
Genotypic Characterization of Clinical Isolates of Staphylococcus aureus from Pakistan




Abstract
:1. Introduction
2. Results
2.1. Antimicrobial Susceptibility and Resistance Genes
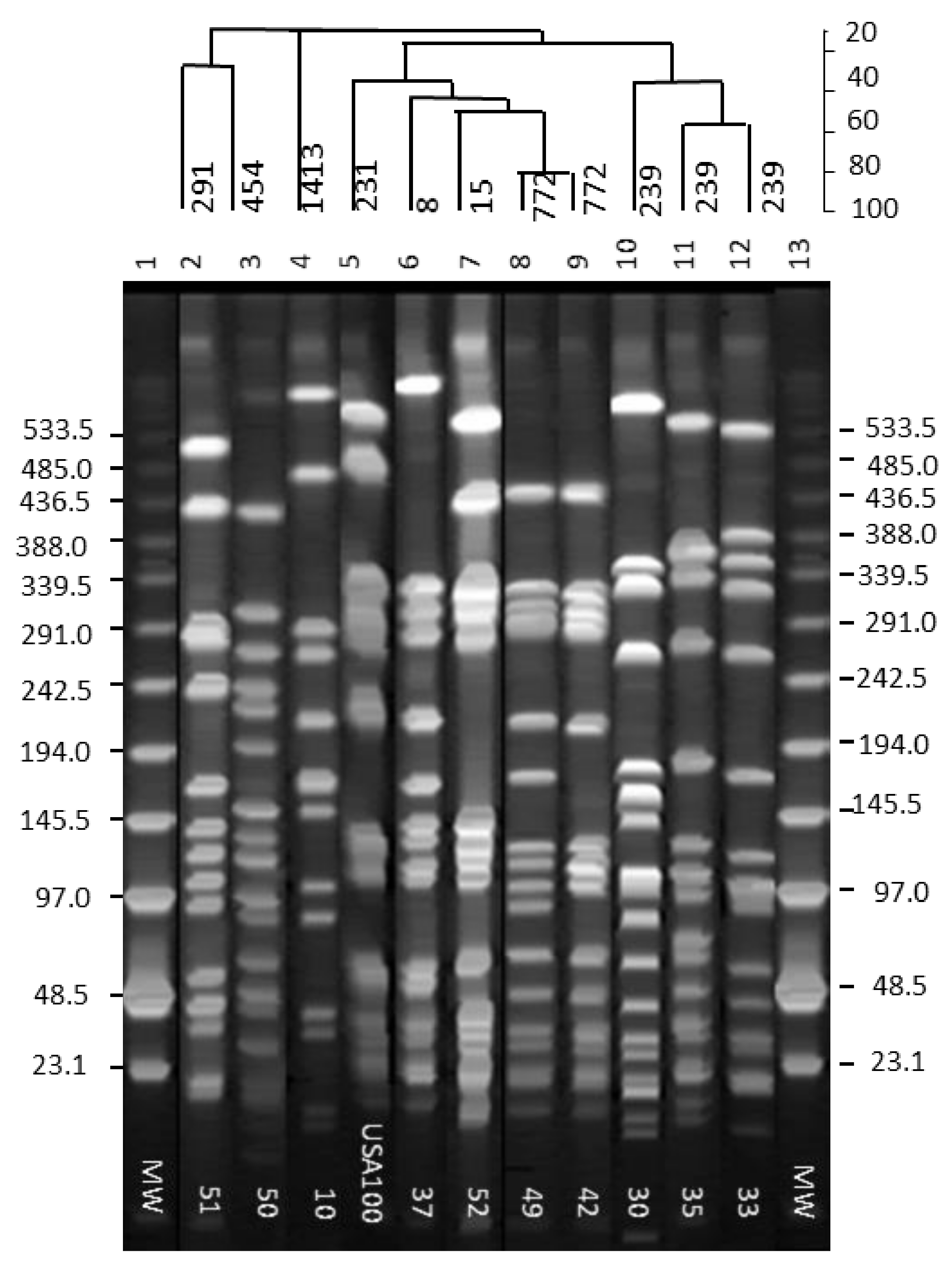
2.2. PFGE Analysis
2.3. PVL Gene Amplification and MLST and spa-Typing
2.4. SCCmec and agr Group Typing
2.5. Capsular Polysaccharides
2.6. Toxin, Adhesin, Virulence and Hemolysin Genes
3. Discussion
4. Materials and Methods
4.1. Isolation, Identification, and Characterization of Isolates
4.2. Antibiogram and MIC Using a Broth Dilution Method
4.3. DNA Plug Preparation and PFGE
4.4. DNA Isolation and Purification
4.5. Hemolysin Assay
4.6. Antimicrobial Resistance Genes
4.7. Toxin and Virulence Genes
4.8. PVL Gene Amplification, MLST and spa Typing
4.9. SCCmec and agr Group Typing
4.9.1. Determination of the Capsular Polysaccharide Type
4.9.2. Sequence Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fey, P.D.; Said-Salim, B.; Rupp, M.E.; Hinrichs, S.H.; Boxrud, D.J.; Davis, C.C.; Kreiswirth, B.N.; Schlieveret, P.M. Comparative molecular analysis of community- or hospital-acquired methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2003, 47, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Fukuta, Y.; Cunningham, C.A.; Harris, P.L.; Wagener, M.M.; Muder, R.R. Identifying the risk factors for hospital-acquired methicillin-resistant Staphylococcus aureus (MRSA) infection among patients colonized with MRSA on admission. Infect. Control Hosp. Epidemiol. 2012, 33, 1219–1225. [Google Scholar] [CrossRef]
- Boucher, H.W.; Corey, G.R. Epidemiology of methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis. 2008, 46 (Suppl. S5), S344–S449. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, K.R. “Celbenin”-resistant staphylococci. Ugeskr. Laeger. 1961, 123, 384–386. [Google Scholar] [PubMed]
- Ahmad, M.K.; Asrar, A. Prevalence of methicillin resistant Staphylococcus aureus in pyogenic community and hospital acquired skin and soft tissues infections. J. Pak. Med. Assoc. 2014, 64, 892–895. [Google Scholar]
- Taj, Y.; Abdullah, F.E.; Kazmi, S.U. Current pattern of antibiotic resistance in Staphylococcus aureus clinical isolates and the emergence of vancomycin resistance. J. Coll. Physicians Surg. Pak. 2010, 20, 728–732. [Google Scholar]
- Hussain, M.S.; Naqvi, A.; Sharaz, M. Methicillin resistant Staphylococcus aureus (MRSA); prevalence and susceptibility pattern of (MRSA) isolated from pus in tertiary care of district hospital of Rahim Yar Khan. Prof. Med. J. 2019, 26, 122–127. [Google Scholar] [CrossRef]
- Watkins, R.R.; David, M.Z.; Salata, R.A. Current concepts on the virulence mechanisms of meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2012, 61, 1179–1193. [Google Scholar] [CrossRef] [Green Version]
- Ladhani, S.; Garbash, M. Staphylococcal skin infections in children: Rational drug therapy recommendations. Paediatr. Drugs 2005, 7, 77–102. [Google Scholar] [CrossRef]
- Bernard, P. Management of common bacterial infections of the skin. Curr. Opin. Infect. Dis. 2008, 21, 122–128. [Google Scholar] [CrossRef]
- Liu, C.; Bayer, A.; Cosgrove, S.E.; Daum, R.S.; Fridkin, S.K.; Gorwitz, R.J.; Kaplan, S.L.; Karchmer, A.W.; Levine, D.P.; Murray, B.E.; et al. Clinical practice guidelines by the infectious diseases society of america for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children: Executive summary. Clin. Infect. Dis. 2011, 52, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Deresinski, S. Methicillin-resistant Staphylococcus aureus: An evolutionary, epidemiologic, and therapeutic odyssey. Clin. Infect. Dis. 2005, 40, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Waites, K.B.; Moser, S.A.; Cloud, G.A.; Hoesley, C.J. Prevalence of inducible clindamycin resistance among community- and hospital-associated Staphylococcus aureus isolates. J. Clin. Microbiol. 2006, 44, 2481–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.K.; Lyons, D.C.; Rosen, T. New and emerging concepts in managing and preventing community-associated methicillin-resistant Staphylococcus aureus infections. Int. J. Dermatol. 2015, 54, 1226–1232. [Google Scholar] [CrossRef]
- Boyle-Vavra, S.; Daum, R.S. Community-acquired methicillin-resistant Staphylococcus aureus: The role of Panton-Valentine leukocidin. Lab. Investig. 2007, 87, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Nair, G.B.; Niederman, M.S. Community-acquired pneumonia: An unfinished battle. Med. Clin. N. Am. 2011, 95, 1143–1161. [Google Scholar] [CrossRef]
- Graves, S.F.; Kobayashi, S.D.; DeLeo, F.R. Community-associated methicillin-resistant Staphylococcus aureus immune evasion and virulence. J. Mol. Med. 2010, 88, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Tristan, A.; Bes, M.; Meugnier, H.; Lina, G.; Bozdogan, B.; Courvalin, P.; Reverdy, M.E.; Enright, M.C.; Vandenesch, F.; Etienne, J. Global distribution of Panton-Valentine leukocidin--positive methicillin-resistant Staphylococcus aureus. Emerg. Infect. Dis. 2007, 13, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, J.; Kamio, Y. Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: Structures, pore-forming mechanism, and organization of the genes. Biosci. Biotechnol. Biochem. 2004, 68, 981–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, M.T.; Feil, E.J.; Lindsay, J.A.; Peacock, S.J.; Day, N.P.; Enright, M.C.; Foster, T.J.; Moore, C.E.; Hurst, L.; Atkin, R.; et al. Complete genomes of two clinical Staphylococcus aureus strains: Evidence for the rapid evolution of virulence and drug resistance. Proc. Natl. Acad. Sci. USA 2004, 101, 9786–9791. [Google Scholar] [CrossRef] [Green Version]
- Hookey, J.V.; Richardson, J.F.; Cookson, B.D. Molecular typing of Staphylococcus aureus based on PCR restriction fragment length polymorphism and DNA sequence analysis of the coagulase gene. J. Clin. Microbiol. 1998, 36, 1083–1089. [Google Scholar] [CrossRef] [Green Version]
- Booth, M.C.; Hatter, K.L.; Miller, D.; Davis, J.; Kowalski, R.; Parke, D.W.; Chodosh, J.; Jett, B.D.; Callegan, M.C.; Penland, R.; et al. Molecular epidemiology of Staphylococcus aureus and Enterococcus faecalis in endophthalmitis. Infect. Immun. 1998, 66, 356–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowrouzian, F.L.; Karami, N.; Welinder-Olsson, C.; Ahren, C. Virulence gene typing of methicillin-resistant Staphylococcus aureus as a complement in epidemiological typing. J. Microbiol. Methods 2013, 93, 173–176. [Google Scholar] [CrossRef]
- Naimi, T.S.; LeDell, K.H.; Como-Sabetti, K.; Borchardt, S.M.; Boxrud, D.J.; Etienne, J.; Johnson, S.K.; Vandenesch, F.; Fridkin, S.; O’Boyle, C.; et al. Comparison of community- and health care-associated methicillin-resistant Staphylococcus aureus infection. JAMA 2003, 290, 2976–2984. [Google Scholar] [CrossRef] [Green Version]
- Calfee, D.P. The epidemiology, treatment, and prevention of transmission of methicillin-resistant Staphylococcus aureus. J. Infus. Nurs. 2011, 34, 359–364. [Google Scholar] [CrossRef]
- Jimenez, J.N.; Ocampo, A.M.; Vanegas, J.M.; Rodriguez, E.A.; Mediavilla, J.R.; Chen, L.; Muskus, C.E.; Vélez, L.A.; Rojas, C.; Restrepo, A.V.; et al. CC8 MRSA strains harboring SCCmec type IVc are predominant in Colombian hospitals. PLoS ONE 2012, 7, e38576. [Google Scholar]
- Teixeira, M.M.; Araujo, M.C.; Silva-Carvalho, M.C.; Beltrame, C.O.; Oliveira, C.C.H.B.; Figueiredo, A.M.S.; Oliveira, A.G. Emergence of clonal complex 5 (CC5) methicillin-resistant Staphylococcus aureus (MRSA) isolates susceptible to trimethoprim-sulfamethoxazole in a Brazilian hospital. Braz. J. Med. Biol. Res. 2012, 45, 637–643. [Google Scholar] [CrossRef]
- Deurenberg, R.H.; Stobberingh, E.E. The molecular evolution of hospital- and community-associated methicillin-resistant Staphylococcus aureus. Curr. Mol. Med. 2009, 9, 100–115. [Google Scholar] [CrossRef]
- Madzgalla, S.; Syed, M.A.; Khan, M.A.; Rehman, S.S.; Muller, E.; Reissig, A.; Ehricht, R.S.; Monecke, S. Molecular characterization of Staphylococcus aureus isolates causing skin and soft tissue infections in patients from Malakand, Pakistan. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1541–1547. [Google Scholar] [CrossRef]
- Zafar, A.; Stone, M.; Ibrahim, S.; Parveen, Z.; Hasan, Z.; Khan, E.; Hasan, R.; Wain, J.; Bamford, K. Prevalent genotypes of meticillin-resistant Staphylococcus aureus: Report from Pakistan. J. Med. Microbiol. 2011, 60, 56–62. [Google Scholar] [CrossRef]
- Shabir, S.; Hardy, K.J.; Abbasi, W.S.; McMurray, C.L.; Malik, S.A.; Wattal, C.; Hawkey, P.M. Epidemiological typing of meticillin-resistant Staphylococcus aureus isolates from Pakistan and India. J. Med. Microbiol. 2010, 59, 330–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Takano, T.; Higuchi, W.; Iwao, Y.; Singur, O.; Reva, I.; Otsuka, Y.; Nakayashiki, T.; Mori, H.; Reva, G.; et al. Comparative genomics and drug resistance of a geographic variant of ST239 methicillin-resistant Staphylococcus aureus emerged in Russia. PLoS ONE 2012, 7, e29187. [Google Scholar] [CrossRef]
- Shore, A.; Rossney, A.S.; Keane, C.T.; Enright, M.C.; Coleman, D.C. Seven novel variants of the staphylococcal chromosomal cassette mec in methicillin-resistant Staphylococcus aureus isolates from Ireland. Antimicrob. Agents Chemother. 2005, 49, 2070–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome E. Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar] [CrossRef] [Green Version]
- Chongtrakool, P.; Ito, T.; Ma, X.X.; Kondo, Y.; Trakulsomboon, S.; Tiensasitorn, C.; Jamklang, M.; Chavalit, T.; Song, J.H.; Hiramatsu, K. Staphylococcal cassette chromosome mec (SCCmec) typing of methicillin-resistant Staphylococcus aureus strains isolated in 11 Asian countries: A proposal for a new nomenclature for SCCmec elements. Antimicrob. Agents Chemother. 2006, 50, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Palavecino, E. Community-acquired methicillin-resistant Staphylococcus aureus infections. Clin. Lab. Med. 2004, 24, 403–418. [Google Scholar] [CrossRef]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef]
- Asghar, A.H. Molecular characterization of methicillin-resistant Staphylococcus aureus isolated from tertiary care hospitals. Pak. J. Med. Sci. 2014, 30, 698–702. [Google Scholar] [CrossRef]
- Campbell, S.J.; Deshmukh, H.S.; Nelson, C.L.; Bae, I.G.; Stryjewski, M.E.; Federspiel, J.J.; Tonthat, G.T.; Rude, T.H.; Barriere, S.L.; Corey, R.; et al. Genotypic characteristics of Staphylococcus aureus isolates from a multinational trial of complicated skin and skin structure infections. J. Clin. Microbiol. 2008, 46, 678–684. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Stevens, D.L.; Hamilton, S.M.; Parimon, T.; Ma, Y.; Kearns, A.M.; Ellis, R.W.; Bryant, A.E. Fatal S. aureus hemorrhagic pneumonia: Genetic analysis of a unique clinical isolate producing both PVL and TSST-1. PLoS ONE 2011, 6, e27246. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Mongodin, E.; Bajolet, O.; Cutrona, J.; Bonnet, N.; Dupuit, F.; Puchelle, E.; Bentzmann, S. Fibronectin-binding proteins of Staphylococcus aureus are involved in adherence to human airway epithelium. Infect. Immun. 2002, 70, 620–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salasia, S.I.; Tato, S.; Sugiyono, N.; Ariyanti, D.; Prabawati, F. Genotypic characterization of Staphylococcus aureus isolated from bovines, humans, and food in Indonesia. J. Vet. Sci. 2011, 12, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Ferry, T.; Thomas, D.; Genestier, A.L.; Bes, M.; Lina, G.; Vandenesch, F.; Etienne, J. Comparative prevalence of superantigen genes in Staphylococcus aureus isolates causing sepsis with and without septic shock. Clin. Infect. Dis. 2005, 41, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Jarraud, S.; Cozon, G.; Vandenesch, F.; Bes, M.; Etienne, J.; Lina, G. Involvement of enterotoxins G and I in staphylococcal toxic shock syndrome and staphylococcal scarlet fever. J. Clin. Microbiol. 1999, 37, 2446–2449. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Friedrich, A.W.; Lubritz, G.; Weilert, M.; Peters, G.; Von Eiff, C. Prevalence of genes encoding pyrogenic toxin superantigens and exfoliative toxins among strains of Staphylococcus aureus isolated from blood and nasal specimens. J. Clin. Microbiol. 2003, 41, 1434–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimian, A.; Najar-Pirayeh, S.; Mirab-Samiee, S.; Naderi, M. Occurrence of methicillin resistant Staphylococcus aureus (MRSA) among clinical samples in tehran-iran and its correlation with polymorphism of specific accessory gene regulator (AGR) groups. Braz. J. Microbiol. 2012, 43, 779–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2009. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2009. [Google Scholar]
- Marasa, B.S.; Iram, S.; Sung, K.; Kweon, O.G.; Cerniglia, C.E.; Khan, S. Molecular characterization of fluoroquinolone resistance of methicillin-resistant clinical Staphylococcus aureus isolates from Rawalpindi, Pakistan. Med. Res. Arch. 2015, 2, 1–14. [Google Scholar]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: Establishing a national database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef] [Green Version]
- Lattar, S.M.; Tuchscherr, L.P.; Caccuri, R.L.; Centron, D.; Becker, K.; Alonso, C.A.; Barberis, C.; Miranda, G.; Buzzola, F.R.; Eiff, C.V.; et al. Capsule expression and genotypic differences among Staphylococcus aureus isolates from patients with chronic or acute osteomyelitis. Infect. Immun. 2009, 77, 1968–1975. [Google Scholar] [CrossRef] [Green Version]
- Haveri, M.; Roslof, A.; Rantala, L.; Pyorala, S. Virulence genes of bovine Staphylococcus aureus from persistent and nonpersistent intramammary infections with different clinical characteristics. J. Appl. Microbiol. 2007, 103, 993–1000. [Google Scholar] [CrossRef]
- Fomda, B.A.; Thokar, M.A.; Bashir, G.; Khan, A.; Kour, A.; Zahoor, D.; Ray, P. Prevalence and genotypic relatedness of methicillin resistant Staphylococcus aureus in a tertiary care hospital. J. Postgrad. Med. 2014, 60, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, F.J.; Fluit, A.C.; Gondolf, M.; Beyrau, R.; Lindenlauf, E.; Verhoef, J.; Ray, P. The prevalence of aminoglycoside resistance and corresponding resistance genes in clinical isolates of staphylococci from 19 European hospitals. J. Antimicrob. Chemother. 1999, 43, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, J.E.; Christensen, H.; Aarestrup, F.M. Diversity and evolution of blaZ from Staphylococcus aureus and coagulase-negative staphylococci. J. Antimicrob. Chemother. 2006, 57, 450–460. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Sadurski, R.; Kray, A.; Boos, M.; Geisel, R.; Kohrer, K.; Jan Verhoefband, J.; Fluit, A.C. Prevalence of macrolide-resistance genes in Staphylococcus aureus and Enterococcus faecium isolates from 24 European university hospitals. J. Antimicrob. Chemother. 2000, 45, 891–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, K.T.; Hanifah, Y.A.; Yusof, M.; Thong, K.L. ermA, ermC, tetM and tetK are essential for erythromycin and tetracycline resistance among methicillin-resistant Staphylococcus aureus strains isolated from a tertiary hospital in Malaysia. Indian J. Med. Microbiol. 2012, 30, 203–207. [Google Scholar] [CrossRef]
- Kadlec, K.; Fessler, A.T.; Hauschild, T.; Schwarz, S. Novel and uncommon antimicrobial resistance genes in livestock-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, L.; Ip, M.; Sun, M.; Sun, J.; Huang, G.; Wang, C.; Deng, L.; Zheng, Y.; Fu, Z.; et al. Molecular and clinical characteristics of clonal complex 59 methicillin-resistant Staphylococcus aureus infections in Mainland China. PLoS ONE 2013, 8, e70602. [Google Scholar] [CrossRef] [Green Version]
- Orii, K.O.; Iwao, Y.; Higuchi, W.; Takano, T.; Yamamoto, T. Molecular characterization of methicillin-resistant Staphylococcus aureus from a fatal case of necrotizing fasciitis in an extremely low-birth-weight infant. Clin. Microbiol. Infect. 2010, 16, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Saunders, N.A.; Holmes, A. Multilocus sequence typing (MLST) of Staphylococcus aureus. Methods Mol. Biol. 2014, 1085, 113–130. [Google Scholar]
- Heym, B.; Le Moal, M.; Armand-Lefevre, L.; Nicolas-Chanoine, M.H. Multilocus sequence typing (MLST) shows that the ‘Iberian’ clone of methicillin-resistant Staphylococcus aureus has spread to France and acquired reduced susceptibility to teicoplanin. J. Antimicrob. Chemother. 2002, 50, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Koreen, L.; Ramaswamy, S.V.; Graviss, E.A.; Naidich, S.; Musser, J.M.; Kreiswirth, B.N. Spa typing method for discriminating among Staphylococcus aureus isolates: Implications for use of a single marker to detect genetic micro- and macrovariation. J. Clin. Microbiol. 2004, 42, 792–799. [Google Scholar] [CrossRef] [Green Version]
- Hallin, M.; Friedrich, A.W.; Struelens, M.J. Spa typing for epidemiological surveillance of Staphylococcus aureus. Methods Mol. Biol. 2009, 551, 189–202. [Google Scholar]
- Votintseva, A.A.; Fung, R.; Miller, R.R.; Knox, K.; Godwin, H.; Wyllie, D.H.; Bowden, R.; Crook, D.W.; Walke, A.S. Prevalence of Staphylococcus aureus protein A (spa) mutants in the community and hospitals in Oxfordshire. BMC Microbiol. 2014, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Hata, E.; Katsuda, K.; Kobayashi, H.; Uchida, I.; Tanaka, K.; Eguchi, M. Genetic variation among Staphylococcus aureus strains from bovine milk and their relevance to methicillin-resistant isolates from humans. J. Clin. Microbiol. 2010, 48, 2130–2139. [Google Scholar] [CrossRef] [Green Version]
- Peacock, S.J.; Moore, C.E.; Justice, A.; Kantzanou, M.; Story, L.; Mackie, K.; O’Neill, G.; Day, N.P.J. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect Immun. 2002, 70, 4987–4996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdier, I.; Durand, G.; Bes, M.; Taylor, K.L.; Lina, G.; Vandenesch, F.; Fattom, A.I.; Etienne, J. Identification of the capsular polysaccharides in Staphylococcus aureus clinical isolates by PCR and agglutination tests. J. Clin. Microbiol. 2007, 45, 725–729. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isolate No. | Aminoglycoside | Chloramphenicol | Erythromycin | Tetracycline | Trimethoprim | Methicillin | Beta-Lactam |
|---|---|---|---|---|---|---|---|
| 10 | [aac(6′)-Ie-aph(2′)-Ia, str] [aac(6′)-Ie-aph(2)-Ia, str] | [catpC221 and catpC223] | [ermB, sat4] | [tetL tetM, tetS, tetW] | [dfrA] | [mecA] | [ND] |
| 15 | [aac(6′)-aph(2″), | [cat(pC194), cat(pC221), catpC223] | [ND] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [ND] |
| 25 | [aac(6′)-aph(2″), aph(3′)-IIIa, aac(6′)-Ie-aph(2’)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [sat4] | [tetL, tetM, tetS, tetW] | [dfrA] | [mecA] | [ND] |
| 30 | [aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia] | [cat(pC194), cat(pC221), catpC223] | [sat4] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [ND] |
| 31 | [aac(6′)-aph(2″), aac(6′)-Ie-aph(2′)-Ia] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| [aac(6′)-aph(2″), aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] | |
| 37 | [aac(6′)-aph(2″), aph(3′)-IIIa | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [ND] |
| 40 | [aac(6′)-aph(2″), ant(4′)-Ia, aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 42 | [aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 49 | [aac(6′)-aph(2″), ant(4′)-Ia, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 32 | [aac(6′)-aph(2″), ant(4′)-Ia, aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetK, tetL, tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 33 | [aac(6′)-aph(2″), aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetK, tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 34 | [aac(6′)-aph(2″), aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 38 | [aac(6′)-aph(2″), aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 41 | [aac(6′)-aph(2″), aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 48 | [aac(6′)-aph(2″), aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB] | [tetM, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 50 | [aac(6′)-Ie-aph(2′)-Ia, str] | [ND] | [sat4] | [tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 51 | [ant(4′)-Ia, and aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia, str] | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetK, tetS, tetW] | [dfrA] | [mecA] | [blaZ] |
| 52 | [ant(4′)-Ia, and aph(3′)-IIIa, aac(6′)-Ie-aph(2′)-Ia] | [cat(pC194), cat(pC221), catpC223] | [ermB, sat4] | [tetS, tetW] | [dfrA] | [mecA] | blaZ] |
| PFGE Profile | Isolate No | Origin | Infection Type | Symptoms/Reasons Admitted for | MLST Type | spa Type | SCCmec Type | agr Group | Capsular Genotype |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 32 | P | HA | DNR Colitis Tracheostomy | ST239 | t030 | III | I | 8 |
| 1 | 33 * | P | CA | Septic Shock | ST239 | t030 | III | I | 8 |
| 1 | 34 | P | CA | Intestinal Obstruction | ST239 | t030 | III | I | NT |
| 1 | 41 | P | HA | Anxiolytic Poisoning | ST239 | t030 | III | I | 8 |
| 1 | 48 | P | CA | SOB, Fever | ST239 | t030 | III | I | 8 |
| 1 | 38 | P | HA | CuSo4 Poisoning | ST8 | New | V | I | 5 |
| 2 | 15 | N | HA | Hepato-Splenomegaly | ST239 | New | III | I | 8 |
| 2 | 25 * | N | CA | Chronic Renal Failure | ST8 | t064 | IVa | I | 5 |
| 2 | 31 * | N | HA | Tetanus, Locked Jaw | ST239 | t030 | III, IVa | I | 8 |
| 2 | 37 | N | HA | Cardiogenic shock | ST8 | t064 | IVa | I | 5 |
| 2 | 40 * | N | HA | Kyphoscoliosis | ST30 | t138 | IVa | I | 8 |
| 3 | 10 * | N | HA | Guillain Barr Syndrome | ST1413 | t314 | III, V | I | 8 |
| 4 | 30 | N | HA | Acute unconsciousness | ST239 | t987 | III | I | 8 |
| 5 | 35 | N | HA | Dilated Cardiomyopathy | ST239 | t030 | III | I | 8 |
| 6 | 42 | N | CA | Hepato-Splenomegaly | ST772 | t5414 | III | I | 5 |
| 7 | 49 | N | CA | Intestinal Obstruction | ST772 | t5414 | III | I | 5 |
| 8 | 50 | P | HA | CuSo4 Poisoning | ST503 | t138 | III, IVa, V | I | 5 |
| 9 | 51 * | N | HA | Chronic Renal Failure | ST291 | t1149 | III, V | I | NT |
| 10 | 52 | P | HA | Neuropathy, Tuberculosis | ST15 | t1509 | III, IVa, V | I | 5 |
| X-Region of the Staphylococcal Protein A Gene | spa Type | Isolate # | |
|---|---|---|---|
| 5′ FL-W1 G1 K1 A1 Q1 K1 A1 O1 M1 Q1-3 FL′ | t987 | 30 | |
| 5′ FL-Y1 H1 G1 C1 M1 B1 Q1 B1 L1 O1 AAGAAGATGGTAACGGAGTACATG-3′ FL | t064 | 25, 37 | |
| 5′ FL-T1 K1 J1 E1 F1 M1 B1 P1 B1-3′ FL | t5414 | 42, 49 | |
| 5′ FL-W1 G1 K1 A1 Q1 Q1-3′ FL | t030 | 31-35, 41, 48 | |
| 5′ FL-X1 K1 A1 O1 M1 Q1-3′ FL | t138 | 40, 50 | |
| 5′ FL-X1 M1 J1 H2 M1-3′ FL | t314 | 10 | |
| 5′ FL-U1 J1 G1 J1-3′ FL | t1509 | 52 | |
| 5′ FL-Y1 H1 G1 C1 M1 B1 Q1 B1 L1 AAATAAGATGGTAACTGATTACATG-3′ FL | New | 38 | |
| 5′- CCCAAGACAGCAACAAGCCTGGTA AGAGAGGACGGCAACAAACCTGGT- | New | 15 | |
| AAAGAAGACAGCAAAAAAACTGGCAAGACGATGGCAACAAGCCGGGCA--3′ FL | |||
| 5′- TGNAAGACGGCAACAAACCTGGTA AAAGAAGACAACGAAAAACCTGGT | New | 51 | |
| AAAGAAGATGGCAACTAGCCTGGT B1 B1 AAAGAAGACGGCTACAAGCCTGGT--3′ FL | |||
| A1: AAAGAAGACAACAAAAAACCTGGC | B1: AAAGAAGACAACAAAAAACCTGGT | C1: AAAGAAGACAACAAAAAGCCTGGC | |
| E1: AAAGAAGACGGCAACAAACCTGGC | F1: AAAGAAGACAACAACAAGCCTGGC | G1: AAAGAAGACAACAACAAGCCTGGT | |
| H1: AAAGAAGACAATAACAAGCCTGGC | H2: AAAGAAGATGGCAACAAGCCTAGT | J1: AAAGAAGACGGCAACAAACCTGGT | |
| K1: AAAGAAGACGGCAACAAACCTGGT | L1: AAAGAAGACGGCAACAAGCCTGGC | M1: AAAGAAGACGGCAACAAGCCTGGT | |
| O1: AAAGAAGATGGCAACAAACCTGGT | P1: AAAGAAGATGGCAACAAGCCTGGC | Q1: AAAGAAGATGGCAACAAGCCTGGT | |
| T1: GAGGAAGACAACAAAAAACCTGGT | X1: GAGGAAGACAACAACAAGCCTGGT | Y1: GAGGAAGACAATAACAAGCCTGGC | |
| W1: GAGGAAGACAACAACAAGCCTGGC | 5′-FL: TAAACGATGCTCAAGCACCAAAAG | 3′-FL: AAGAAGACGGCAACGGAGTACATG | |
| Isolate No. | Toxin Genes | Adhesin Genes | Other Virulence Genes | Hemolysin Activity |
|---|---|---|---|---|
| 10 | [eta, sea, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (ica, cfb, v8) | γ |
| 15 | [seb, sec, hla, hlb, hld, hlg, pvl (lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (cfb, v8) | α |
| 25 | [sea, seb, sec, hla, hlb, hld, hlg, pvl (lukE-D), sed, see, seg, seh, tst | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (ica, cfb, v8) | α |
| 30 | [eta, sea, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | γ |
| 31 | [eta, seb, sec, sei, hla, hlb, hld, hlg, pvl (lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (ica, cfb, v8) | γ |
| 35 | [eta, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | γ |
| 37 | [eta, sea, seb, sec, sej, hla, hlb, hld, hlg, pvl (lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | α |
| 40 | [eta, sea, seb, sec, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | β |
| 42 | [eta, seb, sec, sei, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | α |
| 49 | [eta, sea, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | β |
| 32 | [eta, sea, seb, sec, sei, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | γ |
| 33 | [eta, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | γ |
| 34 | [eta, sec, sei, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | γ |
| 38 | [eta, sea, seb, sec, sei, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | β |
| 41 | [eta, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | α |
| 48 | [eta, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | α |
| 50 | [sea, seb, sec, sei, sej, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | β |
| 51 | [eta, sea, seb, sec, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | β |
| 52 | [eta, sea, seb, sec, hla, hlb, hld, hlg, pvl (lukS, lukF, lukM, lukE-D), sed, see, seg, seh, tst] | (ebpS, bbp, can, clfA, clfB, fnbA, fnbB, map-eap, spa) | (chp, ica, cfb, v8) | α |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.; Marasa, B.S.; Sung, K.; Nawaz, M. Genotypic Characterization of Clinical Isolates of Staphylococcus aureus from Pakistan. Pathogens 2021, 10, 918. https://doi.org/10.3390/pathogens10080918
Khan S, Marasa BS, Sung K, Nawaz M. Genotypic Characterization of Clinical Isolates of Staphylococcus aureus from Pakistan. Pathogens. 2021; 10(8):918. https://doi.org/10.3390/pathogens10080918
Chicago/Turabian StyleKhan, Saeed, Bernard S. Marasa, Kidon Sung, and Mohamed Nawaz. 2021. "Genotypic Characterization of Clinical Isolates of Staphylococcus aureus from Pakistan" Pathogens 10, no. 8: 918. https://doi.org/10.3390/pathogens10080918
APA StyleKhan, S., Marasa, B. S., Sung, K., & Nawaz, M. (2021). Genotypic Characterization of Clinical Isolates of Staphylococcus aureus from Pakistan. Pathogens, 10(8), 918. https://doi.org/10.3390/pathogens10080918