy VarLoad: A Predictive Model for Varroa destructor Load. Pathogens, 10(6), 678. https://doi.org/10.3390/pathogens10060678