
Identification of Toxocara canis Antigen-Interacting Partners by Yeast Two-Hybrid Assay and a Putative Mechanism of These Host–Parasite Interactions



Abstract
:1. Introduction
2. Results
2.1. Construction of Bait Plasmids
2.2. Autoactivation and Toxicity Test
2.3. Yeast Two-Hybrid Screen
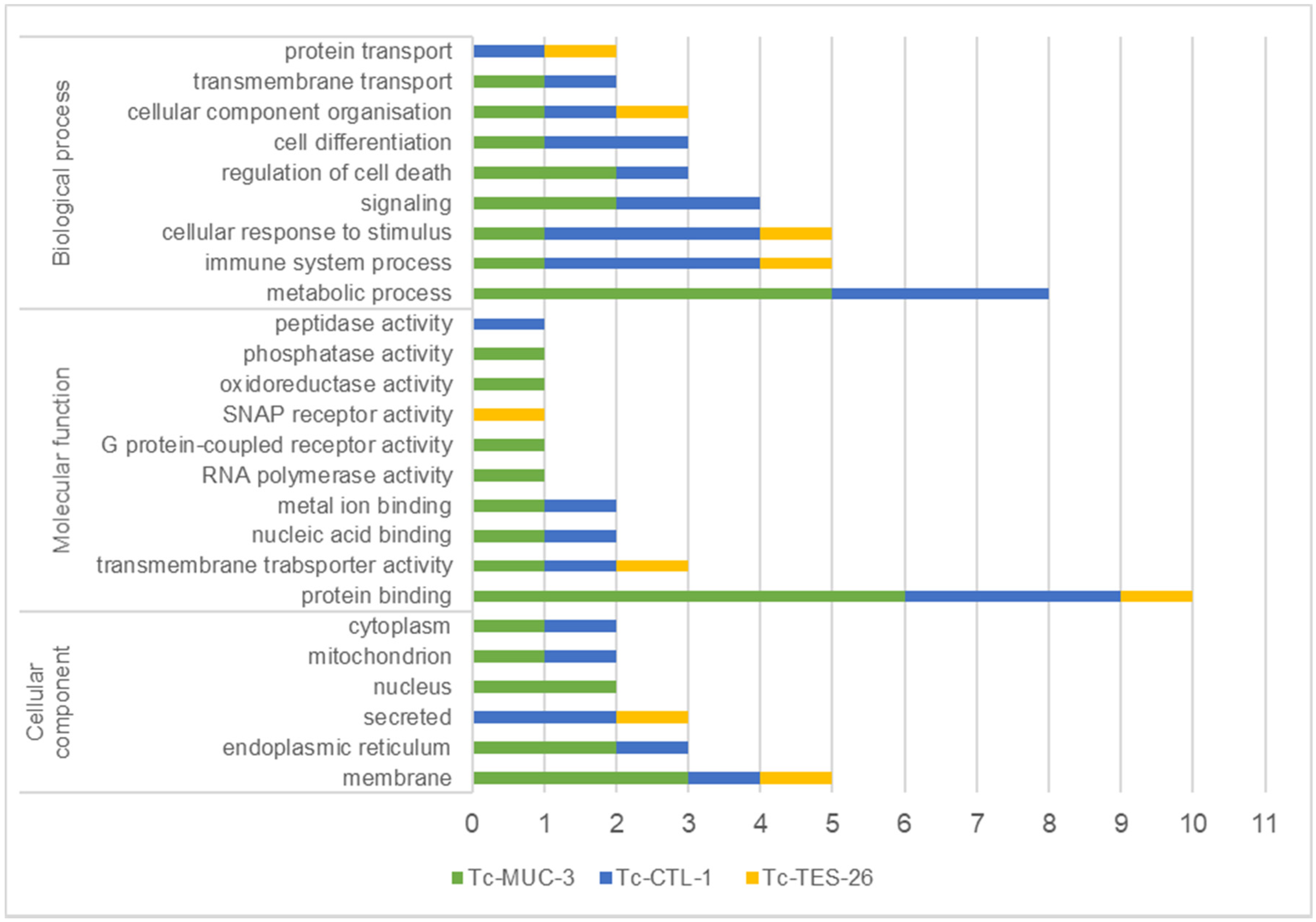
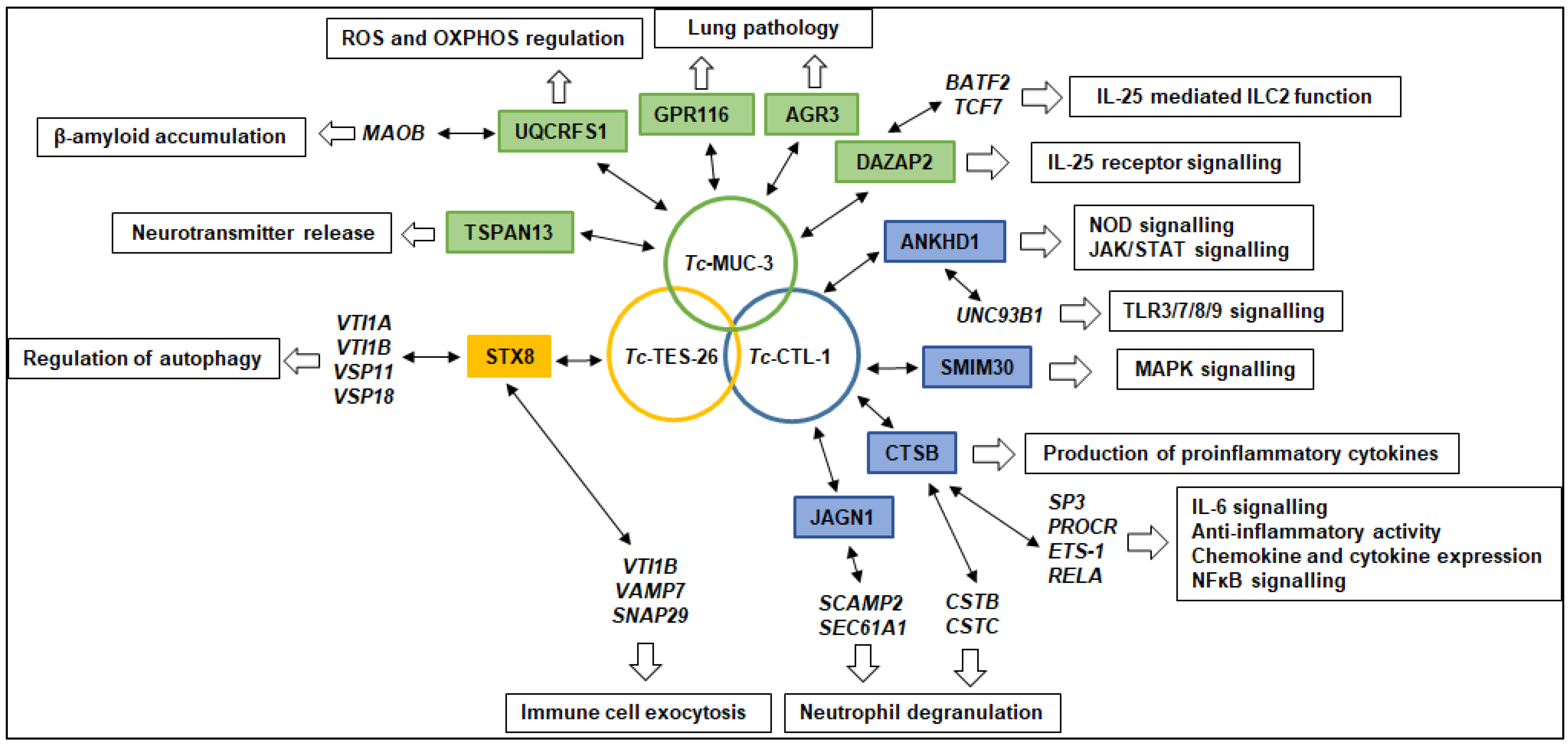
2.4. Identification of Interacting Proteins
3. Discussion
4. Materials and Methods
4.1. Bait Plasmid Construction
4.2. Yeast Transformation, Autoactivation and Toxicity Test
4.3. Yeast Two-Hybrid Screen
4.4. Positive Prey Analysis by Bioinformatics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rostami, A.; Riahi, S.M.; Holland, C.V.; Taghipour, A.; Khalili-Fomeshi, M.; Fakhri, Y.; Omrani, V.F.; Hotez, P.J.; Gasser, R.B. Seroprevalence estimates for toxocariasis in people worldwide: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2019, 13, e0007809. [Google Scholar] [CrossRef]
- Skulinova, K.; Novak, J.; Kasny, M.; Kolarova, L. Seroprevalence of Larval Toxocarosis in the Czech Republic. Acta Parasitol. 2019, 65, 68–76. [Google Scholar] [CrossRef]
- Jarosz, W.; Mizgajska-Wiktor, H.; Kirwan, P.; Konarski, J.; Rychlicki, W.; Wawrzyniak, G. Developmental age, physical fit-ness and Toxocara seroprevalence amongst lower-secondary students living in rural areas contaminated with Toxocara eggs. Parasitology 2010, 137, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.K.; Holland, C.V.; Loxton, K.; Barghouth, U. Cerebral toxocariasis: Silent progression to neurodegenerative disor-ders? Clin. Microbiol. Rev. 2015, 28, 663–686. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Holland, C.V.; Wang, T.; Hofmann, A.; Fan, C.K.; Maizels, R.M.; Hotez, P.J.; Gasser, R.B. Human toxocariasis. Lancet Infect. Dis. 2018, 18, e14–e24. [Google Scholar] [CrossRef]
- Magnaval, J.-F.; Glickman, L.T.; Dorchies, P.; Morassin, B. Highlights of human toxocariasis. Korean J. Parasitol. 2001, 39, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinelli, E.; Brandes, S.; Dormans, J.; Gremmer, E.; Van Loveren, H. Infection with the roundworm Toxocara canis leads to ex-acerbation of experimental allergic airway inflammation. Clin. Exp. Allergy 2008, 38, 649–658. [Google Scholar] [CrossRef]
- Aghaei, S.; Riahi, S.M.; Rostami, A.; Mohammadzadeh, I.; Javanian, M.; Tohidi, E.; Foroutan, M.; Dooki, M.E. Toxocara spp. infection and risk of childhood asthma: A systematic review and meta-analysis. Acta Trop. 2018, 182, 298–304. [Google Scholar] [CrossRef]
- Zhu, X.-Q.; Korhonen, P.K.; Cai, H.; Young, N.; Nejsum, P.; Von Samson-Himmelstjerna, G.; Boag, P.; Tan, P.; Li, Q.; Min, J.; et al. Genetic blueprint of the zoonotic pathogen Toxocara canis. Nat. Commun. 2015, 6, 6145. [Google Scholar] [CrossRef] [Green Version]
- Caby, S.; Pagliazzo, L.; Lancelot, J.; Saliou, J.-M.; Bertheaume, N.; Pierce, R.J.; Roger, E. Analysis of the interactome of Schistosoma mansoni histone deacetylase 8. PLoS Negl. Trop. Dis. 2017, 11, e0006089. [Google Scholar] [CrossRef] [Green Version]
- Luo, T.; McBride, J.W. Ehrlichia chaffeensis TRP32 Interacts with Host Cell Targets That Influence Intracellular Survival. Infect. Immun. 2012, 80, 2297–2306. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fang, R.; Yuan, Y.; Hu, M.; Zhou, Y.; Zhao, J. Identification of host proteins interacting with the integrin-like A domain of Toxoplasma gondii micronemal protein MIC2 by yeast-two-hybrid screening. Parasites Vectors 2014, 7, 543. [Google Scholar] [CrossRef]
- Loukas, A.; Hintz, M.; Linder, D.; Mullin, N.P.; Parkinson, J.; Tetteh, K.K.A.; Maizels, R.M. A family of secreted mucins from the par-asitic nematode Toxocara canis bears diverse mucin domains but shares similar flanking six-cysteine repeat motifs. J. Biol. Chem. 2000, 275, 39600–39607. [Google Scholar] [CrossRef] [Green Version]
- Loukas, A.; Mullin, N.P.; Tetteh, K.K.A.; Moens, L.; Maizels, R.M. A novel C-type lectin secreted by a tissue-dwelling para-sitic nematode. Curr. Biol. 1999, 9, 825–828. [Google Scholar] [CrossRef] [Green Version]
- Loukas, A.; Maizels, R. Helminth C-type Lectins and Host–Parasite Interactions. Parasitol. Today 2000, 16, 333–339. [Google Scholar] [CrossRef]
- Gems, D.; Ferguson, C.J.; Robertson, B.D.; Nieves, R.; Page, A.P.; Blaxter, M.; Maizels, R.M. An Abundant, trans-spliced mRNA from Toxocara canis Infective Larvae Encodes a 26-kDa Protein with Homology to Phosphatidylethanolamine-binding Proteins. J. Biol. Chem. 1995, 270, 18517–18522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maizels, R.M.; Tetteh, K.K.; Loukas, A. Toxocara canis: Genes expressed by the arrested infective larval stage of a parasitic nematode. Int. J. Parasitol. 2000, 30, 495–508. [Google Scholar] [CrossRef]
- Popova, A.; Kzhyshkowska, J.; Nurgazieva, D.; Goerdt, S.; Gratchev, A. Smurf2 regulates IL17RB by proteasomal degrada-tion of its novel binding partner DAZAP2. Immunobiology 2012, 217, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Zepp, J.A.; Wu, L.; Qian, W.; Ouyang, W.; Aronica, M.; Erzurum, S.; Li, X. TRAF4-SMURF2-mediated DAZAP2 degradation is critical for IL-25 signaling and allergic airway inflammation. J. Immunol. 2015, 194, 2826–2837. [Google Scholar] [CrossRef] [Green Version]
- Angkasekwinai, P.; Park, H.; Wang, Y.H.; Wang, Y.H.; Seon, H.C.; Corry, D.B.; Liu, Y.J.; Zhu, Z.; Dong, C. Interleukin 25 promotes the initiation of proallergic type 2 responses. J. Exp. Med. 2007, 204, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inuo, G.; Akao, N.; Ohsaka, H.K.; Saito, I.; Miyasaka, N.; Fujita, K. Toxocara canis adult worm antigen induces proliferative response of healthy human peripheral blood mononuclear cells. Parasite Immunol. 1995, 17, 77–84. [Google Scholar] [CrossRef]
- Faz-López, B.; Ledesma-Soto, Y.; Romero-Sánchez, Y.; Calleja, E.; Martínez-Labat, P.; Terrazas, L.I. Signal Transducer and Activator of Transcription Factor 6 Signaling Contributes to Control Host Lung Pathology but Favors Susceptibility against Toxocara canis Infection. BioMed Res. Int. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hanh, N.T.L.; Lee, Y.L.; Lin, C.L.; Chou, C.M.; Cheng, P.C.; Quang, H.H.; Fan, C.K. Evidence for asthma in the lungs of mice in-oculated with different doses of Toxocara canis. Am. J. Trop. Med. Hyg. 2020, 103, 2305–2314. [Google Scholar] [CrossRef]
- Długosz, E.; Wasyl, K.; Klockiewicz, M.; Wiśniewski, M. Toxocara canis mucins among other excretory-secretory antigens induce in vitro secretion of cytokines by mouse splenocytes. Parasitol. Res. 2015, 114, 3365–3371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleul, C.C.; Fuhlbrigge, R.C.; Casasnovas, J.M.; Aiuti, A.; Springer, T.A. A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1). J. Exp. Med. 1996, 184, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Bao, K.; Carr, T.; Wu, J.; Barclay, W.; Jin, J.; Ciofani, M.; Reinhardt, R.L. BATF Modulates the Th2 Locus Control Region and Regulates CD4+ T Cell Fate during Antihelminth Immunity. J. Immunol. 2016, 197, 4371–4381. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.M.; Patel, P.S.; Bao, K.; Danhorn, T.; O’Connor, B.P.; Reinhardt, R.L. BATF acts as an essential regulator of IL-25–responsive migratory ILC2 cell fate and function. Sci. Immunol. 2020, 5, eaay3994. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, W.; Wang, X. Roles of transcriptional factor 7 in production of inflammatory factors for lung diseases. J. Transl. Med. 2015, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Thiébaut, R.; Esmiol, S.; Lecine, P.; Mahfouz, B.; Hermant, A.; Nicoletti, C.; Parnis, S.; Perroy, J.; Borg, J.-P.; Pascoe, L.; et al. Characterization and Genetic Analyses of New Genes Coding for NOD2 Interacting Proteins. PLoS ONE 2016, 11, e0165420. [Google Scholar] [CrossRef] [PubMed]
- Fisher, K.H.; Fragiadaki, M.; Pugazhendhi, D.; Bausek, N.; Arredondo, M.A.; Thomas, S.J.; Brown, S.; Zeidler, M.P. A genome-wide RNAi screen identifies MASK as a positive regulator of cytokine receptor stability. J. Cell Sci. 2018, 131, jcs209551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnardi, I.; Ng, Y.-S.; Srdanovic, I.; Motaghedi, R.; Rudchenko, S.; Von Bernuth, H.; Zhang, S.-Y.; Puel, A.; Jouanguy, E.; Picard, C.; et al. IRAK-4- and MyD88-Dependent Pathways Are Essential for the Removal of Developing Autoreactive B Cells in Humans. Immunity 2008, 29, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Liu, Z.; Han, H.; Wang, B.; Li, W.; Mao, C.; Liu, S. Peptide SMIM30 promotes HCC development by inducing SRC/YES1 membrane anchoring and MAPK pathway activation. J. Hepatol. 2020, 73, 1155–1169. [Google Scholar] [CrossRef]
- Michel, V.; Bakovic, M. The ubiquitous choline transporter SLC44A1. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Wille, S.; Szekeres, A.; Majdic, O.; Prager, E.; Staffler, G.; Stöckl, J.; Kunthalert, D.; Prieschl, E.E.; Baumruker, T.; Burtscher, H.; et al. Characterization of CDw92 as a Member of the Cho-line Transporter-Like Protein Family Regulated Specifically on Dendritic Cells. J. Immunol. 2001, 167, 5795–5804. [Google Scholar] [CrossRef] [Green Version]
- Fullerton, M.D.; Wagner, L.; Yuan, Z.; Bakovic, M. Impaired trafficking of choline transporter-like protein-1 at plasma membrane and inhibition of choline transport in THP-1 monocyte-derived macrophages. Am. J. Physiol. Physiol. 2006, 290, C1230–C1238. [Google Scholar] [CrossRef] [Green Version]
- Długosz, E.; Basałaj, K.; Zawistowska-Deniziak, A. Cytokine production and signalling in human THP-1 macrophages is dependent on Toxocara canis glycans. Parasitol. Res. 2019, 118, 2925–2933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, S.-D.; Martins, A.; Khazaie, K.; Han, J.; Chan, B.M.C.; Kim, S.O. Cathepsin B Is Involved in the Trafficking of TNF-α-Containing Vesicles to the Plasma Membrane in Macrophages. J. Immunol. 2008, 181, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Hoegen, T.; Tremel, N.; Klein, M.; Angele, B.; Wagner, H.; Kirschning, C.; Pfister, H.-W.; Fontana, A.; Hammerschmidt, S.; Koedel, U. The NLRP3 Inflammasome Contributes to Brain Injury in Pneumococcal Meningitis and Is Activated through ATP-Dependent Lysosomal Cathepsin B Release. J. Immunol. 2011, 187, 5440–5451. [Google Scholar] [CrossRef] [Green Version]
- Ehlting, C.; Häussinger, D.; Bode, J.G. Sp3 is involved in the regulation of SOCS3 gene expression. Biochem. J. 2005, 387, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Russell, L.; Garrett-Sinha, L.A. Transcription factor Ets-1 in cytokine and chemokine gene regulation. Cytokine 2010, 51, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Miraghazadeh, B.; Cook, M.C. Nuclear factor-kappaB in autoimmunity: Man and mouse. Front. Immunol. 2018, 9, 613. [Google Scholar] [CrossRef] [Green Version]
- Waindok, P.; Strube, C. Neuroinvasion of Toxocara canis- and T. cati-larvae mediates dynamic changes in brain cytokine and chemokine profile. J. Neuroinflammation 2019, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, E.; Yoshida, Y.; Shan, B.E.; Yamashita, U. Suppression of macrophage interleukin-12 and tumour necrosis fac-tor-alpha production in mice infected with Toxocara canis. Parasite Immunol. 2001, 23, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Zi, M.; Xu, Y. Involvement of cystatin C in immunity and apoptosis. Immunol. Lett. 2018, 196, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Boztug, K.; Järvinen, P.M.; Salzer, E.; Racek, T.; Mönch, S.; Garncarz, W.; Gertz, E.M.; Schäffer, A.A.; Antonopoulos, A.; Haslam, S.M.; et al. JAGN1 deficiency causes aberrant myeloid cell homeostasis and congenital neutropenia. Nat. Genet. 2014, 46, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Wirnsberger, G.; Zwolanek, F.; Stadlmann, J.; Tortola, L.; Liu, S.W.; Perlot, T.; Järvinen, P.; Dürnberger, G.; Kozieradzki, I.; Sarao, R.; et al. Jagunal homolog 1 is a critical regulator of neutrophil function in fungal host defense. Nat. Genet. 2014, 46, 1028–1033. [Google Scholar] [CrossRef]
- Khandagale, A.; Lazzaretto, B.; Carlsson, G.; Sundin, M.; Shafeeq, S.; Römling, U.; Fadeel, B. JAGN1 is required for fungal killing in neutrophil extracellular traps: Implications for severe congenital neutropenia. J. Leukoc. Biol. 2018, 104, 1199–1213. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Ellena, J.; Liu, L.; Szabo, G.; Cafiso, D.; Castle, D. Secretory Carrier Membrane Protein SCAMP2 and Phosphatidylinositol 4,5-Bisphosphate Interactions in the Regulation of Dense Core Vesicle Exocytosis. Biochemistry 2007, 46, 10909–10920. [Google Scholar] [CrossRef]
- Lang, S.; Pfeffer, S.; Lee, P.H.; Cavalié, A.; Helms, V.; Förster, F.; Zimmermann, R. An update on Sec 61 channel functions, mechanisms, and related diseases. Front. Physiol. 2017, 8, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Resende, N.M.; Gazzinelli-Guimarães, P.H.; Barbosa, F.S.; Oliveira, L.M.; Nogueira, D.S.; Gazzinelli-Guimarães, A.C.; Gonçalves, M.T.P.; Amorim, C.C.O.; Oliveira, F.M.S.; Caliari, M.V.; et al. New insights into the immunopathology of early Toxocara canis infection in mice. Parasites Vectors 2015, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, S.; Vegni-Talluri, M.; Banchieri, L.; Esposito, F. The in vitro adherence of murine eosinophils, neutrophils and non-induced and induced macrophages to infective larvae of Toxocara canis (Nematoda, ascarididae). Int. J. Parasitol. 1990, 20, 603–613. [Google Scholar] [CrossRef]
- Thoreau, V.; Bergès, T.; Callebaut, I.; Guillier-Gencik, Z.; Gressin, L.; Bernheim, A.; Karst, F.; Mornon, J.-P.; Kitzis, A.; Chomel, J.C. Molecular Cloning, Expression Analysis, and Chromosomal Localization of Human Syntaxin 8 (STX8). Biochem. Biophys. Res. Commun. 1999, 257, 577–583. [Google Scholar] [CrossRef]
- Bhat, S.S.; Friedmann, K.S.; Knörck, A.; Hoxha, C.; Leidinger, P.; Backes, C.; Meese, E.; Keller, A.; Rettig, J.; Hoth, M.; et al. Syntaxin 8 is required for efficient lytic granule trafficking in cytotoxic T lymphocytes. Biochim. Biophys. Acta 2016, 1863, 1653–1664. [Google Scholar] [CrossRef]
- Golebiewska, E.M.; Harper, M.T.; Williams, C.M.; Savage, J.S.; Goggs, R.; Von Mollard, G.F.; Poole, A.W. Syntaxin 8 regulates platelet dense granule secretion, aggregation, and thrombus stability. J. Biol. Chem. 2015, 290, 1536–1545. [Google Scholar] [CrossRef] [Green Version]
- Logan, M.R.; Lacy, P.; Odemuyiwa, S.O.; Steward, M.; Davoine, F.; Kita, H.; Moqbel, R. A critical role for vesicle-associated mem-brane protein-7 in exocytosis from human eosinophils and neutrophils. Allergy 2006, 61, 777–784. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E. Selenoprotein P—Expression, Functions, and Roles in Mammals Raymond. Biochim. Biophys. Acta 2009, 1790, 1441–1447. [Google Scholar] [CrossRef] [Green Version]
- Burk, R.F.; Hill, K.E.; Motley, A.K.; Austin, L.M.; Norsworthy, B.K. Deletion of Selenoprotein P Upregulates Urinary Sele-nium Excretion and Depresses Whole-body Selenium Content. Biochim. Biophys. Acta 2006, 176, 1789–1793. [Google Scholar] [CrossRef] [Green Version]
- Yeung, K.J.A.; Smith, A.; Zhao, A.; Madden, K.B.; Elfrey, J.; Sullivan, C.; Levander, O.; Urban, J.F.; Shea-Donohue, T. Impact of vitamin E or selenium deficiency on nematode-induced alterations in murine intestinal function. Exp. Parasitol. 2005, 109, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Pilarczyk, B.; Doligalska, M.J.; Donskow-Schmelter, K.; Balicka-Ramisz, A.; Ramisz, A. Selenium supplementation enhances the protective response to Toxocara canis larvae in mice. Parasite Immunol. 2008, 30, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, E.; Withagen, C.; Fonville, M.; Verlaan, A.; Dormans, J.; Van Loveren, H.; Nicoll, G.; Maizels, R.M.; Van Der Giessen, J. Persistent airway hy-per-responsiveness and inflammation in Toxocara canis-infected BALB/c mice. Clin. Exp. Allergy 2005, 35, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, E.; Aranzamendi, C. Toxocara infection and its association with allergic manifestations. Metab. Immune Disord. Drug Targets 2012, 12, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, E.; Brandes, S.; Dormans, J.; Fonville, M.; Hamilton, C.M.; van der Giessen, J. Toxocara canis: Effect of inoculum size on pulmonary pathology and cytokine expression in BALB/c mice. Exp. Parasitol. 2007, 115, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Długosz, E.; Wiśniewski, M. Toxocara canis glycans influence antigen recognition by mouse IgG1 and IgM antibodies. Acta Parasitol. 2016, 61, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Ariestanti, D.M.; Ando, H.; Hirose, S.; Nakamura, N. Targeted Disruption of Ig-Hepta/Gpr116 Causes Emphysema-like Symptoms That Are Associated with Alveolar Macrophage Activation. J. Biol. Chem. 2015, 290, 11032–11040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niaudet, C.; Hofmann, J.J.; Mäe, M.A.; Jung, B.; Gaengel, K.; Vanlandewijck, M.; Ekvärn, E.; Salvado, M.D.; Mehlem, A.; Al Sayegh, S.; et al. Gpr116 Receptor Regulates Distinctive Functions in Pneumocytes and Vascular Endothelium. PLoS ONE 2015, 10, e0137949. [Google Scholar] [CrossRef] [PubMed]
- Autilio, C.; Pérez-Gil, J. Understanding the principle biophysics concepts of pulmonary surfactant in health and disease. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 104, F443–F451. [Google Scholar] [CrossRef]
- Bonser, L.R.; Schroeder, B.W.; Ostrin, L.A.; Baumlin, N.; Olson, J.L.; Salathe, M.; Erle, D.J. The Endoplasmic Reticulum Resident Protein AGR3. Required for Regulation of Ciliary Beat Frequency in the Airway. Am. J. Respir. Cell Mol. Biol. 2015, 53, 536–543. [Google Scholar] [CrossRef] [Green Version]
- Schleimer, R.P.; Berdnikovs, S. Etiology of epithelial barrier dysfunction in patients with type 2 inflammatory diseases. J. Allergy Clin. Immunol. 2017, 139, 1752–1761. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.D.; Biterova, E.; Salin, M.; Wierenga, R.K.; Ruddock, L.W. Crystal structure of human anterior gradient protein 3. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2018, 74, 425–430. [Google Scholar] [CrossRef]
- Finsterer, J.; Auer, H. Neurotoxocariasis. Rev. Inst. Med. Trop. Sao Paulo 2007, 49, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Mallmann, R.T.; Wilmes, T.; Lichvarova, L.; Bührer, A.; Lohmüller, B.; Castonguay, J.; Lacinova, L.; Klugbauer, N. Tetraspanin-13 modulates volt-age-gated CaV2.2 Ca2+ channels. Sci. Rep. 2013, 3, 1777. [Google Scholar] [CrossRef] [Green Version]
- Maio, N.; Kim, K.S.; Singh, A.; Rouault, T.A. A Single Adaptable Cochaperone-Scaffold Complex Delivers Nascent Iron-Sulfur Clusters to Mammalian Respiratory Chain Complexes I–III. Cell Metab. 2017, 25, 945–953.e6. [Google Scholar] [CrossRef] [Green Version]
- Diaz, F.; Enríquez, J.A.; Moraes, C.T. Cells Lacking Rieske Iron-Sulfur Protein Have a Reactive Oxygen Species-Associated Decrease in Respiratory Complexes I and IV. Mol. Cell. Biol. 2012, 32, 415–429. [Google Scholar] [CrossRef] [Green Version]
- Büeler, H. Mitochondrial and Autophagic Regulation of Adult Neurogenesis in the Healthy and Diseased Brain. Int. J. Mol. Sci. 2021, 22, 3342. [Google Scholar] [CrossRef]
- Schedin-Weiss, S.; Inoue, M.; Hromadkova, L.; Teranishi, Y.; Yamamoto, N.G.; Wiehager, B.; Bogdanovic, N.; Winblad, B.; Sandebring-Matton, A.; Frykman, S.; et al. Monoamine oxidase B is elevated in Alzheimer disease neurons, is associated with γ-secretase and regulates neuronal amyloid β-peptide levels. Alzheimer’s Res. Ther. 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.; Heuer, L.; Janecek-Erfurth, E.; Beineke, A.; Strube, C. Histopathological characterization of Toxocara canis- and T. cati-induced neurotoxocarosis in the mouse model. Parasitol. Res. 2019, 118, 2591–2600. [Google Scholar] [CrossRef]
- Chou, C.M.; Lee, Y.L.; Liao, C.W.; Huang, Y.C.; Fan, C.K. Enhanced expressions of neurodegeneration-associated factors, UPS impairment, and excess Aβ accumulation in the hippocampus of mice with persistent cerebral toxocariasis. Parasites Vectors 2017, 10, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.-M.; Fan, C.-K. Significant apoptosis rather autophagy predominates in astrocytes caused by Toxocara canis larval excretory-secretory antigens. J. Microbiol. Immunol. Infect. 2020, 53, 250–258. [Google Scholar] [CrossRef]
- Menzies, F.M.; Fleming, A.; Rubinsztein, D.C. Compromised autophagy and neurodegenerative diseases. Nat. Rev. Neurosci. 2015, 16, 345–357. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. 2021. Available online: http://www.uniprot.org (accessed on 5 March 2021).
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinfor-matics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bait | Interacting Protein | NCBI ID | UniProt ID |
|---|---|---|---|
| Tc-MUC-3 | Tetraspanin 13 (TSPAN13) | NM_014399.3 | O95857 |
| Small integral membrane protein 30 (SMIM30) | NM_001352688.1 | A4D0T7 | |
| RNA polymerase II subunit G (POLR2G) | NM_002696.2 | P62487 | |
| Adhesion G protein-coupled receptor F5 (ADGRF5) | NM_015234.4 | Q8IZF2 | |
| Ubiquinol-cytochrome c reductase, Rieske iron-sulphur polypeptide 1 (UQCRFS1) | NM_006003.2 | P47985 | |
| Anterior gradient 3, protein disulphide isomerase family member (AGR3) | NM_176813.4 | Q8TD06 | |
| DAZ associated protein 2 (DAZAP2) | NM_014764.3 | Q15038 | |
| Transmembrane protein 199 (TMEM199) | NM_152464.2 | Q8N511 | |
| Protein phosphatase 2C-like domain containing 1data (PP2D1) | NM_001252657.1 | A8MPX8 | |
| Tc-CTL-1 | Solute carrier family 44 member 1 (SLC44A1) | NM_080546.4 | Q8WWI5 |
| Selenoprotein P (SELENOP) | NM_001093726.2 | P49908 | |
| Jagunal homolog 1 (JAGN1) | NM_032492.3 | Q8N5M9 | |
| Cathepsin B (CTSB) | NM_001317237.1 | P07858 | |
| Ankyrin repeat and KH domain containing 1 (ANKHD1) | NM_017747.2 | Q8IWZ3 | |
| Tc-TES-26 | Syntaxin 8 (STX-8) | AF062077.1 | Q9UNK0 |
| Cysteine rich secretory protein 2 (CRISP) | NM_001142435.3 | P16562 |
| Construct | Primer Sequence | Restriction Site |
|---|---|---|
| pGBKT7-muc-3 | F: CGGAATTCCAATCGATATTCGCAGCA | EcoRI |
| R: CGGGATCCCGAACAAAAACCGCACGA | BamHI | |
| pGBKT7-ctl-1 | F: CGGAATTCTGCGTCAACAACAATGAC | EcoRI |
| R: CGGGATCCGAGAGGTCTCTTGCATAC | BamHI | |
| pGBKT7-tes-26 | F: CGGAATTCCAACAGTGTATGGACAGC | EcoRI |
| R: CGGGATCCGGCCTGCGATCGATAGAA | BamHI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Długosz, E.; Milewska, M.; Bąska, P. Identification of Toxocara canis Antigen-Interacting Partners by Yeast Two-Hybrid Assay and a Putative Mechanism of These Host–Parasite Interactions. Pathogens 2021, 10, 949. https://doi.org/10.3390/pathogens10080949
Długosz E, Milewska M, Bąska P. Identification of Toxocara canis Antigen-Interacting Partners by Yeast Two-Hybrid Assay and a Putative Mechanism of These Host–Parasite Interactions. Pathogens. 2021; 10(8):949. https://doi.org/10.3390/pathogens10080949
Chicago/Turabian StyleDługosz, Ewa, Małgorzata Milewska, and Piotr Bąska. 2021. "Identification of Toxocara canis Antigen-Interacting Partners by Yeast Two-Hybrid Assay and a Putative Mechanism of These Host–Parasite Interactions" Pathogens 10, no. 8: 949. https://doi.org/10.3390/pathogens10080949
APA StyleDługosz, E., Milewska, M., & Bąska, P. (2021). Identification of Toxocara canis Antigen-Interacting Partners by Yeast Two-Hybrid Assay and a Putative Mechanism of These Host–Parasite Interactions. Pathogens, 10(8), 949. https://doi.org/10.3390/pathogens10080949