
Immunoinformatic Study of Recombinant LigA/BCon1-5 Antigen and Evaluation of Its Diagnostic Potential in Primary and Secondary Binding Tests for Serodiagnosis of Porcine Leptospirosis

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Material and Methods
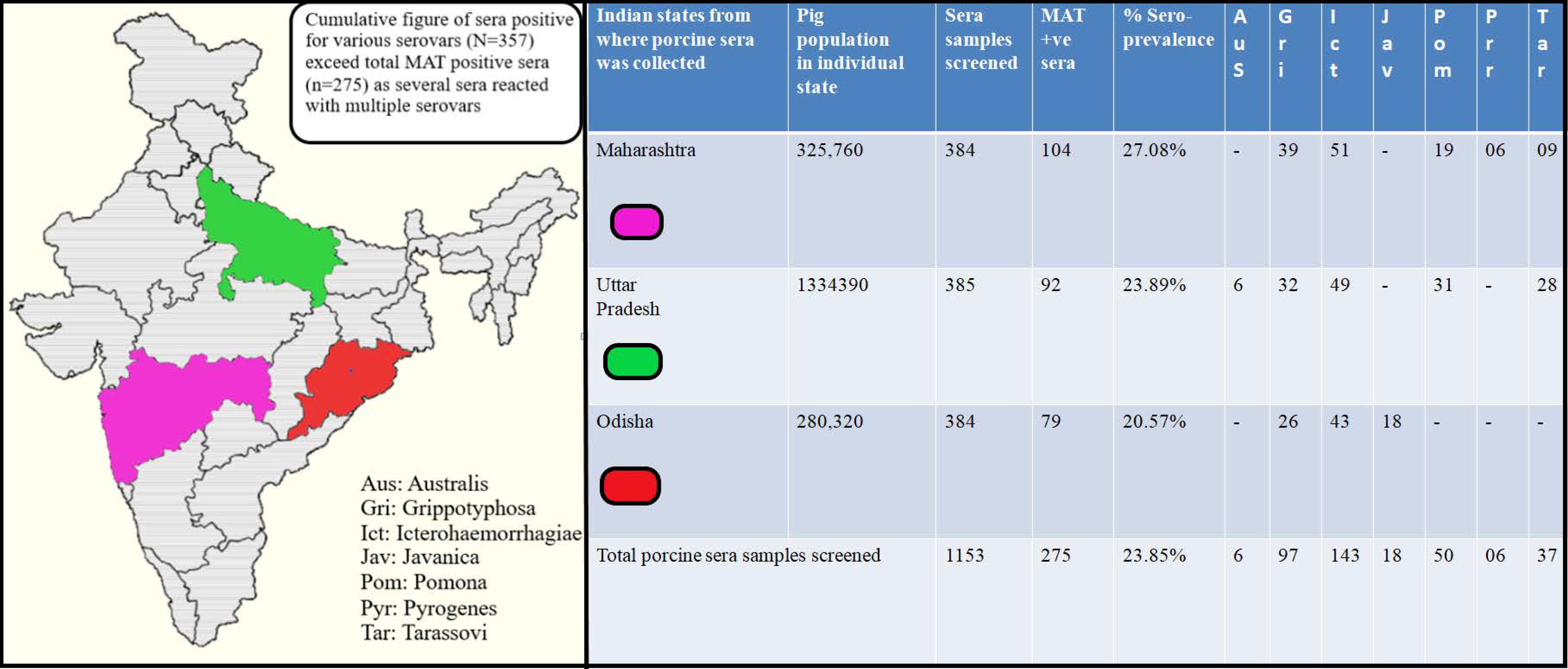
2.1. Serum Sample Collection and Processing
2.2. Leptospiral Serovars and Strains Used in Microscopic Agglutination Test (MAT)
2.3. Microscopic Agglutination Test (MAT)
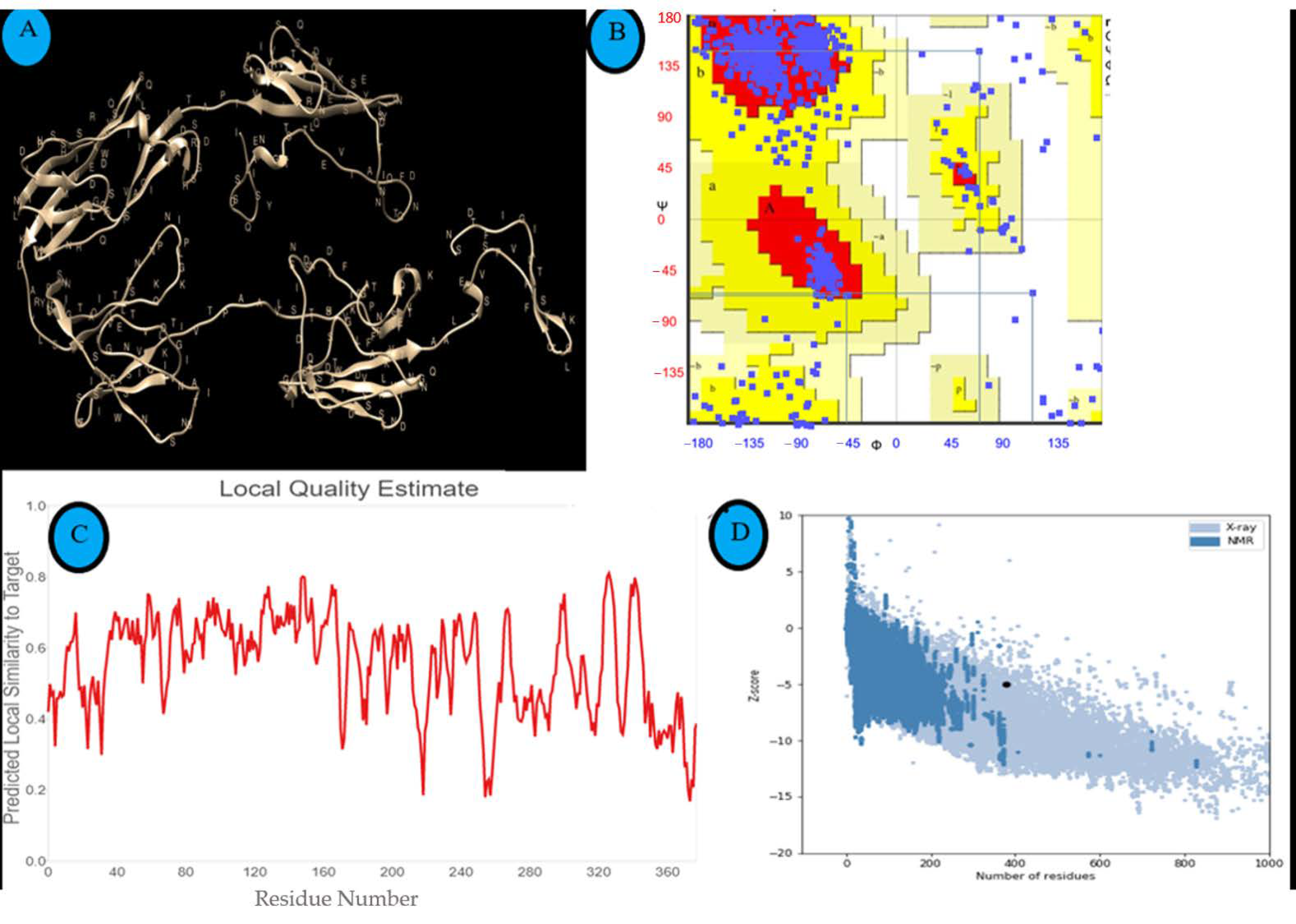
2.4. Prediction of the Three-Dimensional (3D) Conformation of LigA/BCon1-5 Protein
2.5. Assessment and Validation of the Predicted Structure of LigA/BCon1-5 Antigen
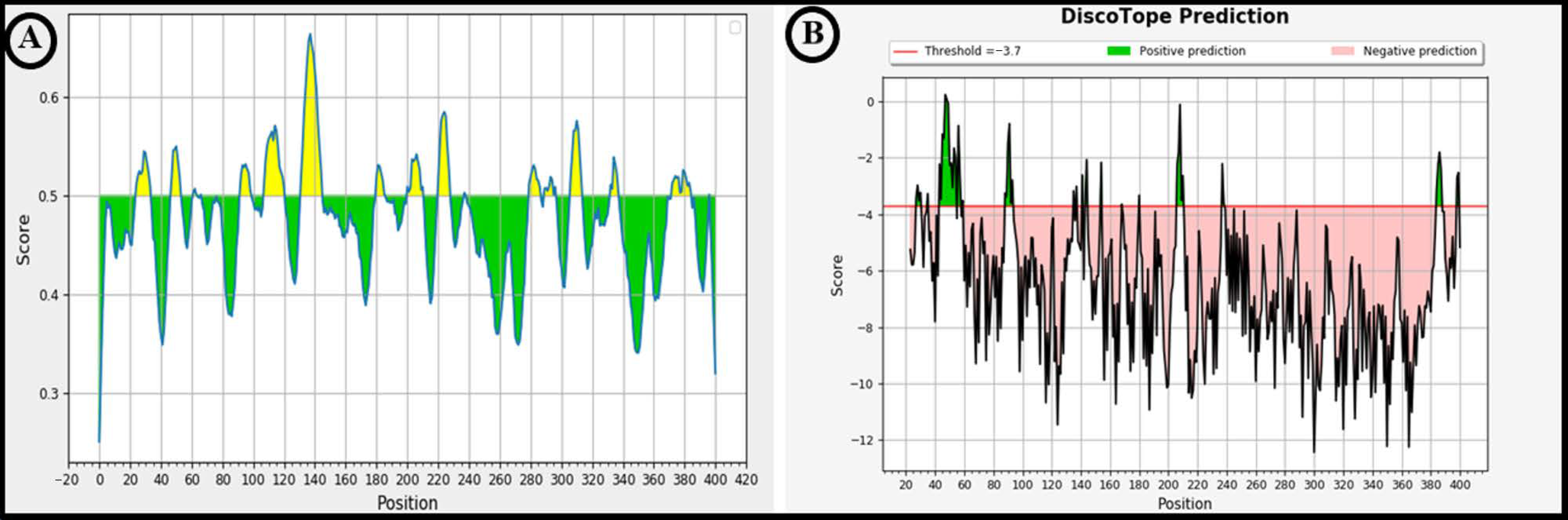
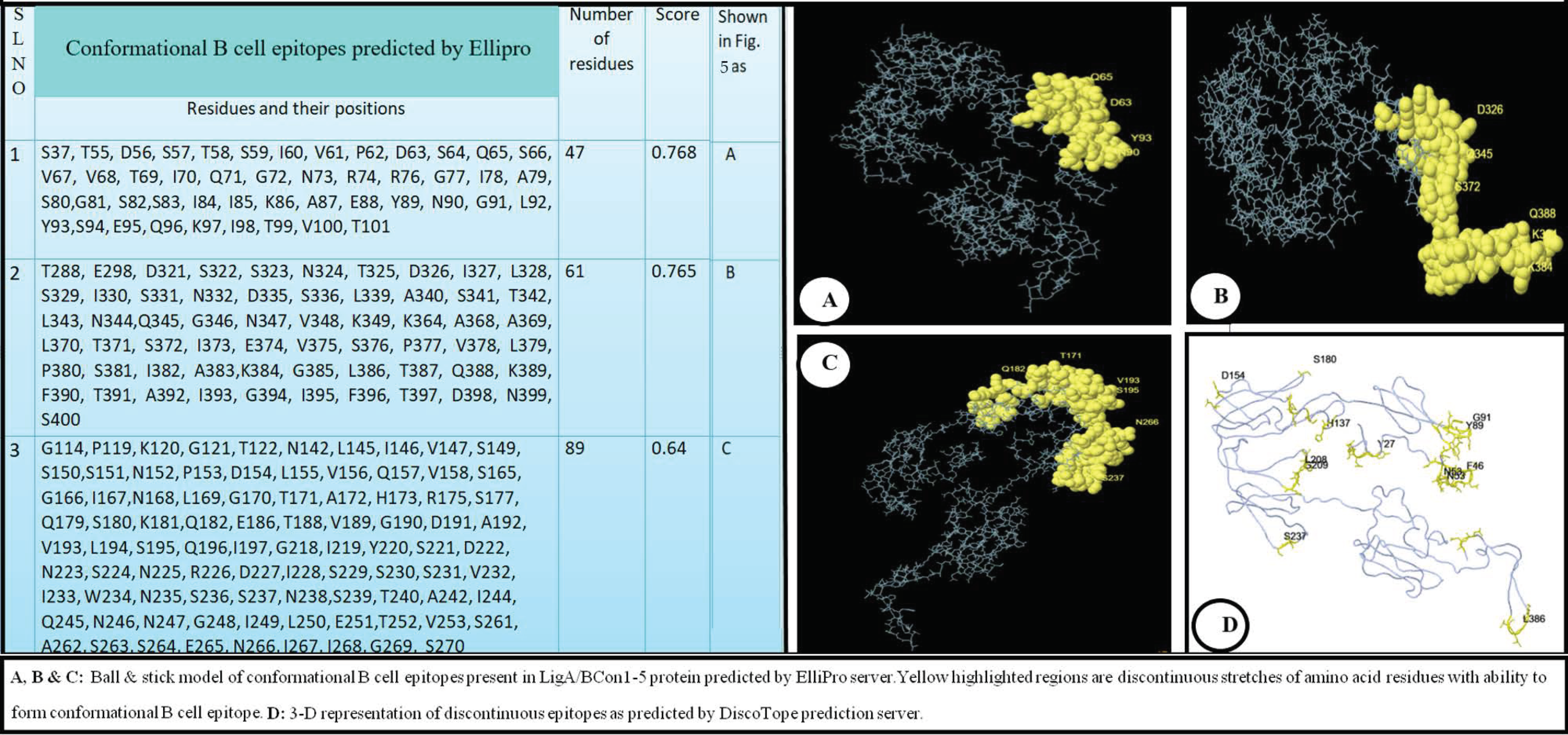
2.6. Prediction of Linear and Conformational (Discontinuous) B-Cell Epitopes Present on LigA/BCon1-5 Protein
2.7. Prediction of T-Cell Epitopes (MHC Class I and II Peptides) Present on LigA/BCon1-5 Protein
2.8. Induction of Expression and Purification of Recombinant LigA/BCon1-5 Antigen
2.9. Western Blot Analysis
2.10. Recombinant LigA/BCon1-5 Based Latex Agglutination Test (rLigA/BCon1-5 Based LAT)
2.11. Recombinant LigA/BCon1-5 Based IgG Dot-ELISA Dipstick Test
2.12. Statistical Analysis
3. Results
3.1. Microscopic Agglutination Test
3.2. Three-Dimensional (3D) Conformation of LigA/BCon1-5 Protein
3.3. Assessment and Validation of the Predicted Structure of LigA/BCon1-5 Antigen
3.4. Prediction of Linear and Conformational (Discontinuous) B-Cell Epitopes Present on LigA/BCon1-5 Protein
3.5. Prediction of T-Cell Epitopes (MHC Class I and IIPeptides) Present on LigA/BCon1-5 Protein
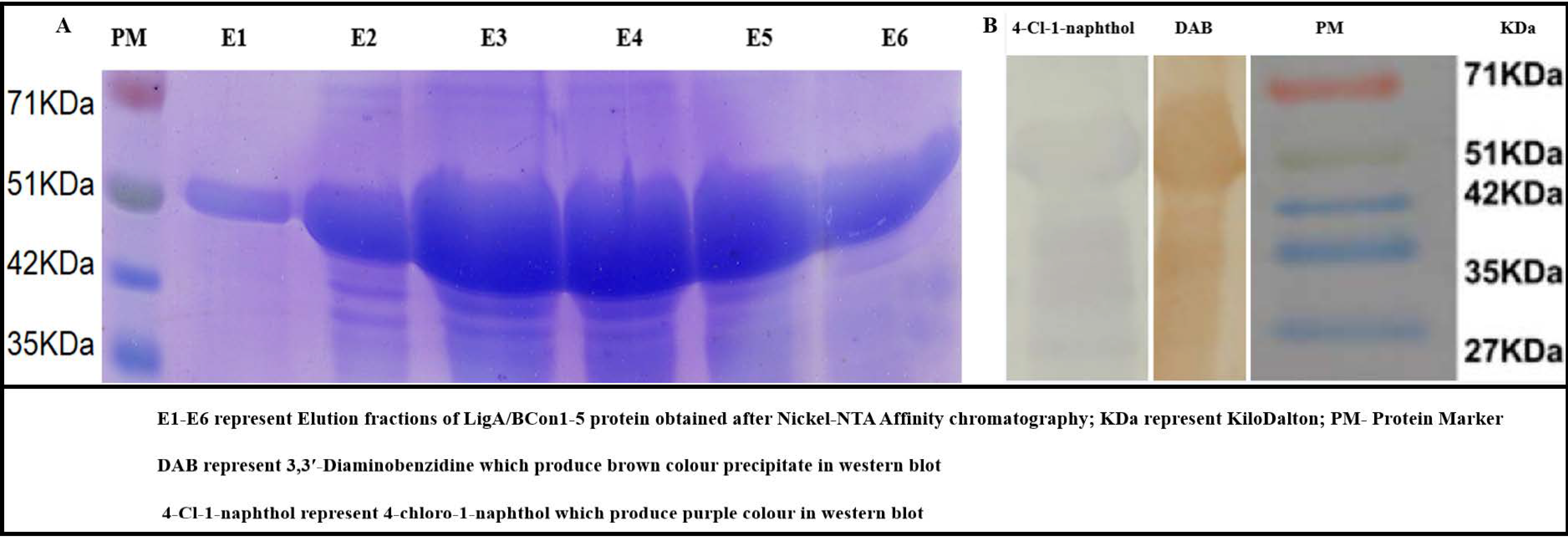
3.6. Recombinant LigA/BCon1-5 Antigen Expression
3.7. Western Blot Analysis
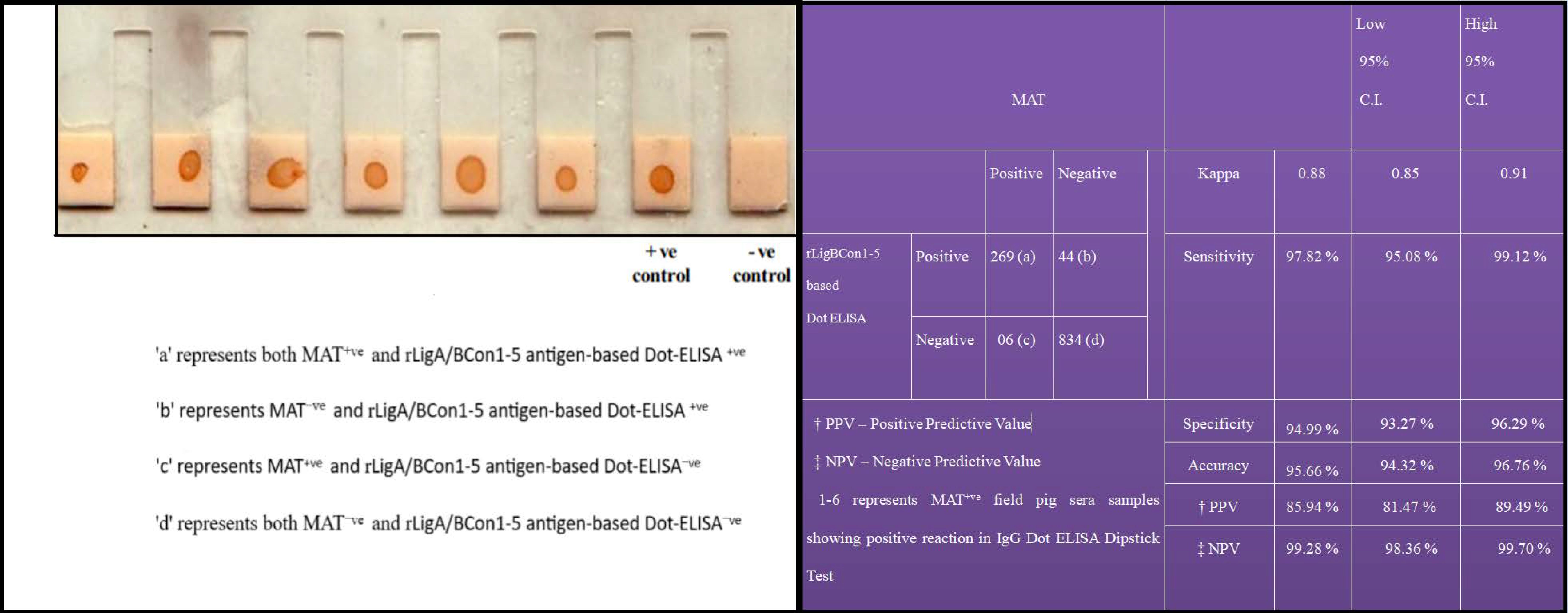
3.8. Recombinant LigA/BCon1-5 Based IgG Dot-ELISA Dipstick Test
3.9. Latex Agglutination Test and Correlation between MAT Titre and LAT Score
4. Discussion
5. Conclusions and Further Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wasiński, B. Infections of swine caused by Leptospira serovars of serogroup Sejroe possibilities of recognition with the use of PCR. Bull. Vet. Inst. Pulawy 2014, 58, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Pozzi, P.; Alborali, G.L.; Etinger, M.; Hadani, Y. Epidemiological Investigation of the Prevalence of Leptospira Spp. in Pigs in Israel. Isr. J. Vet. Med. 2020, 75, 14–21. [Google Scholar]
- Lee, H.S.; Thanh, T.L.; Ly, N.K.; Nguyen-Viet, H.; Thakur, K.K.; Grace, D. Seroprevalence of leptospirosis and Japanese encephalitis in swine in ten provinces of Vietnam. PLoS ONE 2019, 14, e0214701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertasio, C.; Papetti, A.; Scaltriti, E.; Tagliabue, S.; D’Incau, M.; Boniotti, M.B. Serological survey and molecular typing reveal new Leptospira serogroup Pomona strains among pigs of northern Italy. Pathogens 2020, 9, 332. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.A. Leptospirosis. In Diseases of Swine, 8th ed.; Iowa State University Press: Ames, IA, USA, 1999; pp. 483–493. [Google Scholar]
- Strutzberg-Minder, K.; Tschentscher, A.; Beyerbach, M. Passive surveillance of Leptospira infection in swine in Germany. Porc. Health Manag. 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- André-Fontaine, G. Leptospirosis in domestic animals in France: Serological results from 1988 to 2007. Rev. Sci. Tech. 2016, 35, 913–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadsuthi, S.; Bicout, D.J.; Wiratsudakul, A.; Suwancharoen, D.; Petkanchanapong, W.; Modchang, C.; Triampo, W.; Ratanakorn, P.; Chalvet-Monfray, K. Investigation on predominant Leptospira serovars and its distribution in humans and livestock in Thailand, 2010–2015. PLoS Negl. Trop. Dis. 2017, 11, e0005228. [Google Scholar] [CrossRef] [PubMed]
- Budihal, S.V.; Perwez, K. Leptospirosis diagnosis: Competancy of various laboratory tests. J. Clin. Diagn. Res. 2014, 8, 199–202. [Google Scholar] [CrossRef]
- Senthilkumar, T.; Subathra, M.; Phil, M.; Ramadass, P.; Ramaswamy, V. Rapid serodiagnosis of leptospirosis by latex agglutination test and flow-through assay. Indian J. Med. Microbiol. 2008, 26, 45–49. [Google Scholar] [CrossRef]
- Tansuphasiri, U.; Deepradit, S.; Phulsuksombati, D.; Tangkanakul, W. A test strip IgM dot-ELISA assay using leptospiral antigen of endemic strains for serodiagnosis of acute leptospirosis. J. Med. Assoc. Thai. 2005, 88, 391–398. [Google Scholar]
- Shekatkar, S.; Harish, B.N.; Parija, S.C. IgM Dot-ELISA Assay using prevalent Leptospira strain for diagnosis of leptospirosis. Int. J. Collab. Res. Intern. Med. Public Health 2010, 2, 338–346. [Google Scholar]
- Sumarningsih; Tarigan, S.; Susanti; Kusmiyati. Recombinant LipL32 protein for leptospirosis detection in Indonesia. Procedia Chem. 2016, 18, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Joseph, S.; Thomas, N.; Thangapandian, E.; Singh, V.P.; Verma, R.; Srivastava, S.K. Evaluation and comparison of native and recombinant LipL21 protein based ELISAs for diagnosis of bovine leptospirosis. J. Vet. Sci. 2012, 13, 99–101. [Google Scholar] [CrossRef]
- Natarajaseenivasan, K.; Vijayachari, P.; Sharma, S.; Sugunan, A.P.; Selvin, J.; Sehgal, S.C. Serodiagnosis of severe leptospirosis: Evaluation of ELISA based on the recombinant OmpL1 or LipL41 antigens of Leptospira interrogans serovar autumnalis. Ann. Trop. Med. Parasitol. 2008, 102, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Yan, W.; Xiang, H.; He, H.; Yang, M.; Ijaz, M.; Useh, N.; Hsieh, C.L.; McDonough, P.L.; McDonough, S.P.; et al. Recombinant Antigens rLipL21, rLoa22, rLipL32 and rLigACon4-8 for Serological Diagnosis of Leptospirosis by Enzyme-Linked Immunosorbent Assays in Dogs. PLoS ONE 2014, 9, e111367. [Google Scholar] [CrossRef]
- Hoelzle, K.; Grimm, J.; Ritzmann, M.; Heinritzi, K.; Torgerson, P.; Hamburger, A.; Wittenbrink, M.M.; Hoelzle, L.E. Use of recombinant antigens to detect antibodies against Mycoplasma suis with correlation of serological results to hematological findings. Clin. Vaccine Immunol. 2007, 14, 1616–1622. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Chen, Y.; Yan, J. Recombinant Multiepitope Protein for Diagnosis of Leptospirosis. Clin. Vaccine Immunol. 2008, 15, 1711–1714. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, J.; Barocchi, M.A.; Croda, J.; Young, T.A.; Sanchez, Y.; Siqueira, I.; Bolin, C.A.; Reis, M.G.; Riley, L.W.; Haake, D.A.; et al. Pathogenic Leptospira species express surface-exposed proteins belonging to the bacterial immunoglobulin superfamily. Mol. Microbiol. 2003, 49, 929–946. [Google Scholar] [CrossRef]
- Fernandes, L.G.V.; Hornsby, R.L.; Nascimento, A.L.T.O. Genetic manipulation of pathogenic Leptospira: CRISPR interference (CRISPRi)-mediated gene silencing and rapid mutant recovery at 37 °C. Sci. Rep. 2021, 11, 1768. [Google Scholar] [CrossRef]
- Castiblanco-Valencia, M.M.; Fraga, T.R.; Breda, L.C.; Vasconcellos, S.A.; Figueira, C.P.; Picardeau, M.; Wunder, E.; Ko, A.I.; Barbosa, A.S.; Isaac, L. Acquisition of negative complement regulators by the saprophyte Leptospira bifexa expressing LigA or LigB confers enhanced survival in human serum. Immunol. Lett. 2016, 173, 61–68. [Google Scholar] [CrossRef]
- Croda, J.; Ramos, J.G.; Matsunaga, J.; Queiroz, A.; Homma, A.; Riley, L.W.; Haake, D.A.; Reis, M.G.; Ko, A.I. Leptospira immunoglobulin-like proteins as a serodiagnostic marker for acute leptospirosis. J. Clin. Microbiol. 2007, 45, 1528–1534. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, G.M.; McBride, A.J.A.; Picardeau, M.; Ribeiro, S.G.; Moreira, Â.N.; Morel, V.; Reis, M.G.; Ko, A.I.; Dellagostin, O.A. Distribution of the leptospiral immunoglobulin-like (lig) genes in pathogenic Leptospira species and application of ligB to typing leptospiral isolates. J. Med. Microbiol. 2009, 58, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, R.U.; Chang, Y.F.; Hassan, F.; McDonough, S.P.; Pough, M.; Barr, S.C.; Simpson, K.W.; Mohammed, H.O.; Shin, S. Expression of leptospiral immunoglobulin-like protein by Leptospira interrogans and evaluation of its diagnostic potential in a kinetic ELISA. J. Med. Microbiol. 2004, 53, 975–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kihara, D.; Skolnick, J. Local energy landscape flattening: Parallel hyperbolic Monte Carlo sampling of protein folding. Proteins 2002, 48, 192–201. [Google Scholar] [CrossRef]
- Zhang, Y. Template-based modeling and free modeling by I-TASSER in CASP7. Proteins Struct. Funct. Bioinf. 2007, 69, 108–117. [Google Scholar] [CrossRef]
- Thornton, J.M.; Edwards, M.S.; Taylor, W.R.; Barlow, D.J. Location of ‘continuous’ antigenic determinants in the protruding regions of proteins. EMBO J. 1986, 5, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Eswar, N.W.B.; Marti-Renom, M.A.; Madhusudhan, M.S.; Eramian, D.; Shen, M.Y.; Pieper, U.; Sali, A. Comparative Protein Structure Modeling with MODELLER.InCurrent Protocols in Bioinformatics; Wiley: New York, NY, USA, 2006; pp. 561–563. [Google Scholar]
- Webb, B.; Sali, A. Comparative Protein Structure Modeling using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5.6.1–5.6.37. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, J.; Bui, H.H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Andersen, P.H.; Nielsen, M.; Lund, O. Prediction of residues in discontinuous B-cell epitopes using protein 3D structures. Protein Sci. 2006, 15, 2558–2567. [Google Scholar] [CrossRef]
- Grande, T.L. Calculating the Sample Size with a Finite Population in Excel (Video File). 2016. Available online: https://www.youtube.com/watch?v=gLD4tENS82c&t=189s (accessed on 6 June 2021).
- USDA (United States Department of Agriculture). National Veterinary Services Laboratories. Microtitre technique for detection of Leptospira antibodies. Proc. Annu. Meet. US Anim. Health Assoc. 1987, 91, 65–73. [Google Scholar]
- Mishra, P.K.K.; Nimmanapalli, R. In silico characterization of Leptospira interrogans DNA ligase A and delineation of its antimicrobial stretches. Ann. Microbiol. 2019, 69, 1329–1350. [Google Scholar]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [Green Version]
- Bowie, J.U.; Lüthy, R.; Eisenberg, D. A method to identify protein sequences that fold into a known three-dimensional structure. Science 1991, 253, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Lüthy, R.; Bowie, J.U.; Eisenberg, D. Assessment of protein models with three-dimensional profiles. Nature 1992, 356, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of non bonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontius, J.; Richelle, J.; Wodak, S.J. Deviations from Standard Atomic Volumes as a Quality Measure for Protein Crystal Structures. J. Mol. Biol. 1996, 264, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Kringelum, J.V.; Lundegaard, C.; Lund, O.; Nielsen, M. Reliable B Cell Epitope Predictions: Impacts of Method Development and Improved Benchmarking. PLoS Comput. Biol. 2012, 8, e1002829. [Google Scholar] [CrossRef]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48, W449–W454. [Google Scholar] [CrossRef]
- Deneke, Y.; Sabarinath, T.; Gogia, N.; Lalsiamthara, J.; Viswas, K.N.; Chaudhuri, P. Evaluation of recombinant LigB antigen-based indirect ELISA and latex agglutination test for the serodiagnosis of bovine leptospirosis in India. Mol. Cell. Probes 2014, 28, 141–146. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Madhan Mohan, C.; Ramadass, P.; Nachimuthu, K. Recombinant antigen-based Latex Agglutination Test for Rapid Serodiagnosis of leptospirosis. Vet. Res. Commun. 2007, 31, 9–15. [Google Scholar] [CrossRef]
- Smits, H.L.; van der Hoorn, M.A.; Goris, M.G.; Gussenhoven, G.C.; Yersin, C.; Sasaki, D.M. Simple latex agglutination assay for rapid serodiagnosis of human leptospirosis. J. Clin. Microbiol. 2000, 38, 1272–1275. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, R.H. Validation of serological assays for diagnosis of infectious diseases. Rev. Sci. Tech. Off. Int. Epiz. 1998, 17, 469–486. [Google Scholar] [CrossRef]
- Ngugi, J.N.; Fèvre, E.M.; Mgode, G.F.; Obonyo, M.; Mhamphi, G.G.; Otieno, C.A.; Cook, E.A.J. Seroprevalence and associated risk factors of leptospirosis in slaughter pigs; a neglected public health risk, western Kenya. BMC Vet. Res. 2019, 15, 403. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, M.; Umapathy, B.L. Deciphering leptospirosis-a diagnostic mystery, an insight. J. Med Res. Health Sci. 2015, 4, 693–701. [Google Scholar] [CrossRef]
- Okello, A.L.; Burniston, S.; Conlan, J.V.; Inthavong, P.; Khamlome, B.; Welburn, S.C.; Gilbert, J.; Allen, J.; Blacksell, S.D. Prevalence of Endemic Pig-Associated Zoonoses in Southeast Asia: A Review of Findings from the Lao People’s Democratic Republic. Am. J. Trop. Med. Hyg. 2015, 92, 1059–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faine, S.B.; Adler, B.; Bolin, C.; Perolat, P. Leptospira and Leptospirosis, 2nd ed.; MediSci: Melburne, Australia, 1999. [Google Scholar]
- Behera, S.K.; Sabarinath, T.; Chaudhary, P.; Kumar, A.; Das, S.C.; Agarwal, R.K. Evaluation of recombinant LipL32 based latex agglutination test for serodiagnosis of porcine leptospirosis. Vet. World 2014, 7, 17–20. [Google Scholar] [CrossRef]
- Favero, A.C.M.; Pinheiro, S.R.; Vasconcellos, S.A.; Morais, Z.M.; Ferreira, F.; Ferreira Neto, J.S. Most frequent serovars of leptospires in serological tests of buffaloes, sheeps, goats, horses, swines and dogs from several Brazilian states. Cienc. Rural. 2002, 32, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Valença, R.M.B.; Mota, R.A.; Castro, V.; Anderlini, G.A.; Pinheiro Junior, J.W.; Brandespim, D.F.; Guerra, M.M.P. Prevalence and risk factors associated with leptospira spp. infection in technified swine farms in the state of Alagoas, Brazil risk factors associated with leptospira spp. in swine farms. Transbound. Emerg. Dis. 2013, 60, 79–86. [Google Scholar] [CrossRef]
- Delbem, A.C.B.; Freitas, J.C.; Bracarense, A.P.F.R.L.; Muller, E.E.; Oliveira, R.C. Leptospirosis in slaughtered sows: Serological and histopathological investigation. Braz. J. Microbiol. 2002, 33, 174–177. [Google Scholar] [CrossRef] [Green Version]
- Ptak, C.P.; Hsieh, C.L.; Lin, Y.P.; Maltsev, A.S.; Raman, R.; Sharma, Y.; Oswald, R.E.; Chang, Y.F. NMR Solution Structure of the Terminal Immunoglobulin-like Domain from the Leptospira Host-Interacting Outer Membrane Protein, LigB. Biochemistry 2014, 53, 5249–5260. [Google Scholar] [CrossRef]
- Mei, S.; Zhang, J.; Zhang, X.; Tu, X. Solution structure of a bacterial immunoglobulin-like domain of the outer membrane protein (LigB) from Leptospira. Proteins 2015, 83, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.L.; Ptak, C.P.; Tseng, A.; Suguiura, I.M.S.; McDonough, S.P.; Sritrakul, T. Extended low-resolution structure of a Leptospira antigen offers high bactericidal antibody accessibility amenable to vaccine design. eLife 2017, 6, e30051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiltgen, M. Algorithms for Structure Comparison and Analysis: Homology Modelling of Proteins InEncyclopedia of Bioinformatics and Computational Biology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 38–61. [Google Scholar]
- Ramakrishnan, C.; Lakshmi, B.; Kurien, A.; Devipriya, D.; Srinivasan, N. Structural compromise of disallowed conformations in peptide and protein structures. Protein Pept. Lett. 2007, 14, 672–682. [Google Scholar] [CrossRef]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins. Proteins 1993, 17, 355–362. [Google Scholar] [CrossRef]
- Karosiene, E.; Rasmussen, M.; Blicher, T.; Lund, O.; Nielsen, M. NetMHCIIpan-3.0, a common pan-specific MHC class II prediction method including all three human MHC class II isotypes, HLA-DR, HLA-DP and HLA-DQ. Immunogenetics 2013, 65, 711–724. [Google Scholar] [CrossRef]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zheng, H.; Zhang, Y.; Wang, Y.; Zhang, J.; Li, Z.; Cui, S.; Xin, X.; Ye, Q.; Chang, Y.F.; et al. Genomic Analysis of a New Serovar of Leptospira weilii Serogroup Manhao. Front. Microbiol. 2017, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Musso, D.; Scola, B.L. Laboratory diagnosis of leptospirosis: A challenge. J. Microbiol. Immunol. Infect. 2013, 46, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hull-Jackson, C.; Glass, M.B.; Ari, M.D.; Bragg, S.L.; Branch, S.L.; Whittington, C.U.; Edwards, C.N.; Levett, P.N. Evaluation of a commercial latex agglutination assay for serological diagnosis of leptospirosis. J. Clin. Microbiol. 2006, 44, 1853–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.N. | Residue Position | Residue | Contact Number | Propensity Score | Discotope Score |
|---|---|---|---|---|---|
| 1. | 27 | TYR | 2 | −3.591 | −3.408 |
| 2. | 28 | GLN | 3 | −2.973 | −2.976 |
| 3. | 29 | ASP | 7 | −3.074 | −3.525 |
| 4. | 30 | SER | 4 | −3.131 | −3.231 |
| 5. | 35 | GLY | 4 | −3.18 | −3.274 |
| 6. | 43 | THR | 6 | −1.736 | −2.227 |
| 7. | 44 | ALA | 20 | −1.33 | −3.477 |
| 8. | 45 | ILE | 4 | −0.794 | −1.163 |
| 9. | 46 | PHE | 7 | −0.591 | −1.328 |
| 10. | 47 | ASP | 2 | 0.531 | 0.24 |
| 11. | 48 | ASN | 3 | 0.474 | 0.075 |
| 12. | 49 | GLY | 1 | 0.042 | −0.078 |
| 13. | 50 | THR | 7 | −1.634 | −2.251 |
| 14. | 51 | ASN | 7 | −1.566 | −2.19 |
| 15. | 52 | GLN | 15 | −1.506 | −3.058 |
| 16. | 53 | ASN | 3 | −1.485 | −1.659 |
| 17. | 54 | ILE | 1 | −2.099 | −1.973 |
| 18. | 56 | ASP | 0 | −0.973 | −0.861 |
| 19. | 57 | SER | 13 | −1.195 | −2.553 |
| 20. | 59 | SER | 8 | −2.936 | −3.518 |
| 21. | 88 | GLU | 10 | −2.378 | −3.254 |
| 22. | 89 | TYR | 16 | −1.74 | −3.38 |
| 23. | 90 | ASN | 7 | −0.73 | −1.451 |
| 24. | 91 | GLY | 1 | −0.767 | −0.793 |
| 25. | 92 | LEU | 19 | −1.608 | −3.608 |
| 26. | 93 | TYR | 6 | −2.374 | −2.791 |
| 27. | 135 | GLY | 1 | −3.453 | −3.171 |
| 28. | 137 | HIS | 12 | −1.846 | −3.014 |
| 29. | 141 | SER | 6 | −2.173 | −2.613 |
| 30. | 143 | ASP | 9 | −2.552 | −3.294 |
| 31. | 144 | PRO | 0 | −2.338 | −2.069 |
| 32. | 154 | ASP | 0 | −2.449 | −2.167 |
| 33. | 168 | ASN | 8 | −3.083 | −3.649 |
| 34. | 180 | SER | 7 | −2.855 | −3.332 |
| 35. | 206 | ILE | 8 | −1.525 | −2.269 |
| 36. | 207 | PRO | 8 | −1.009 | −1.813 |
| 37. | 208 | LEU | 0 | −0.125 | −0.111 |
| 38. | 209 | GLY | 14 | −2.144 | −3.507 |
| 39. | 210 | LYS | 1 | −2.851 | −2.638 |
| 40. | 237 | SER | 0 | −2.495 | −2.208 |
| 41. | 238 | ASN | 9 | −2.897 | −3.599 |
| 42. | 384 | LYS | 6 | −2.736 | −3.111 |
| 43. | 385 | GLY | 0 | −2.522 | −2.232 |
| 44. | 386 | LEU | 0 | −2.034 | −1.8 |
| 45. | 387 | THR | 3 | −2.273 | −2.357 |
| 46. | 398 | ASP | 0 | −3.043 | −2.693 |
| 47. | 399 | ASN | 0 | −2.853 | −2.525 |
| S.N. | Peptide Sequence | Allele | Position | Score |
|---|---|---|---|---|
| 1. | VPDSQSVVTI | SLA-1*0101 | 61–70 | 0.245636 |
| 2. | IEVSPVLPSI | SLA-6*0101 | 373–382 | 0.210248 |
| 3. | HQDISNDPLI | SLA-1*0101 | 137–146 | 0.207865 |
| 4. | ETVDTGIVTI | SLA-1*0101 | 251–260 | 0.203195 |
| 5. | SGSSIIKAEY | SLA-2*0101 | 80–89 | 0.182214 |
| 6. | SIANGTSTTL | SLA-1*0101 | 31–40 | 0.178052 |
| 7. | SIAKGLTQKF | SLA-2*0101 | 381–390 | 0.163866 |
| 8. | SHQDISNDPL | SLA-1*0101 | 136–145 | 0.163175 |
| 9. | SENIIGSVKL | SLA-6*0101 | 264–273 | 0.162667 |
| 10. | STDFKVTQAA | SLA-1*0101 | 360–369 | 0.15193 |
| S.N. | Peptide Sequence | Allele | Position | Adjusted Rank * | Method Used |
|---|---|---|---|---|---|
| 1. | GSVKLIVTPAALVSI | HLA-DRB1*01:01 | 269–283 | 0.91 | Consensus (comb.lib./smm/nn) |
| 2. | GSTDFKVTQAALTSI | HLA-DRB1*01:01 | 359–373 | 1.10 | Consensus (comb.lib./smm/nn) |
| 3. | LSFFHLLLGNSNPTI | HLA-DRB1*01:01 | 6–20 | 1.30 | Consensus (comb.lib./smm/nn) |
| 4. | QSVVTIQGNRVRGIA | HLA-DRB1*01:01 | 65–79 | 3.60 | Consensus (comb.lib./smm/nn) |
| 5. | FHLLLGNSNPTITRI | HLA-DRB1*01:01 | 9–23 | 5.40 | Consensus (comb.lib./smm/nn) |
| 6. | PLIVWSSSNPDLVQV | HLADQA1*01:01/DQB1*02:01 | 144–158 | 7.30 | NetMHCIIpan |
| 7. | GKKQKLIATGIYSDN | HLA-DRB1*01:01 | 209–223 | 12 | Consensus (comb.lib./smm/nn) |
| 8. | AEEMTVGDAVLSQIQ | HLADQA1*01:01/DQB1*02:01 | 184–198 | 13.00 | NetMHCIIpan |
| 9. | KKQKLIATGIYSDNS | HLA-DRB1*01:01 | 210–224 | 13.0 | Consensus (comb.lib./smm/nn) |
| 10. | NRVRGIASGSSIIKA | HLA-DRB1*01:01 | 73–87 | 14.0 | Consensus (comb.lib./smm/nn) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behera, S.K.; Sabarinath, T.; Mishra, P.K.K.; Deneke, Y.; Kumar, A.; ChandraSekar, S.; Senthilkumar, K.; Verma, M.; Ganesh, B.; Gurav, A.; et al. Immunoinformatic Study of Recombinant LigA/BCon1-5 Antigen and Evaluation of Its Diagnostic Potential in Primary and Secondary Binding Tests for Serodiagnosis of Porcine Leptospirosis. Pathogens 2021, 10, 1082. https://doi.org/10.3390/pathogens10091082
Behera SK, Sabarinath T, Mishra PKK, Deneke Y, Kumar A, ChandraSekar S, Senthilkumar K, Verma M, Ganesh B, Gurav A, et al. Immunoinformatic Study of Recombinant LigA/BCon1-5 Antigen and Evaluation of Its Diagnostic Potential in Primary and Secondary Binding Tests for Serodiagnosis of Porcine Leptospirosis. Pathogens. 2021; 10(9):1082. https://doi.org/10.3390/pathogens10091082
Chicago/Turabian StyleBehera, Sujit Kumar, Thankappan Sabarinath, Prasanta Kumar K. Mishra, Yosef Deneke, Ashok Kumar, Shanmugam ChandraSekar, Kuppusamy Senthilkumar, MedRam Verma, Balasubramanian Ganesh, Amol Gurav, and et al. 2021. "Immunoinformatic Study of Recombinant LigA/BCon1-5 Antigen and Evaluation of Its Diagnostic Potential in Primary and Secondary Binding Tests for Serodiagnosis of Porcine Leptospirosis" Pathogens 10, no. 9: 1082. https://doi.org/10.3390/pathogens10091082
APA StyleBehera, S. K., Sabarinath, T., Mishra, P. K. K., Deneke, Y., Kumar, A., ChandraSekar, S., Senthilkumar, K., Verma, M., Ganesh, B., Gurav, A., & Hota, A. (2021). Immunoinformatic Study of Recombinant LigA/BCon1-5 Antigen and Evaluation of Its Diagnostic Potential in Primary and Secondary Binding Tests for Serodiagnosis of Porcine Leptospirosis. Pathogens, 10(9), 1082. https://doi.org/10.3390/pathogens10091082