
Rapid Generation of Coronaviral Immunity Using Recombinant Peptide Modified Nanodiamonds

 ,
,  , , , ,
, , , ,  and
and 
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Properties of Nanodiamond-Based Nanostructures as Vaccine Adjuvant

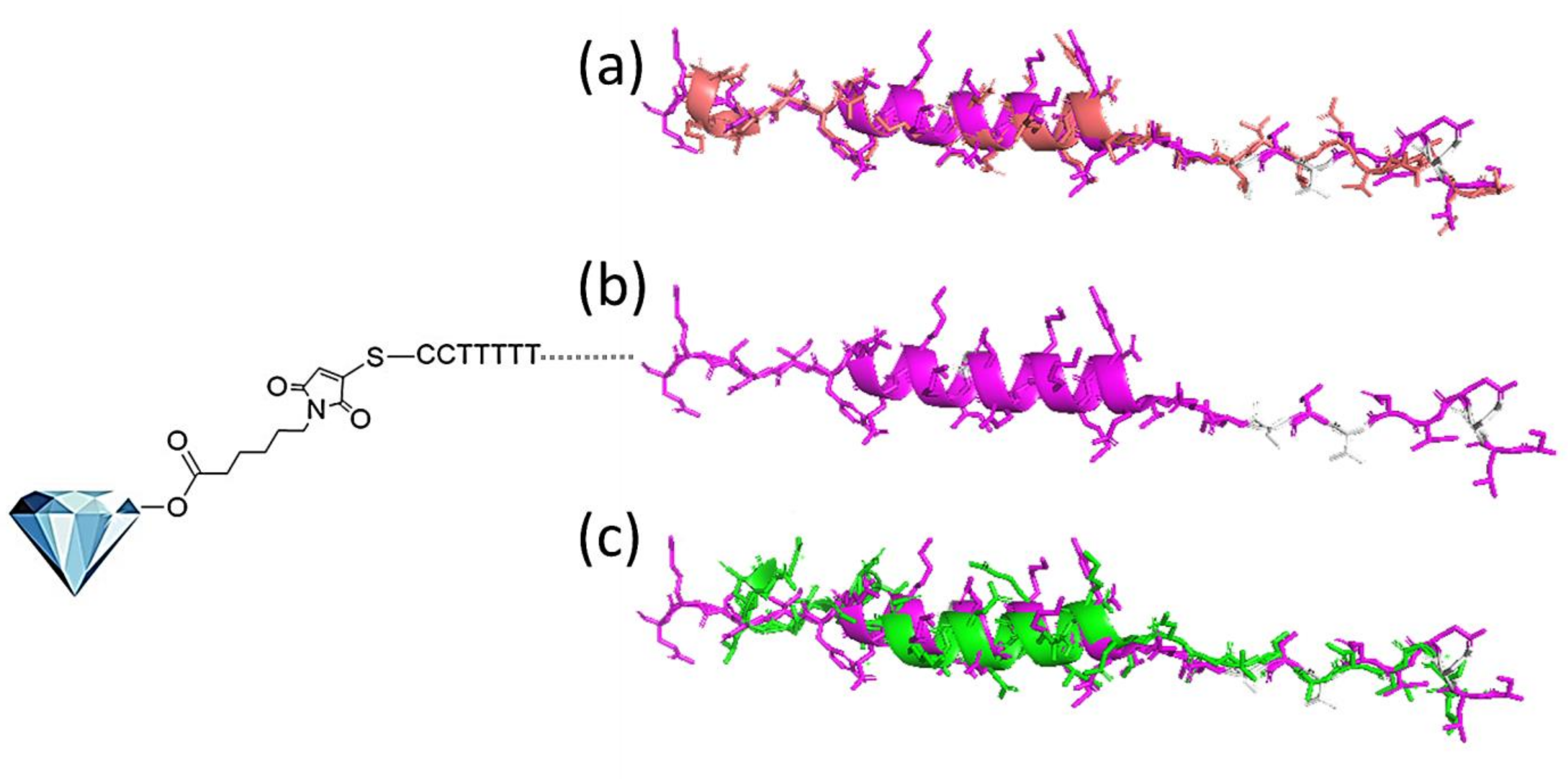
2.2. Integration of HR2 Mimicking Peptide
2.3. Immune Response to ND-Pancoronaviral Peptide coNjugate in Mice
2.4. Activation of Neutrophil Extracellular Trap and Subsequent Sequestration
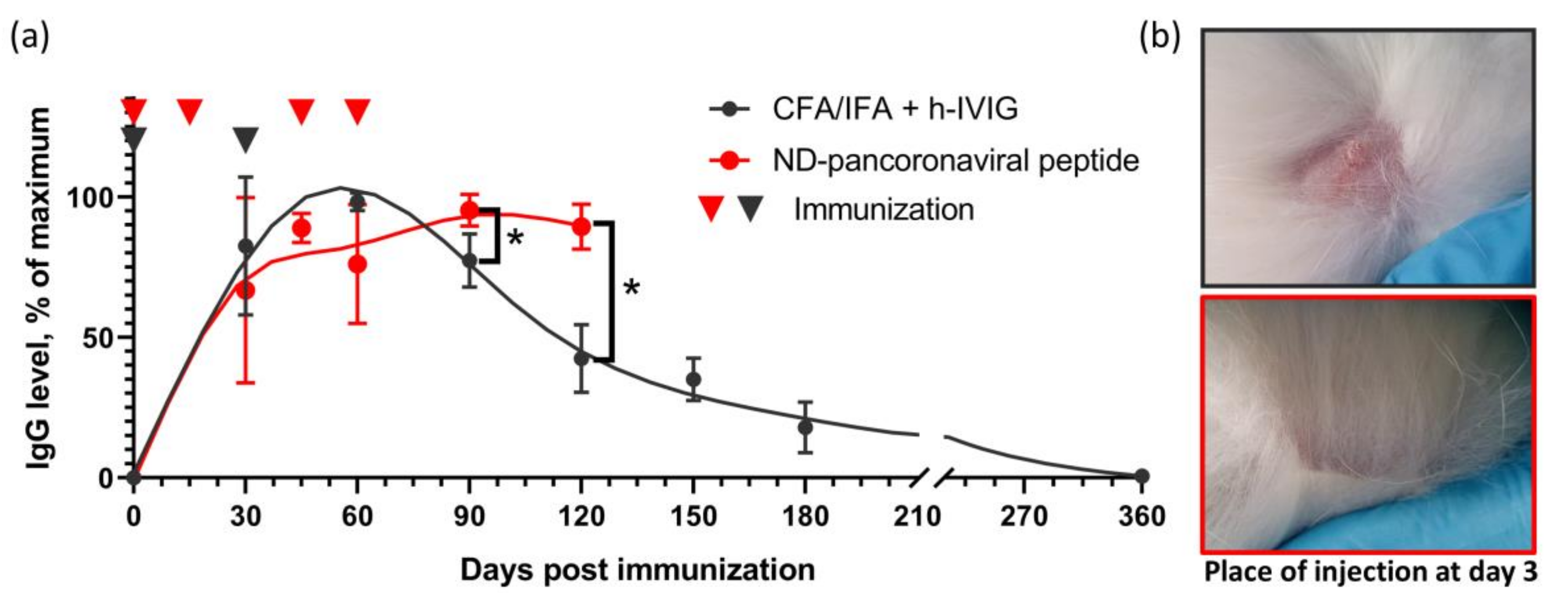
2.5. Duration of Immune Response in Rabbits
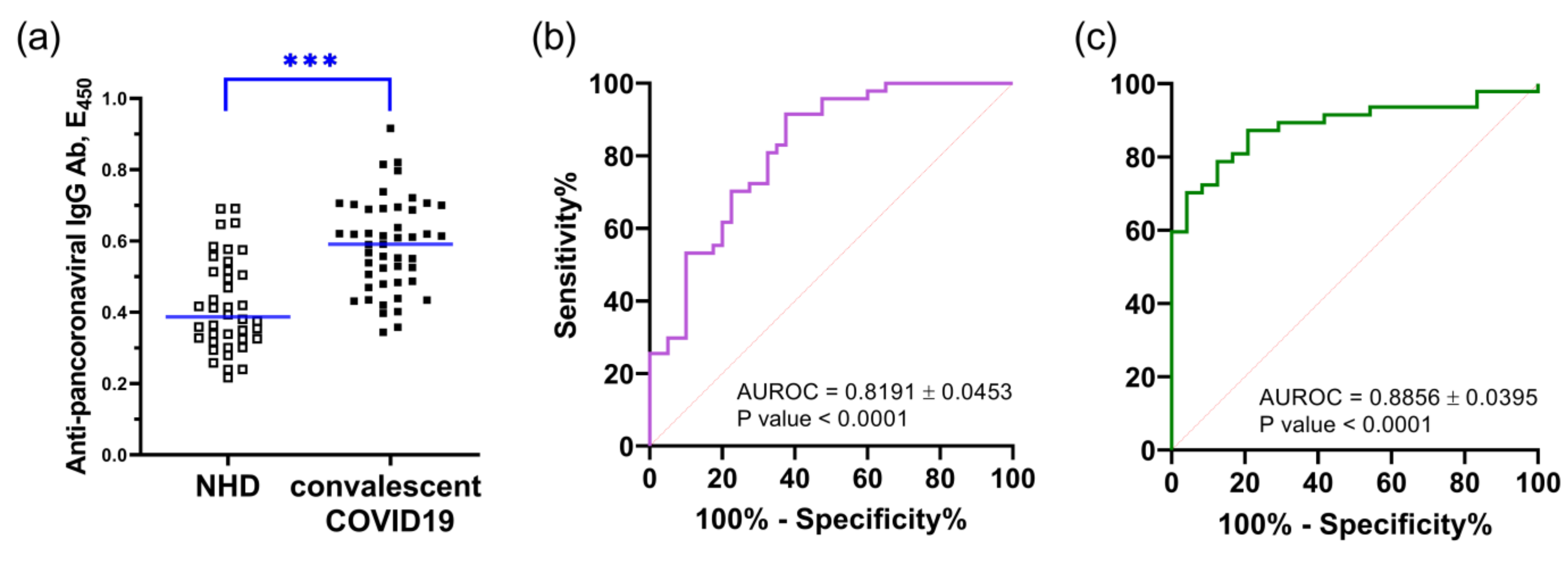
2.6. Cross-Reactivity of Peptide with Human Convalescent COVID Sera
3. Conclusions
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plotkina, S.A. Vaccination against the major infectious diseases. Comptes Rendus l’Académie des Sci.-Ser. III-Sci. la Vie 1999, 322, 943–951. [Google Scholar] [CrossRef]
- Shen, A.K.; Cooke, M.T. Infectious disease vaccines. Nat. Rev. Drug Discov. 2019, 18, 169–170. [Google Scholar] [CrossRef]
- Arvin, A.M.; Fink, K.; Schmid, M.A.; Cathcart, A.; Spreafico, R.; Havenar-Daughton, C.; Lanzavecchia, A.; Corti, D.; Virgin, H.W. A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature 2020, 584, 353–363. [Google Scholar] [CrossRef]
- Welsh, R.M.; Fujinami, R.S. Pathogenic epitopes, heterologous immunity and vaccine design. Nat. Rev. Microbiol. 2007, 5, 555–563. [Google Scholar] [CrossRef] [PubMed]
- van Riel, D.; de Wit, E. Next-generation vaccine platforms for COVID-19. Nat. Mater. 2020, 19, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef]
- Maiti, D.; Tong, X.; Mou, X.; Yang, K. Carbon-Based Nanomaterials for Biomedical Applications: A Recent Study. Front. Pharmacol. 2019, 9, 1401. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.D.; Singh, R.K.; Kim, H.-W. Carbon-based nanomaterials as an emerging platform for theranostics. Mater. Horizons 2019, 6, 434–469. [Google Scholar] [CrossRef]
- Cha, C.; Shin, S.R.; Annabi, N.; Dokmeci, M.R.; Khademhosseini, A. Carbon-Based Nanomaterials: Multifunctional Materials for Biomedical Engineering. ACS Nano 2013, 7, 2891–2897. [Google Scholar] [CrossRef]
- Nekoueian, K.; Amiri, M.; Sillanpää, M.; Marken, F.; Boukherroub, R.; Szunerits, S. Carbon-based quantum particles: An electroanalytical and biomedical perspective. Chem. Soc. Rev. 2019, 48, 4281–4316. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, W.; Li, J.; Tao, L.; Wei, Y. A comparative study of cellular uptake and cytotoxicity of multi-walled carbon nanotubes, graphene oxide, and nanodiamond. Toxicol. Res. 2012, 1, 62–68. [Google Scholar] [CrossRef]
- Mochalin, V.N.; Shenderova, O.; Ho, D.; Gogotsi, Y. The properties and applications of nanodiamonds. Nat. Nanotechnol. 2012, 7, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Barras, A.; Martin, F.A.; Bande, O.; Baumann, J.-S.; Ghigo, J.-M.; Boukherroub, R.; Beloin, C.; Siriwardena, A.; Szunerits, S. Glycan-functionalized diamond nanoparticles as potent E. coli anti-adhesives. Nanoscale 2013, 5, 2307. [Google Scholar] [CrossRef]
- Turcheniuk, V.; Turcheniuk, K.; Bouckaert, J.; Barras, A.; Dumych, T.; Bilyy, R.; Zaitsev, V.; Siriwardena, A.; Wang, Q.; Boukherroub, R.; et al. Affinity of Glycan-Modified Nanodiamonds towards Lectins and Uropathogenic Escherichia Coli. ChemNanoMat 2016, 2, 307–314. [Google Scholar] [CrossRef]
- Khanal, M.; Larsonneur, F.; Raks, V.; Barras, A.; Baumann, J.S.; Martin, F.A.; Boukherroub, R.; Ghigo, J.M.; Ortiz Mellet, C.; Zaitsev, V.; et al. Inhibition of type 1 fimbriae-mediated Escherichia coli adhesion and biofilm formation by trimeric cluster thiomannosides conjugated to diamond nanoparticles. Nanoscale 2015, 7, 2325–2335. [Google Scholar] [CrossRef]
- Khanal, M.; Vausselin, T.; Barras, A.; Bande, O.; Turcheniuk, K.; Benazza, M.; Zaitsev, V.; Teodorescu, C.M.; Boukherroub, R.; Siriwardena, A.; et al. Phenylboronic-Acid-Modified Nanoparticles: Potential Antiviral Therapeutics. ACS Appl. Mater. Interfaces 2013, 5, 12488–12498. [Google Scholar] [CrossRef] [PubMed]
- Irvine, D.J.; Hanson, M.C.; Rakhra, K.; Tokatlian, T. Synthetic Nanoparticles for Vaccines and Immunotherapy. Chem. Rev. 2015, 115, 11109–11146. [Google Scholar] [CrossRef]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.-X.; Mitter, N.; Yu, C.; Middelberg, A.P.J. Nanoparticle vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef]
- Kossovsky, N.; Gelman, A.; Hnatyszyn, H.J.; Rajguru, S.; Garrell, R.L.; Torbati, S.; Freitas, S.S.F.; Chow, G.-M. Surface-Modified Diamond Nanoparticles as Antigen Delivery Vehicles. Bioconjug. Chem. 1995, 6, 507–511. [Google Scholar] [CrossRef]
- Pham, N.B.; Ho, T.T.; Nguyen, G.T.; Le, T.T.; Le, N.T.; Chang, H.C.; Pham, M.D.; Conrad, U.; Chu, H.H. Nanodiamond enhances immune responses in mice against recombinant HA/H7N9 protein. J. Nanobiotechnology 2017, 15, 1–12. [Google Scholar] [CrossRef]
- Muñoz, L.E.; Bilyy, R.; Biermann, M.H.C.; Kienhöfer, D.; Maueröder, C.; Hahn, J.; Brauner, J.M.; Weidner, D.; Chen, J.; Scharin-Mehlmann, M.; et al. Nanoparticles size-dependently initiate self-limiting NETosis-driven inflammation. Proc. Natl. Acad. Sci. 2016, 113, E5856–E5865. [Google Scholar] [CrossRef]
- Bilyy, R.; Bila, G.; Vishchur, O.; Vovk, V.; Herrmann, M. Neutrophils as Main Players of Immune Response Towards Nondegradable Nanoparticles. Nanomaterials 2020, 10, 1273. [Google Scholar] [CrossRef]
- Bilyy, R.; Paryzhak, S.; Turcheniuk, K.; Dumych, T.; Barras, A.; Boukherroub, R.; Wang, F.; Yushin, G.; Szunerits, S. Aluminum oxide nanowires as safe and effective adjuvants for next-generation vaccines. Mater. Today 2019, 22, 58–66. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Q.; Zhu, Y.; Chan, K.-H.H.; Qin, L.; Li, Y.; Wang, Q.; Chan, J.F.-W.W.; Du, L.; Yu, F.; et al. Structure-based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor. Nat. Commun. 2014, 5, 3067. [Google Scholar] [CrossRef]
- Rabets, A.; Bila, G.; Grytsko, R.; Samborsky, M.; Rebets, Y.; Vari, S.; Pagneux, Q.; Barras, A.; Boukherroub, R.; Szunerits, S.; et al. Development of antibodies to pan-coronavirus spike peptides in convalescent COVID-19 patients. medRxiv 2020. [Google Scholar] [CrossRef]
- Rabets, A.; Bila, G.; Grytsko, R.; Samborskyy, M.; Rebets, Y.; Vari, S.G.; Pagneux, Q.; Barras, A.; Boukherroub, R.; Szunerits, S.; et al. The Potential of Developing Pan-Coronaviral Antibodies to Spike Peptides in Convalescent COVID-19 Patients. Arch. Immunol. Ther. Exp. 2021, 69, 5. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Cao, D.; Zhang, Y.; Ma, J.; Qi, J.; Wang, Q.; Lu, G.; Wu, Y.; Yan, J.; Shi, Y.; et al. Cryo-EM structures of MERS-CoV and SARS-CoV spike glycoproteins reveal the dynamic receptor binding domains. Nat. Commun. 2017, 8, 15092. [Google Scholar] [CrossRef] [PubMed]
- Łoczechin, A.; Séron, K.; Barras, A.; Giovanelli, E.; Belouzard, S.; Chen, Y.-T.; Metzler-Nolte, N.; Boukherroub, R.; Dubuisson, J.; Szunerits, S. Functional Carbon Quantum Dots as Medical Countermeasures to Human Coronavirus. ACS Appl. Mater. Interfaces 2019, 11, 42964–42974. [Google Scholar] [CrossRef]
- Huang, X.; Li, M.; Xu, Y.; Zhang, J.; Meng, X.; An, X.; Sun, L.; Guo, L.; Shan, X.; Ge, J.; et al. Novel Gold Nanorod-Based HR1 Peptide Inhibitor for Middle East Respiratory Syndrome Coronavirus. ACS Appl. Mater. Interfaces 2019, 11, 19799–19807. [Google Scholar] [CrossRef] [PubMed]
- Williams, O.A.; Hees, J.; Dieker, C.; Jäger, W.; Kirste, L.; Nebel, C.E. Size-Dependent Reactivity of Diamond Nanoparticles. ACS Nano 2010, 4, 4824–4830. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Z.; Welsher, K.; Robinson, J.T.; Goodwin, A.; Zaric, S.; Dai, H. Nano-graphene oxide for cellular imaging and drug delivery. Nano Res. 2008, 1, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Siriwardena, A.; Khanal, M.; Barras, A.; Bande, O.; Mena-Barragan, T.; Ortiz Mellet, C.; Garcia Fernandez, J.M.; Boukherroub, R.; Szunerits, S. Unprecedented inhibition of glycosidase-catalyzed substrate hydrolysis by nanodiamond-grafted O-glycosides. RSC Adv. 2015, 5, 100568–100578. [Google Scholar] [CrossRef]
- Kong, N.; Jiang, T.; Zhou, Z.; Fu, J. Cytotoxicity of polymerized resin cements on human dental pulp cells in vitro. Dent. Mater. 2009, 25, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Biermann, M.H.C.; Podolska, M.J.; Knopf, J.; Reinwald, C.; Weidner, D.; Maueröder, C.; Hahn, J.; Kienhöfer, D.; Barras, A.; Boukherroub, R.; et al. Oxidative burst-dependent NETosis is implicated in the resolution of necrosis-associated sterile inflammation. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, Y.; Chen, Y.; Wu, B.; Qin, K.; Zhao, J.; Lou, Y.; Tan, W. Discovery of a novel canine respiratory coronavirus support genetic recombination among betacoronavirus1. Virus Res. 2017, 237, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Stümer, J.; Biermann, M.H.C.H.C.; Knopf, J.; Magorivska, I.; Kastbom, A.; Svärd, A.; Janko, C.; Bilyy, R.; Schett, G.; Sjöwall, C.; et al. Altered glycan accessibility on native immunoglobulin G complexes in early rheumatoid arthritis and its changes during therapy. Clin. Exp. Immunol. 2017, 189, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Platt, J.L.; Cascalho, M. IgM in the kidney: A multiple personality disorder. Kidney Int. 2015, 88, 439–441. [Google Scholar] [CrossRef]
- Allen, I.C. Delayed-type hypersensitivity models in mice. Methods Mol Biol 2013, 1031, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science (80-.) 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Schauer, C.; Janko, C.; Munoz, L.E.L.E.; Zhao, Y.Y.; Kienhöfer, D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; et al. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Stephen, J.; Scales, H.E.; Benson, R.A.; Erben, D.; Garside, P.; Brewer, J.M. Neutrophil swarming and extracellular trap formation play a significant role in Alum adjuvant activity. NPJ Vaccines 2017, 2, 1. [Google Scholar] [CrossRef]
- Zost, S.J.; Gilchuk, P.; Case, J.B.; Binshtein, E.; Chen, R.E.; Nkolola, J.P.; Schäfer, A.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lai, D.; Zhang, H.; Jiang, H.; Tian, X.; Ma, M.; Qi, H.; Meng, Q.; Guo, S.; Wu, Y.; et al. Linear epitopes of SARS-CoV-2 spike protein elicit neutralizing antibodies in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 1095–1097. [Google Scholar] [CrossRef]
- Zhu, S.; Meng, Q.; Wang, L.; Zhang, J.; Song, Y.; Jin, H.; Zhang, K.; Sun, H.; Wang, H.; Yang, B. Highly Photoluminescent Carbon Dots for Multicolor Patterning, Sensors, and Bioimaging. Angew. Chemie Int. Ed. 2013, 52, 3953–3957. [Google Scholar] [CrossRef]
- Belouzard, S.; Chu, V.C.; Whittaker, G.R. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc. Natl. Acad. Sci. USA 2009, 106, 5871–5876. [Google Scholar] [CrossRef]
- Turcheniuk, K.; Dumych, T.; Bilyy, R.; Turcheniuk, V.; Bouckaert, J.; Vovk, V.; Chopyak, V.; Zaitsev, V.; Mariot, P.; Prevarskaya, N.; et al. Plasmonic photothermal cancer therapy with gold nanorods/reduced graphene oxide core/shell nanocomposites. RSC Adv. 2016, 6, 1600–1610. [Google Scholar] [CrossRef]
- ThermoFisherScientific Custom Rabbit Polyclonal Antibody Production Protocols. Available online: https://www.thermofisher.com/ua/en/home/life-science/antibodies/custom-antibodies/custom-antibody-production/custom-polyclonal-antibody-production/custom-rabbit-polyclonal-antibody-production-protocols.html (accessed on 2 July 2019).
- Sin, Y.M.; Sedgwick, A.D.; Chea, E.P.; Willoughby, D.A. Mast cells in newly formed lining tissue during acute inflammation: A six day air pouch model in the mouse. Ann. Rheum. Dis. 1986, 45, 873–877. [Google Scholar] [CrossRef]
- Bila, G.; Schneider, M.; Peshkova, S.; Krajnik, B.; Besh, L.; Lutsyk, A.; Matsyura, O.; Bilyy, R. Novel approach for discrimination of eosinophilic granulocytes and evaluation of their surface receptors in a multicolor fluorescent histological assessment. Ukr. Biochem. J. 2020, 92, 99–106. [Google Scholar] [CrossRef]
- Kiessig, S.; Abel, U.; Risse, P.; Friedrich, J.; Heinz, F.; Kunz, C. Problems of cut-off level determination in enzyme immunoassays: The case of TBE-ELISA. Klin. Lab. 1993, 39, 877–886. [Google Scholar]
- Crowther, J.R. The ELISA Guidebook; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2009; Volume 516, ISBN 978-1-60327-253-7. [Google Scholar]
- Biermann, M.H.C.C.; Boeltz, S.; Pieterse, E.; Knopf, J.; Rech, J.; Bilyy, R.; van der Vlag, J.; Tincani, A.; Distler, J.H.W.W.; Krönke, G.; et al. Autoantibodies Recognizing Secondary NEcrotic Cells Promote Neutrophilic Phagocytosis and Identify Patients With Systemic Lupus Erythematosus. Front. Immunol. 2018, 9, 989. [Google Scholar] [CrossRef] [PubMed]
- Bozhenko, M.; Boichuk, M.; Bila, G.; Nehrych, T.; Bilyy, R. Freezing influences, the exposure of IgG glycans in sera from multiple sclerosis patients. Ukr. Biochem. J. 2020, 92, 21–31. [Google Scholar] [CrossRef]
- Brandt, B.W.; Heringa, J.; Leunissen, J.A.M. SEQATOMS: A web tool for identifying missing regions in PDB in sequence context. Nucleic Acids Res. 2008, 36, W255–W259. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | ζ (mV) 1 | Size (nm) | Hydrodynamic Size (nm) 2 | PDI 1 | C1s 3 (at. %) | O1s (at. %) | N1s (at. %) |
|---|---|---|---|---|---|---|---|
| ND | +35.3 ± 1.6 | 14.0 ± 5.0 | 89 ± 13 | 0.246 ± 0.002 | 88.4 | 10.1 | 1.5 |
| GO-PEG | −25.3± 2.4 | >200 | - | - | 75.4 | 22.3 | 2.3 |
| CQD | −9.9 ± 3.4 | 5.1 ± 0.2 | 11 ± 0.1 | 0.220 ± 0.110 | 72.6 | 12.5 | 14.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilyy, R.; Pagneux, Q.; François, N.; Bila, G.; Grytsko, R.; Lebedin, Y.; Barras, A.; Dubuisson, J.; Belouzard, S.; Séron, K.; et al. Rapid Generation of Coronaviral Immunity Using Recombinant Peptide Modified Nanodiamonds. Pathogens 2021, 10, 861. https://doi.org/10.3390/pathogens10070861
Bilyy R, Pagneux Q, François N, Bila G, Grytsko R, Lebedin Y, Barras A, Dubuisson J, Belouzard S, Séron K, et al. Rapid Generation of Coronaviral Immunity Using Recombinant Peptide Modified Nanodiamonds. Pathogens. 2021; 10(7):861. https://doi.org/10.3390/pathogens10070861
Chicago/Turabian StyleBilyy, Rostyslav, Quentin Pagneux, Nathan François, Galyna Bila, Roman Grytsko, Yuri Lebedin, Alexandre Barras, Jean Dubuisson, Sandrine Belouzard, Karin Séron, and et al. 2021. "Rapid Generation of Coronaviral Immunity Using Recombinant Peptide Modified Nanodiamonds" Pathogens 10, no. 7: 861. https://doi.org/10.3390/pathogens10070861
APA StyleBilyy, R., Pagneux, Q., François, N., Bila, G., Grytsko, R., Lebedin, Y., Barras, A., Dubuisson, J., Belouzard, S., Séron, K., Boukherroub, R., & Szunerits, S. (2021). Rapid Generation of Coronaviral Immunity Using Recombinant Peptide Modified Nanodiamonds. Pathogens, 10(7), 861. https://doi.org/10.3390/pathogens10070861