
Co-Infection of Escherichia coli, Enterococcus faecalis and Chlamydia psittaci Contributes to Salpingitis of Laying Layers and Breeder Ducks


Abstract
:1. Introduction
2. Results
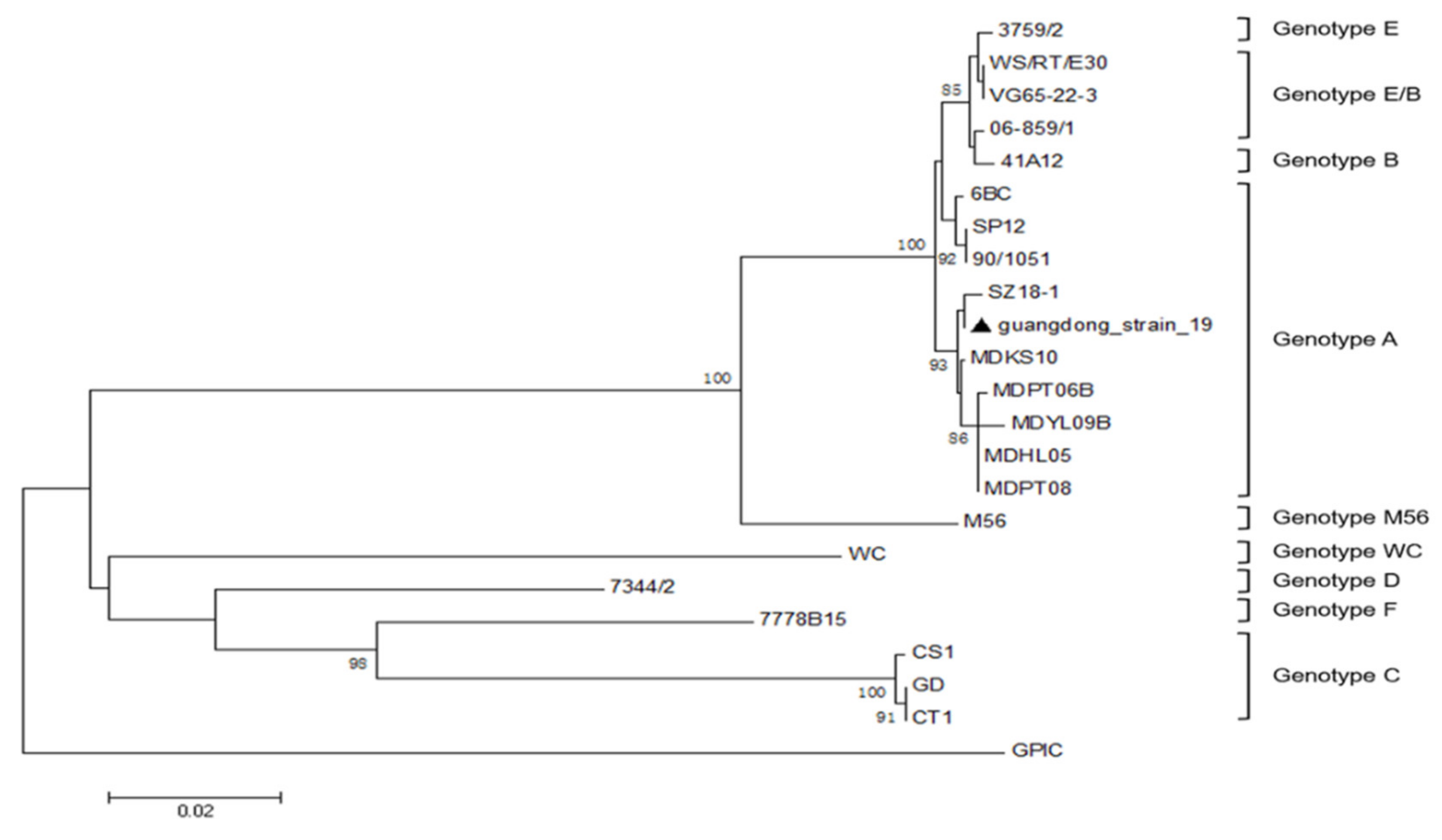
2.1. Seroprevalence of C. psittaci in the Affected Flocks
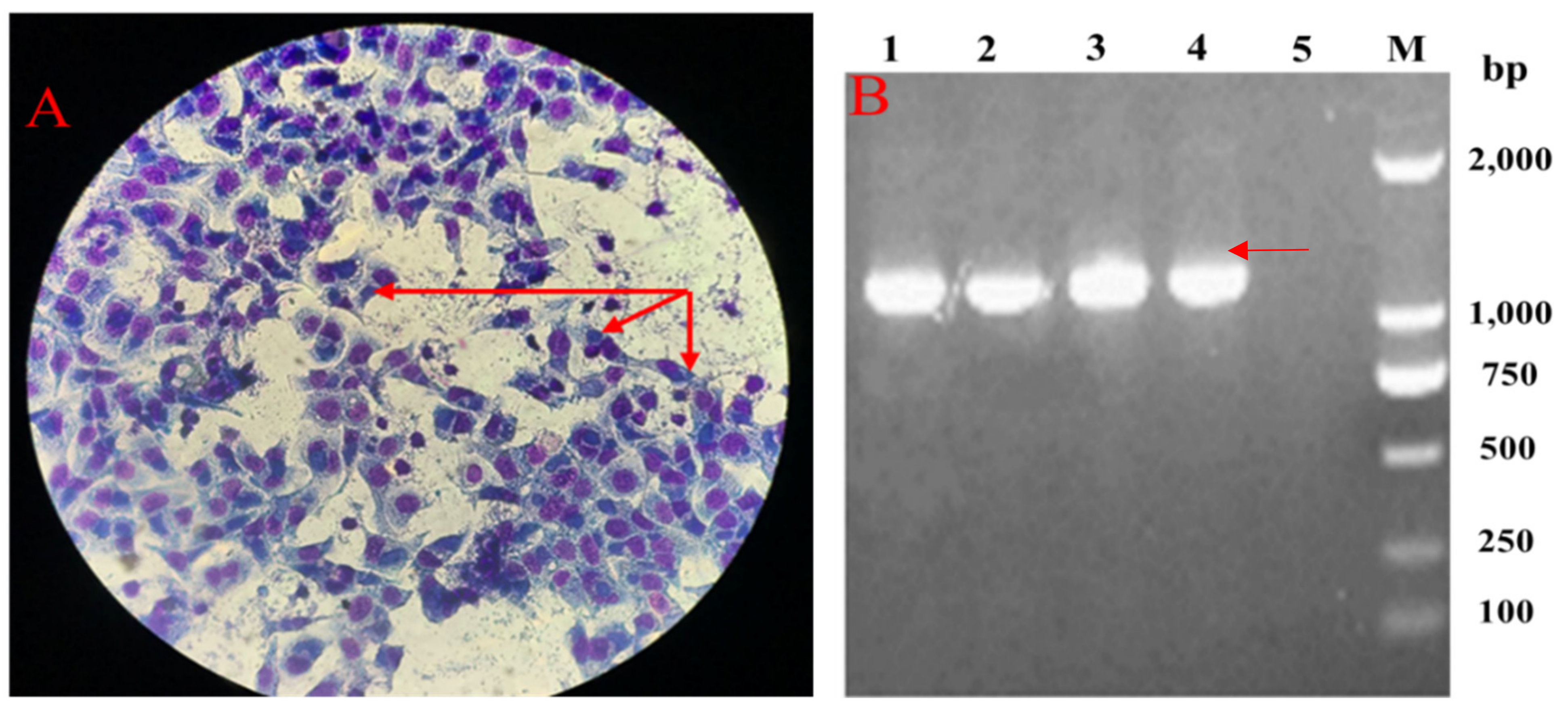
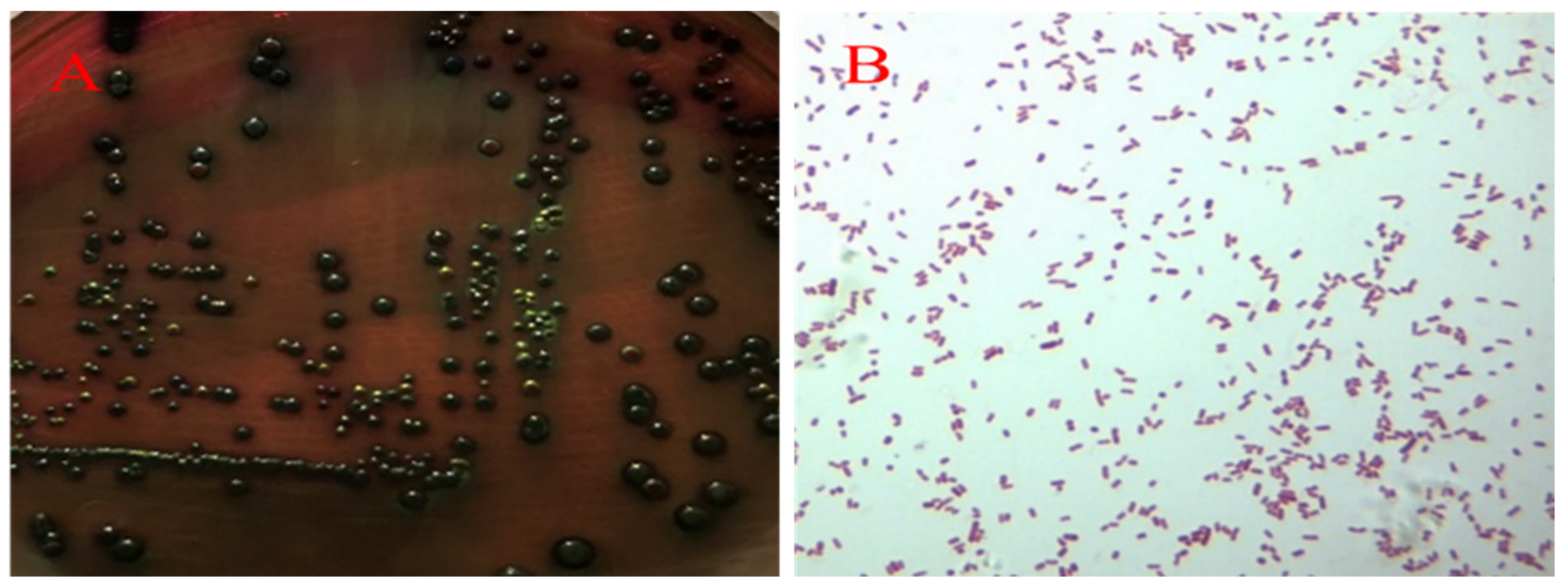
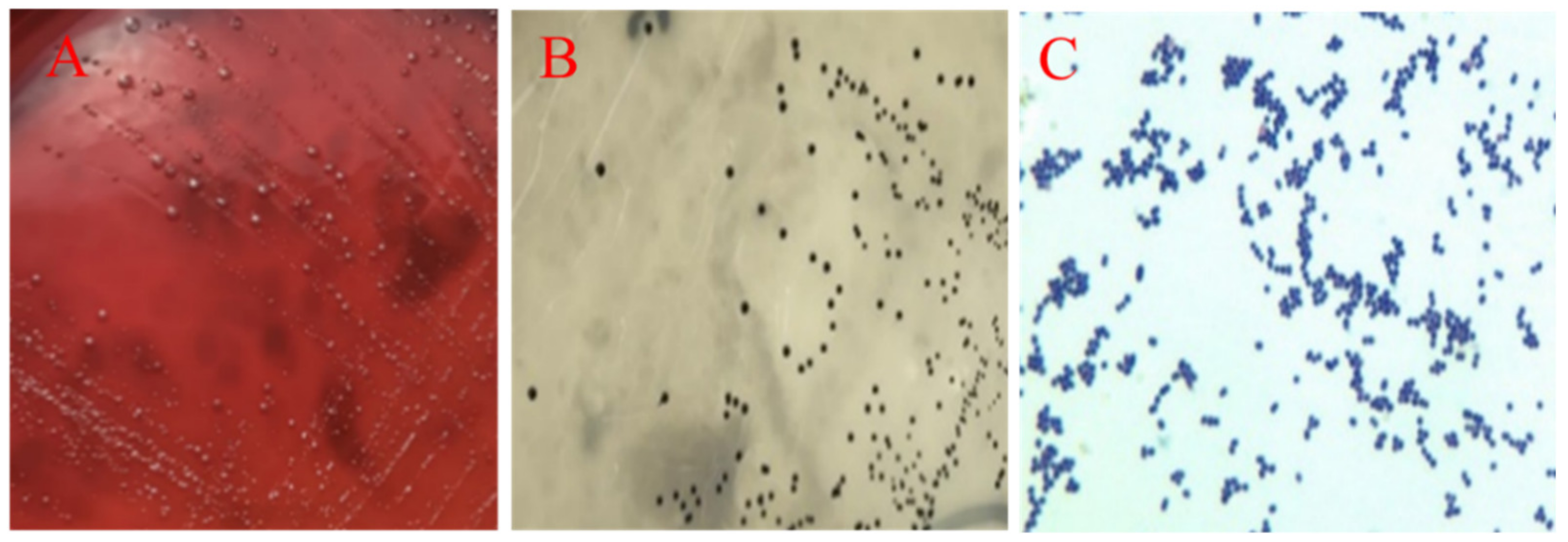
2.2. Pathogens Isolated from the Breeder Ducks with Salpingitis
2.3. Identification and Propagation of E. coli and E. faecalis
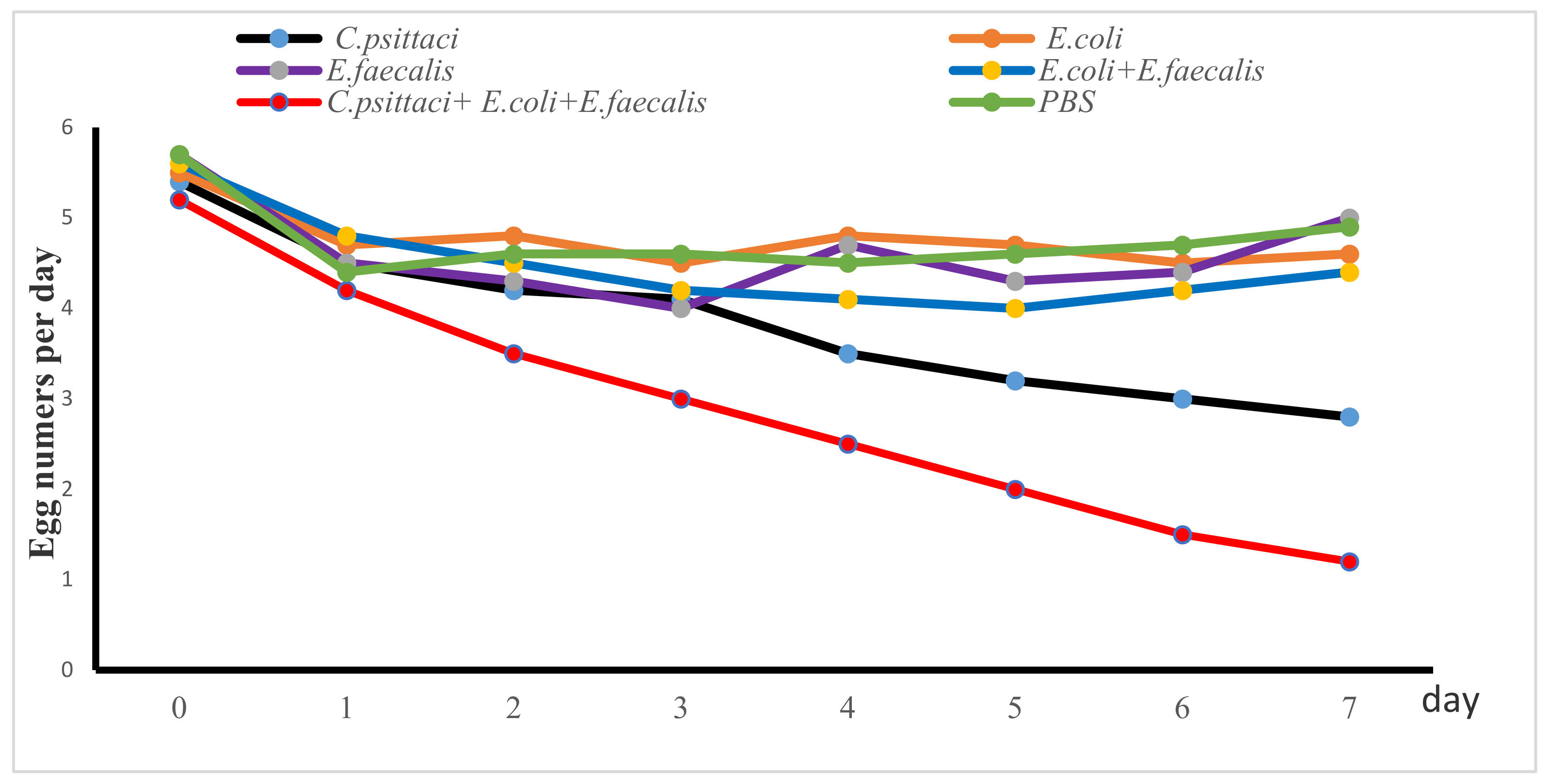
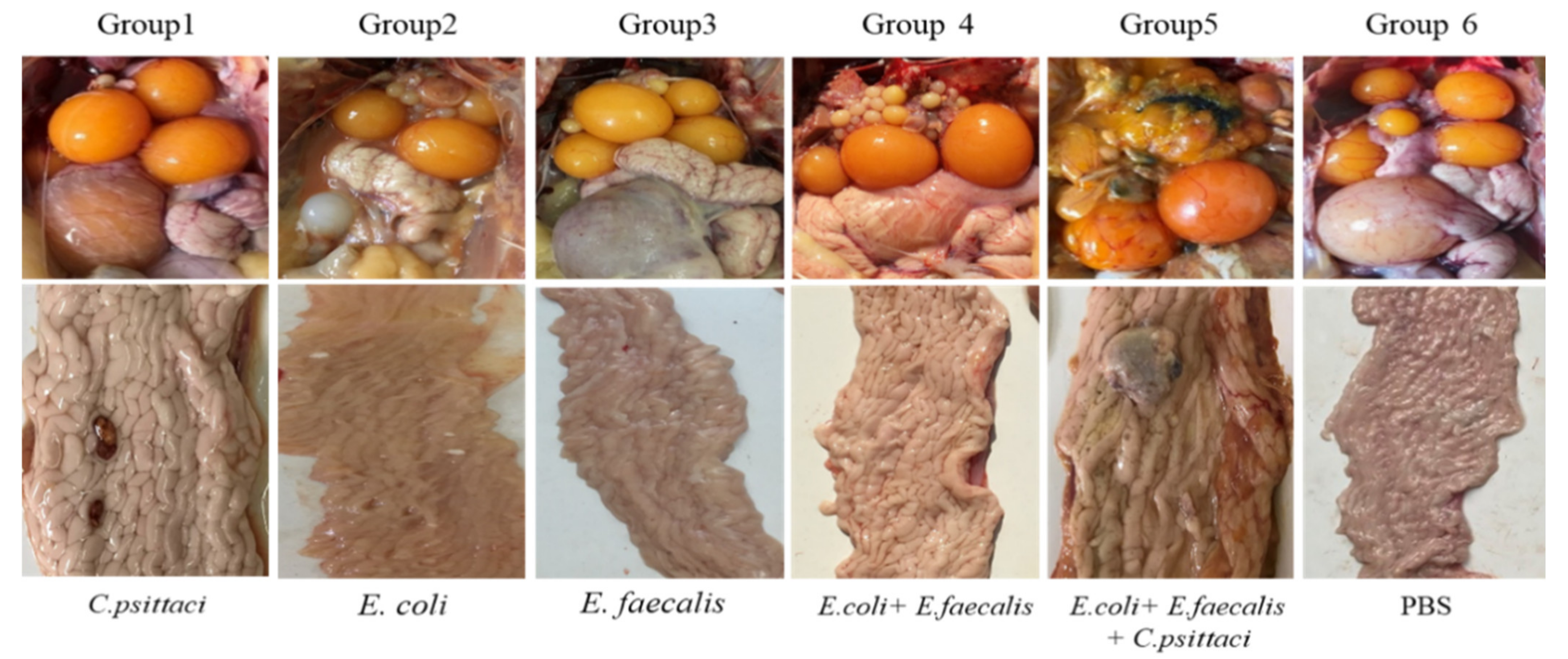
2.4. Salpingitis Induced by E. coli, E. faecalis, C. psittaci or Synergetic Infection in Breeder Layers
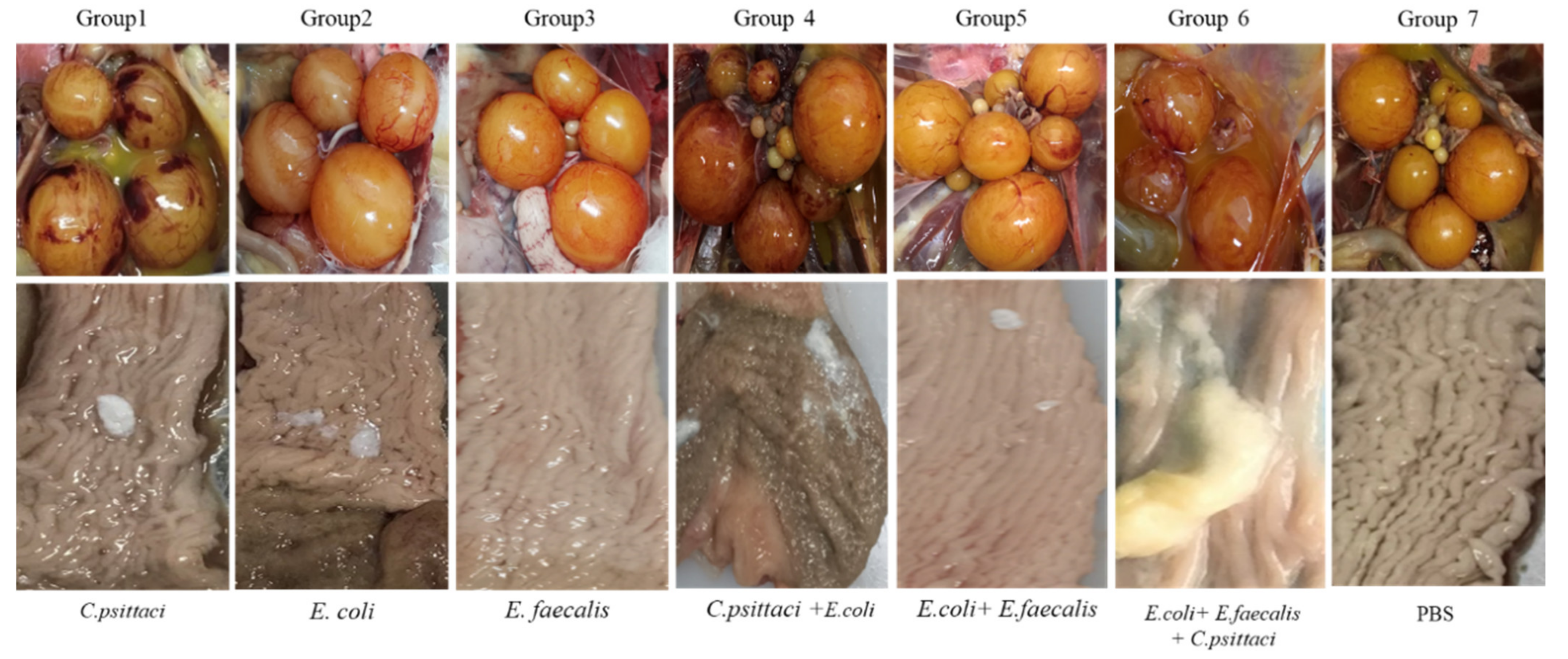
2.5. Salpingitis Induced by E. coli, E. faecalis, C. psittaci or Synergetic Infection in Breeder Ducks
3. Discussion
Multi-Agents of Salpingitis during Production Peak of Laying Hens and Breeder Ducks
4. Materials and Methods
4.1. Seroprevalence of C. psittaci Infection in the Affected Breeder Ducks
4.2. Chlamydial Isolation and Identification
4.3. Identification and Propagation of E. coli and E. faecalis
4.4. Experimental Infection in Breeder Layer
4.5. Experimental Infection in Breeder Ducks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, W.; Chen, T.; Liao, L.; Wang, Z.; Xiao, J.; Lu, J.; Song, C.; Qin, J.; Chen, F.; Chang, Y.F.; et al. A parrot-type Chlamydia psittaci strain is in association with egg production drop in laying ducks. Transb. Emerg. Dis. 2019, 66, 2002–2010. [Google Scholar] [CrossRef]
- Crespo, R.; Walker, R.L.; Nordhausen, R.; Sawyer, S.J.; Manalac, R.B. Salpingitis in Pekin ducks associated with concurrent infection with Tetratrichomonas sp. and Escherichia coli. J. Vet. Diagn. Investig. 2001, 13, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cammayo, P.L.T.; Fernandez-Colorado, C.P.; Flores, R.A.; Roy, A.; Kim, S.; Lillehoj, H.S.; Kim, W.H.; Min, W. IL-17A treatment influences murine susceptibility to experimental Riemerella anatipestifer infection. Dev. Comp. Immun. 2020, 106, 103633. [Google Scholar] [CrossRef] [PubMed]
- Volokhov, D.V.; Grozner, D.; Gyuranecz, M.; Ferguson-Noel, N.; Gao, Y.; Bradbury, J.M.; Whittaker, P.; Chizhikov, V.E.; Szathmary, S.; Stipkovits, L. Mycoplasma anserisalpingitidis sp. nov., isolated from European domestic geese (Anser anser domesticus) with reproductive pathology. Int. J. Syst. Evol. Microbiol. 2020, 70, 2369–2381. [Google Scholar] [CrossRef]
- Gyuranecz, M.; Mitter, A.; Kovacs, A.B.; Grozner, D.; Kreizinger, Z.; Bali, K.; Banyai, K.; Morrow, C.J. Isolation of Mycoplasma anserisalpingitidis from swan goose (Anser cygnoides) in China. BMC Vet. Res. 2020, 16, 178. [Google Scholar] [CrossRef]
- Bisgaard, M. Salpingitis in web-footed birds: Prevalence, aetiology and significance. Avian Pathol. 1995, 24, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, F.T.; Williams, N.J.; Wattret, A.; Jones, T. Observations on salpingitis, peritonitis and salpingoperitonitis in a layer breeder flock. Vet. Rec. 2005, 157, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.H.; Bisgaard, M.; Christensen, J.P.; Kabell, S.; Christensen, H. Pathology and Molecular Characterization of Escherichia coli Associated with the Avian Salpingitis-Peritonitis Disease Syndrome. Avian Dis. 2016, 60, 1–7. [Google Scholar] [CrossRef]
- Zhang, J.J.; Kang, T.Y.; Kwon, T.; Koh, H.; Chandimali, N.; Huynh, D.L.; Wang, X.Z.; Kim, N.; Jeong, D.K. Specific Chicken Egg Yolk Antibody Improves the Protective Response against Gallibacterium anatis Infection. Infect. Immun. 2019, 87, e00619-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Pors, S.E.; Olsen, R.H.; Bojesen, A.M. Transmission and pathogenicity of Gallibacterium anatis and Escherichia coli in embryonated eggs. Vet. Microbiol. 2018, 217, 76–81. [Google Scholar] [CrossRef]
- Mortensen, S.; Johansen, A.E.; Thofner, I.; Christensen, J.P.; Pors, S.E.; Fresno, A.H.; Moller-Jensen, J.; Olsen, J.E. Infectious potential of human derived uropathogenic Escherichia coli UTI89 in the reproductive tract of laying hens. Vet. Microb. 2019, 239, 108445. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, C.; de Souza-Pilz, M.; Bojesen, A.M.; Bisgaard, M.; Hess, M. Tissue distribution of haemolytic Gallibacterium anatis isolates in laying birds with reproductive disorders. Avian Pathol. 2009, 38, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, L.L.; Thofner, I.; Bisgaard, M.; Christensen, J.P.; Olsen, R.H.; Christensen, H. Longitudinal study of transmission of Escherichia coli from broiler breeders to broilers. Vet. Microb. 2017, 207, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Bonfante, F.; Mazzetto, E.; Zanardello, C.; Fortin, A.; Gobbo, F.; Maniero, S.; Bigolaro, M.; Davidson, I.; Haddas, R.; Cattoli, G.; et al. A G1-lineage H9N2 virus with oviduct tropism causes chronic pathological changes in the infundibulum and a long-lasting drop in egg production. Vet. Res. 2018, 49, 83. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, A.A.; Kariyawasam, S. An experimental infection model for Escherichia coli egg peritonitis in layer chickens. Avian Dis. 2014, 58, 25–33. [Google Scholar] [CrossRef]
- Bager, R.J.; Nesta, B.; Pors, S.E.; Soriani, M.; Serino, L.; Boyce, J.D.; Adler, B.; Bojesen, A.M. The fimbrial protein FlfA from Gallibacterium anatis is a virulence factor and vaccine candidate. Infect. Immun. 2013, 81, 1964–1973. [Google Scholar] [CrossRef] [Green Version]
- Landman, W.J.M.; van Eck, J.H.H. The efficacy of inactivated Escherichia coli autogenous vaccines against the E. coli peritonitis syndrome in layers. Avian Pathol. 2017, 46, 658–665. [Google Scholar] [CrossRef]
- Li, L.; Thofner, I.; Christensen, J.P.; Ronco, T.; Pedersen, K.; Olsen, R.H. Evaluation of the efficacy of an autogenous Escherichia coli vaccine in broiler breeders. Avian Pathol. 2017, 46, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Krishnegowda, D.N.; Dhama, K.; Mariappan, A.K.; Munuswamy, P.; Yatoo, M.I.; Tiwari, R.; Karthik, K.; Bhatt, P.; Reddy, M.R. Etiology, epidemiology, pathology, and advances in diagnosis, vaccine development, and treatment of Gallibacterium anatis infection in poultry: A review. Vet. Q. 2020, 40, 16–34. [Google Scholar] [CrossRef] [Green Version]
- El-Adawy, H.; Bocklisch, H.; Neubauer, H.; Hafez, H.M.; Hotzel, H. Identification, differentiation and antibiotic susceptibility of Gallibacterium isolates from diseased poultry. Irish Vet. J. 2018, 71, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Qi, Z.; Jiang, W.; Quan, S.; Sheng, T.; Tu, J.; Shao, Y.; Qi, K. Effects of probiotics on cecal microbiome profile altered by duck Escherichia coli 17 infection in Cherry Valley ducks. Microb. Pathog. 2020, 138, 103849. [Google Scholar] [CrossRef]
- Fossum, O.; Jansson, D.S.; Etterlin, P.E.; Vågsholm, I. Causes of mortality in laying hens in different housing systems in 2001 to 2004. Acta Vet. Scand. 2009, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wittenbrink, M.M.; Mrozek, M.; Bisping, W. Isolation of Chlamydia psittaci from a chicken egg: Evidence of egg transmission. Series B 1993, 40, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Lublin, A.; Shudari, G.; Mechani, S.; Weisman, Y. Egg transmission of Chlamydia psittaci in turkeys. Vet. Rec. 1996, 139, 300. [Google Scholar] [PubMed]
- Vanrompay, D.; Butaye, P.; Vannerom, A.; Ducatelle, R.; Haesebrouck, F. The prevalence of Chlamydia psittaci infections in Belgian commercial turkey poults. Vet. Microb. 1997, 54, 85–93. [Google Scholar] [CrossRef]
- Vorimore, F.; Thebault, A.; Poisson, S.; Cleva, D.; Robineau, J.; de Barbeyrac, B.; Durand, B.; Laroucau, K. Chlamydia psittaci in ducks: A hidden health risk for poultry workers. Pathog. Dis. 2015, 73, 1–9. [Google Scholar] [CrossRef]
- Berrang, M.E.; Cox, N.A.; Frank, J.F.; Buhr, R.J. Bacterial Penetration of the Eggshell and Shell Membranes of the Chicken Hatching Egg: A Review. J. Appl. Poul. Res. 1999, 8, 499–504. [Google Scholar] [CrossRef]
- Ahmed, B.; de Boeck, C.; Dumont, A.; Cox, E.; de Reu, K.; Vanrompay, D. First Experimental Evidence for the Transmission of Chlamydia psittaci in Poultry through Eggshell Penetration. Transb. Emerg. Dis. 2017, 64, 167–170. [Google Scholar] [CrossRef]
- Bruggeman, V.; D’hondt, E.; Berghman, L.; Onagbesan, O.; Vanmontfort, D.; Vandesande, F.; Decuypere, E. The effect of food intake from 2 to 24 weeks of age on LHRH-I content in the median eminence and gonadotrophin levels in pituitary and plasma in female broiler breeder chickens. Gen. Comp. Endocrinol. 1998, 112, 200–209. [Google Scholar] [CrossRef]
- Pal, S.; Theodor, I.; Peterson, E.M.; Delamaza, L.M. Monoclonal immunoglobulin A antibody to the major outer membrane protein of the Chlamydia trachomatis mouse pneumonitis biovar protects mice against a chlamydial genital challenge. Vaccine 1997, 15, 575–582. [Google Scholar] [CrossRef]
- Hafner, L.M. Pathogenesis of fallopian tube damage caused by Chlamydia trachomatis infections. Contraception 2015, 92, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Koroleva, E.A.; Kobets, N.V.; Zayakin, E.S.; Luyksaar, S.I.; Shabalina, L.A.; Zigangirova, N.A. Small Molecule Inhibitor of Type Three Secretion Suppresses Acute and Chronic Chlamydia trachomatis Infection in a Novel Urogenital Chlamydia Model. Biomed. Res. Int. 2015, 2015, 484853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuchardt, L.; Rupp, J. Chlamydia trachomatis as the Cause of Infectious Infertility: Acute, Repetitive or Persistent Long-Term Infection? Curr. Top. Microbiol. Immunol. 2018, 412, 159–182. [Google Scholar] [PubMed]
- Pan, Q.; Liu, A.; He, C. Co-infection of Ornithobacterium rhinotracheale with Streptococcus zooepidemicus in chickens. Avian Dis. 2012, 56, 680–684. [Google Scholar] [CrossRef]
- Chu, J.; Zhang, Q.; Zhang, T.; Han, E.; Zhao, P.; Khan, A.; He, C.; Wu, Y. Chlamydia psittaci infection increases mortality of avian influenza virus H9N2 by suppressing host immune response. Sci Rep. 2016, 6, 29421. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, A.B.; Kreizinger, Z.; Forro, B.; Grozner, D.; Mitter, A.; Marton, S.; Bali, K.; Sawicka, A.; Tomczyk, G.; Banyai, K.; et al. The core genome multi-locus sequence typing of Mycoplasma anserisalpingitidis. BMC Genom. 2020, 21, 403. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.H.; Thofner, I.C.; Pors, S.E.; Dos Santos, T.P.; Christensen, J.P. Experimental induced avian E. coli salpingitis: Significant impact of strain and host factors on the clinical and pathological outcome. Vet. Micro. 2016, 188, 59–66. [Google Scholar] [CrossRef]
- Elbestawy, A.R.; Ellakany, H.F.; Abd El-Hamid, H.S.; Bekheet, A.A.; Mataried, N.E.; Nasr, S.M.; Amarin, N.M. Isolation, characterization, and antibiotic sensitivity assessment of Gallibacterium anatis biovar haemolytica, from diseased Egyptian chicken flocks during the years 2013 and 2015. Poultry Sci. 2018, 97, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Geens, T.; Desplanques, A.; van Loock, M.; Bonner, B.M.; Kaleta, E.F.; Magnino, S.; Andersen, A.A.; Everett, K.D.; Vanrompay, D. Sequencing of the Chlamydophila psittaci ompA gene reveals a new genotype, E/B, and the need for a rapid discriminatory genotyping method. J. Clin. Microbiol. 2005, 43, 2456–2461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Li, S.; Yang, J.; Pang, W.; Yang, L.; He, C. Isolation and characterization of Chlamydophila psittaci isolated from laying hens with cystic oviducts. Avian Dis. 2008, 52, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehricht, R.; Slickers, P.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes. 2006, 20, 60–63. [Google Scholar] [CrossRef]
- Vogler, B.R.; Trinkler, M.; Marti, H.; Borel, N.; Pesch, T.; Prahauser, B.; Hoop, R.; Mattmann, P.; Albini, S. Survey on Chlamydiaceae in cloacal swabs from Swiss turkeys demonstrates absence of Chlamydia psittaci and low occurrence of Chlamydia gallinacean. PLoS ONE 2019, 14, e0226091. [Google Scholar] [CrossRef]
- Smith, K.A.; Bradley, K.K.; Stobierski, M.G.; Tengelsen, L.A. Compendium of measures to control Chlamydophila psittaci (formerly Chlamydia psittaci) infection among humans (psittacosis) and pet birds. Javma-J. Am. Vet. Med. A 2005, 226, 532–539. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeder Ducks | Clinical Health Flock | Suspected Flock | ||
|---|---|---|---|---|
| Sheldrake | Muscovy | Sheldrake | Muscovy | |
| Flocks | 5 | 6 | 7 | 8 |
| No. breeder ducks | 2100 | 2400 | 2800 | 3200 |
| Egg production (%) | 82.5 | 85.7 | 49.5 | 31.2 |
| No. blood samples | 50 | 60 | 70 | 80 |
| No. positive samples | 5 | 7 | 45 | 68 |
| No. negative samples | 45 | 53 | 25 | 12 |
| Positive (%) | 10.0 | 11.6 | 64.2 | 85.0 |
| Group | Inoculates | Chlamydia (IFU/g) | E. coli (CFU/g) | E. faecalis (CFU/g) |
|---|---|---|---|---|
| 1 | C. psittaci | 1.0 × 105 | -- | -- |
| 2 | E. coli | -- | 1.0 × 106 | -- |
| 3 | E. faecalis | -- | -- | 1.0 × 104 |
| 4 | E. coli + E. faecalis | -- | 1.1 × 108 | 1.0 × 108 |
| 5 | C. psittaci + E. coli + E. faecalis | 0.7 × 105 | 2.0 × 108 | 2.1 × 108 |
| 6 | PBS | -- | 1.0 × 102 | -- |
| Group | Inoculates | Chlamydia (IFU/g) | E. coli (CFU/g) | E. faecalis (CFU/g) |
|---|---|---|---|---|
| 1 | C. psittaci | 2.5 × 105 | -- | -- |
| 2 | E. coli | -- | 2.3 × 106 | -- |
| 3 | E.faecalis | -- | -- | 1.5 × 106 |
| 4 | E. coli + E. faecalis | -- | 1.2 × 108 | 2.6 × 107 |
| 5 | E. coli + C. psittaci | 3.1 × 106 | 1.2 × 108 | -- |
| 6 | C. psittaci + E. coli + E. faecalis | 1.3 × 106 | 2.0 × 108 | 1.1 × 108 |
| 7 | PBS | -- | 2.2 × 102 | -- |
| Primers | Sequence (5′→3′) | Products | References |
|---|---|---|---|
| 23S rRNA | CTGAAACCAGTAGCTTATAAGCGGT ACCTCGCCGTTTAACTTAACTCC FAM-CTCATCATGCAAAAGGCACGCCG-TAMRA | 111 bp | Ralf Ehricht et al., 2006 [41] |
| C. psittaci-incA | GCCATCATGCTTGTTTCGTTT
CGGCGTGCCACTTGAGA FAM-TCATTGTCATTATGGTGATTCAGGA-TAMRA | 74 bp | Vogler BR, Trinkler M, et al., 2019 [42] |
| ompA | ATGAAAAAACTCTTGAAATCG TTAGAATCTGAATTGAGCYTTCATYT | 1209 bp | Smith KA, Bradley KK et al., 2005 [43] |
| Strain | Genotype | District | Host | GenBank No. |
|---|---|---|---|---|
| MDHL05 | A | Taiwan, China | Muscovy duck | MK032046.1 |
| MDYL09B | A | Taiwan, China | Muscovy duck | MK032048.1 |
| MDPT06B | A | Taiwan, China | Muscovy duck | MK032049.1 |
| MDPT08 | A | Taiwan, China | Muscovy duck | MK032050.1 |
| MDSK10 | A | Taiwan, China | Muscovy duck | MK032051.1 |
| SZ-18-1 | A | China | Duck | MK751470.1 |
| CS1 | C | China | Duck | EU009493.1 |
| CAU1 | A | China | Duck | EU101714.1 |
| CAU2 | A | China | Chicken | EF202608.1 |
| SP12 | A | China | Bird | EU856032.1 |
| 06-859/1 | E/B | France | Duck | EU159263.2 |
| WS/RT/E30 | E/B | Germany | Duck | AY762613.1 |
| GD | C | Germany | Duck | AF269261.1 |
| VG65-22-3 | E/B | Germany | Duck | EU019091.1 |
| 90/1051 | A | Belgium | Parrot | AY762608.1 |
| 6BC | A | America | Parrot | X56980.1 |
| 41A12 | B | Belgium | Turkey | AY762609.1 |
| 3759/2 | E | Italy | Pigeon | AY762611.1 |
| M56 | M56 | America | Hares and muskrats | AF269268.1 |
| WC | WC | America | Cow | AF269269.1 |
| 7344/2 | D | Italy | Pigeon | AY762610.1 |
| CT1 | C | America | Turkey | AF269260.1 |
| 7778B15 | F | Belgium | Turkey | AY762612.1 |
| GPIC | Chlamydia caviae | America | Guinea pig | AF269282.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, H.; Quan, H.; Zhang, Y.; Li, Q.; Wang, Y.; Yuan, S.; Huang, S.; He, C. Co-Infection of Escherichia coli, Enterococcus faecalis and Chlamydia psittaci Contributes to Salpingitis of Laying Layers and Breeder Ducks. Pathogens 2021, 10, 755. https://doi.org/10.3390/pathogens10060755
Fang H, Quan H, Zhang Y, Li Q, Wang Y, Yuan S, Huang S, He C. Co-Infection of Escherichia coli, Enterococcus faecalis and Chlamydia psittaci Contributes to Salpingitis of Laying Layers and Breeder Ducks. Pathogens. 2021; 10(6):755. https://doi.org/10.3390/pathogens10060755
Chicago/Turabian StyleFang, Huanxin, Hongkun Quan, Yuhang Zhang, Qiang Li, Yihui Wang, Sheng Yuan, Shujian Huang, and Cheng He. 2021. "Co-Infection of Escherichia coli, Enterococcus faecalis and Chlamydia psittaci Contributes to Salpingitis of Laying Layers and Breeder Ducks" Pathogens 10, no. 6: 755. https://doi.org/10.3390/pathogens10060755
APA StyleFang, H., Quan, H., Zhang, Y., Li, Q., Wang, Y., Yuan, S., Huang, S., & He, C. (2021). Co-Infection of Escherichia coli, Enterococcus faecalis and Chlamydia psittaci Contributes to Salpingitis of Laying Layers and Breeder Ducks. Pathogens, 10(6), 755. https://doi.org/10.3390/pathogens10060755