Maize (Zea mays L.): A New Host for Ligustrum witches’ Broom Phytoplasma



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Detection of Phytoplasma in Field-Collected Samples
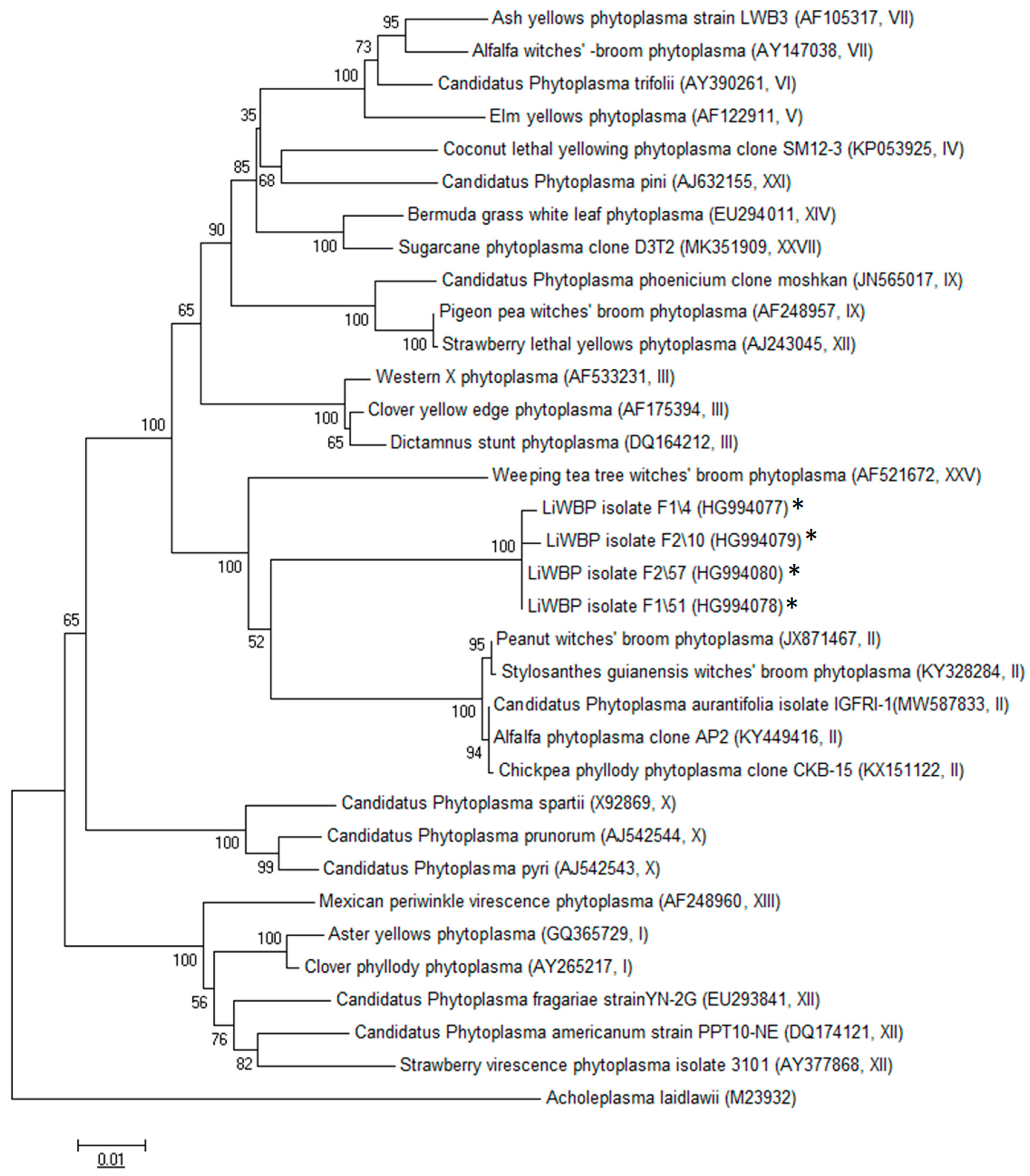
2.2. Virtual RFLP and Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. DNA Extraction
4.3. Polymerase Chain Reaction (PCR)
4.4. Sequencing, Virtual RFLP and Phylogenetic Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turkish Statistical Institute (TUIK). Crop Production Statistics. 2019. Available online: https://data.tuik.gov.tr/ (accessed on 10 March 2021).
- Bayramoglu, Z.; Bozdemir, M. Türkiye’de üretilen mısırın ekonomik gelişim seyri. Türk Tarım-Gıda Bilim Ve Teknoloji Dergisi 2018, 6, 1092–1100. [Google Scholar] [CrossRef]
- Turkish Statistical Institute (TUIK). Crop Production Statistics. 2021. Available online: https://data.tuik.gov.tr/ (accessed on 10 March 2021).
- Ertunç, F. A new threat for Turkish horticulture: Phytoplasma diseases and their vector. Vet. J. Ank. Univ. 2013, 60, 221–224. [Google Scholar]
- Canik, D.; Ertunc, F.; Paltrinieri, S.; Contaldo, N.; Bertaccini, A. Identification of different phytoplasmas infecting grapevine in Turkey. Bull. Insectol. 2019, 64, 225–226. [Google Scholar]
- Çağlar, B.K.; Satar, S.; Bertaccini, A.; Elbeaino, T. Detection and seed transmission of Bermuda grass phytoplasma in maize in Turkey. J. Phytopathol. 2019, 167, 248–255. [Google Scholar] [CrossRef]
- Çağlar, B.K.; Elbeaino, T. Detection and characterization of Ligustrum witches’ broom phytoplasma in a new host (Robinia Pseudoacacia, L.) in Adana Province of Turkey. In Proceedings of the 5th Plant Protection Congress, Antalya, Turkey, 3–5 February 2014; p. 289. [Google Scholar]
- Çağlar, B.K.; Satar, S.; Elbeaino, T. Detection and molecular characterization of Bermudagrass (Cynodon dactylon) white leaf phytoplasma from Turkey. Int. J. Agric. Biol. 2013, 15, 90–94. [Google Scholar]
- Çağlar, B.K.; Elbeaino, T.; Kusek, M.; Pehlivan, D.; Fidan, H.; Portakaldali, M. Stolbur phytoplasma infections in potato and tomato plants from different locations in Turkey. J. Turk. Phytopathol. 2013, 39, 1–8. [Google Scholar]
- Ozdemir, Z.; Cagirgan, M.I. Identification and characterization of a phytoplasma disease of jute (Corchorus olitorius L.) from south-western Turkey. Crop Prot. 2015, 74, 1–8. [Google Scholar] [CrossRef]
- Catal, M.; Ikten, C.; Yol, E.; Ustun, R.; Uzun, B. First report of a 16SrIX group (pigeon pea witches’ broom) phytoplasma associated with sesame phyllody in Turkey. Plant Dis. 2013, 97, 835. [Google Scholar] [CrossRef] [PubMed]
- Çaglayan, K.; Gazel, M.; Ulubas, S.Ç.; Bozkurt, I.A.; Elçi, E. Phytoplasma diseases of stone fruit trees in Turkey and their containment. Phytopathogenic Mollicutes 2011, 12, 95–97. [Google Scholar] [CrossRef]
- Sertkaya, G.; Martini, M.; Musetti, R.; Osler, R. Detection and molecular characterization of phytoplasmas infecting sesame and solanaceous crops in Turkey. Bull. Insectol. 2007, 60, 141–142. [Google Scholar]
- Çağlar, B.K.; Elbeaino, T. A novel phytoplasma associated with witches’-broom disease of Ligustrum ovalifolium in Turkey. Eur. J. Plant Pathol. 2013, 137, 113–117. [Google Scholar] [CrossRef]
- Wei, W.; Lee, I.M.; Davis, R.E.; Suo, X.; Zhao, Y. Automated RFLP pattern comparison and similarity coefficient calculation for rapid delineation of new and distinct phytoplasma 16Sr subgroup lineages. Int. J. Syst. Evol. Microbiol. 2008, 58, 2368–2377. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- IRPCM Phytoplasma/Spiroplasma Working Team-Phytoplasma Taxonomy Group. “Candidatus Phytoplasma”, a taxon for the wall-less, non-helical prokaryotes that colonize plant phloem and insects. Int. J. Syst. Evol. Microbiol. 2004, 54, 1243–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrens, U.; Seemüller, E. Detection of DNA of plant pathogenic mycoplasma-like organisms by a polymerase chain reaction that amplifies a sequence of the 16S rRNA gene. Phytopathology 1992, 82, 828–832. [Google Scholar] [CrossRef]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA genes from culturable and nonculturable mollicutes. J. Microbiol. Methods 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Schneider, B.; Seemuller, E.; Smart, C.D.; Kirkpatrick, B.C. Phylogenetic classification of plant pathogenic mycoplasma-like organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmolgy; Razin, S., Tully, J.G., Eds.; Academic Press: New York, NY, USA, 1995; pp. 369–380. [Google Scholar]
- Lee, I.M.; Hammond, R.W.; Davis, R.E.; Gundersen, D.E. Universal amplification and analysis of pathogen 16S rDNA for classification and identification of mycoplasmalike organism. Phytopathology 1993, 83, 834–842. [Google Scholar] [CrossRef]
- Gundersen, D.E.; Lee, I.M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol. Medit. 1996, 35, 144–151. [Google Scholar]
- Zhao, Y.; Wei, W.; Lee, I.M.; Shao, J.; Suo, X.; Davis, R.E. The iPhyClassifier, an interactive online tool for phytoplasma classification and taxonomic assignment. In Phytoplasma; Dickinson, M., Hodgetts, J., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 329–338. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çağlar, B.K.; Pehlivan, S.; Atakan, E.; Elbeaino, T. Maize (Zea mays L.): A New Host for Ligustrum witches’ Broom Phytoplasma. Pathogens 2021, 10, 723. https://doi.org/10.3390/pathogens10060723
Çağlar BK, Pehlivan S, Atakan E, Elbeaino T. Maize (Zea mays L.): A New Host for Ligustrum witches’ Broom Phytoplasma. Pathogens. 2021; 10(6):723. https://doi.org/10.3390/pathogens10060723
Chicago/Turabian StyleÇağlar, Behçet Kemal, Serkan Pehlivan, Ekrem Atakan, and Toufic Elbeaino. 2021. "Maize (Zea mays L.): A New Host for Ligustrum witches’ Broom Phytoplasma" Pathogens 10, no. 6: 723. https://doi.org/10.3390/pathogens10060723
APA StyleÇağlar, B. K., Pehlivan, S., Atakan, E., & Elbeaino, T. (2021). Maize (Zea mays L.): A New Host for Ligustrum witches’ Broom Phytoplasma. Pathogens, 10(6), 723. https://doi.org/10.3390/pathogens10060723