
Challenges in Drug Discovery for Intracellular Bacteria


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Challenges in Drug Discovery against Intracellular Bacteria
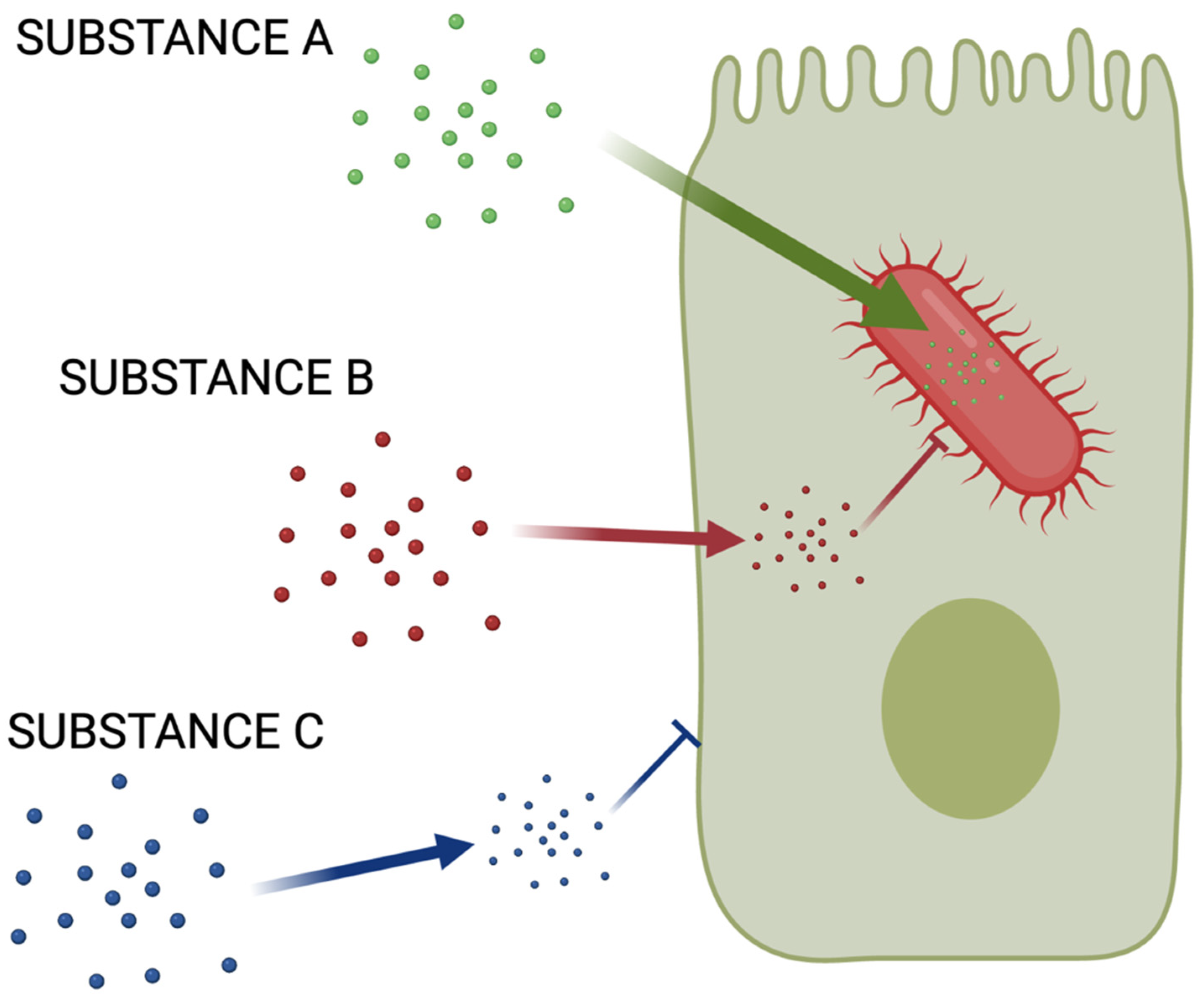
2.1. Chemical Space Is a Severe Limitation
2.2. Finding Good Drug Targets
2.3. Current Models of Studying Virulence
3. Perspective
- Find better targets.
- Develop cheaper, more effective models (maybe in vitro, for instance broth microdilution).
- Understanding chemical space that is best to permeate both human and bacterial cells.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 22 January 2021).
- Rowley, J.; Hoorn, S.V.; Korenromp, E.; Low, N.; Unemo, M.; Abu-Raddad, L.J.; Chico, R.M.; Smolak, A.; Newman, L.; Gottlieb, S.; et al. Chlamydia, Gonorrhoea, Trichomoniasis and Syphilis: Global Prevalence and Incidence Estimates, 2016. Bull. World Health Organ. 2019, 97, 548–562P. [Google Scholar] [CrossRef]
- Millet, J.P.; Orcau, A.; de Olalla, P.G.; Casals, M.; Rius, C.; Cayla, J.A. Tuberculosis Recurrence and Its Associated Risk Factors among Successfully Treated Patients. J. Epidemiol. Community Health 2009, 63, 799–804. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. 2019 Antibacterial Agents in Clinical Development: An Analysis of the Antibacterial Clinical Development Pipeline. Available online: https://www.who.int/publications/i/item/9789240000193 (accessed on 22 January 2021).
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 22 January 2021).
- Moulder, J.W. Comparative Biology of Intracellular Parasitism. Microbiol. Rev. 1985, 49, 298–337. [Google Scholar] [CrossRef]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, 1356540. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable Deaths and Disability-Adjusted Life-Years Caused by Infections with Antibiotic-Resistant Bacteria in the Eu and the European Economic Area in 2015: A Population-Level Modelling Analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Eggleston, K.; Rotimi, V.; Zeckhauser, R.J. Antibiotic Resistance as a Global Threat: Evidence from China, Kuwait and the United States. Glob. Health 2006, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Garneau-Tsodikova, S. Resisting Resistance: Gearing up for War. MedChemCommun 2019, 10, 1512–1516. [Google Scholar] [CrossRef]
- Daley, C.L.; Iaccarino, J.M.; Lange, C.; Cambau, E.; Wallace, R.J.; Andrejak, C.; Bottger, E.C.; Brozek, J.; Griffith, D.E.; Guglielmetti, L.; et al. Treatment of Nontuberculous Mycobacterial Pulmonary Disease: An Official Ats/Ers/Escmid/Idsa Clinical Practice Guideline: Executive Summary. Clin. Infect. Dis. 2020, 71, e1–e36. [Google Scholar] [CrossRef] [PubMed]
- Nahid, P.; Dorman, S.E.; Alipanah, N.; Barry, P.M.; Brozek, J.L.; Cattamanchi, A.; Chaisson, L.H.; Chaisson, R.E.; Daley, C.L.; Grzemska, M.; et al. Official American Thoracic Society/Centers for Disease Control and Prevention/Infectious Diseases Society of America Clinical Practice Guidelines: Treatment of Drug-Susceptible Tuberculosis. Clin. Infect. Dis. 2016, 63, e147–e195. [Google Scholar] [CrossRef]
- Tunkel, A.R.; Hartman, B.J.; Kaplan, S.L.; Kaufman, B.A.; Roos, K.L.; Scheld, W.M.; Whitley, R.J. Practice Guidelines for the Management of Bacterial Meningitis. Clin. Infect. Dis. 2004, 39, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, D.N.; Chambers, H.F.; Saag, M.S.; Pavia, A.T.; Black, D.; Boucher, H.W.; Freedman, D.O.; Kim, K.; Schwartz, B.S. (Eds.) The Sanford Guide to Antimicrobial Therapy 2020, 50th ed.; Antimicrobial Therapy, Inc.: Sperryville, VA, USA, 2020. [Google Scholar]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An Official Ats/Idsa Statement: Diagnosis, Treatment, and Prevention of Nontuberculous Mycobacterial Diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular Bacterial Biofilm-Like Pods in Urinary Tract Infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Justice, S.S.; Hung, C.; Theriot, J.A.; Fletcher, D.A.; Anderson, G.G.; Footer, M.J.; Hultgren, S.J. Differentiation and Developmental Pathways of Uropathogenic Escherichia Coli in Urinary Tract Pathogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 1333–1338. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.S.; Bastidas, R.J.; Saka, H.A.; Carpenter, V.K.; Richards, K.L.; Plano, G.V.; Valdivia, R.H. The Chlamydia Trachomatis Type Iii Secretion Chaperone Slc1 Engages Multiple Early Effectors, Including Tepp, a Tyrosine-Phosphorylated Protein Required for the Recruitment of Crki-Ii to Nascent Inclusions and Innate Immune Signaling. PLoS Pathog. 2014, 10, e1003954. [Google Scholar] [CrossRef]
- Kawana, K.; Quayle, A.J.; Ficarra, M.; Ibana, J.A.; Shen, L.; Kawana, Y.; Yang, H.; Marrero, L.; Yavagal, S.; Greene, S.J.; et al. Cd1d Degradation in Chlamydia Trachomatis-Infected Epithelial Cells Is the Result of Both Cellular and Chlamydial Proteasomal Activity. J. Biol. Chem. 2007, 282, 7368–7375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for Bad Bugs: Confronting the Challenges of Antibacterial Discovery. Nat. Rev. Drug. Discov. 2007, 6, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.F.; Holmes, D.J.; Payne, D.J. Finding the Gems Using Genomic Discovery: Antibacterial Drug Discovery Strategies—The Successes and the Challenges. Drug Discov. Today Ther. Strateg. 2004, 1, 519–527. [Google Scholar] [CrossRef]
- Brown, E.D.; Wright, G.D. Antibacterial Drug Discovery in the Resistance Era. Nature 2016, 529, 336–343. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, R.; Moser, H.E. Physicochemical Properties of Antibacterial Compounds: Implications for Drug Discovery. J. Med. Chem. 2008, 51, 2871–2878. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Drug-Like Properties and the Causes of Poor Solubility and Poor Permeability. J. Pharmacol. Toxicol. Methods 2001, 44, 235–249. [Google Scholar] [CrossRef]
- Ebejer, J.P.; Charlton, M.H.; Finn, P.W. Are the Physicochemical Properties of Antibacterial Compounds Really Different from Other Drugs? J. Cheminform. 2016, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A. Enabling Design of Screening Libraries for Antibiotic Discovery by Modeling Chembl Data. Eur. J. Pharm. Sci. 2019, 143, 105166. [Google Scholar] [CrossRef]
- Parker, E.N.; Drown, B.S.; Geddes, E.J.; Lee, H.Y.; Ismail, N.; Lau, G.W.; Hergenrother, P.J. Implementation of Permeation Rules Leads to a Fabi Inhibitor with Activity against Gram-Negative Pathogens. Nat. Microbiol. 2020, 5, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.F.; Drown, B.S.; Riley, A.P.; Garcia, A.; Shirai, T.; Svec, R.L.; Hergenrother, P.J. Predictive Compound Accumulation Rules Yield a Broad-Spectrum Antibiotic. Nature 2017, 545, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.J.; Krishnamoorthy, G.; Wolloscheck, D.; Walker, J.K.; Rybenkov, V.V.; Parks, J.M.; Zgurskaya, H.I. Molecular Properties That Define the Activities of Antibiotics in Escherichia Coli and Pseudomonas Aeruginosa. ACS Infect. Dis. 2018, 4, 1223–1234. [Google Scholar] [CrossRef]
- Krishnamoorthy, G.; Leus, I.V.; Weeks, J.W.; Wolloscheck, D.; Rybenkov, V.V.; Zgurskaya, H.I. Synergy between Active Efflux and Outer Membrane Diffusion Defines Rules of Antibiotic Permeation into Gram-Negative Bacteria. mBio 2017, 8, e01172-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamoorthy, G.; Wolloscheck, D.; Weeks, J.W.; Croft, C.; Rybenkov, V.V.; Zgurskaya, H.I. Breaking the Permeability Barrier of Escherichia Coli by Controlled Hyperporination of the Outer Membrane. Antimicrob. Agents Chemother. 2016, 60, 7372–7381. [Google Scholar] [CrossRef] [Green Version]
- Mansbach, R.A.; Leus, I.V.; Mehla, J.; Lopez, C.A.; Walker, J.K.; Rybenkov, V.V.; Hengartner, N.W.; Zgurskaya, H.I.; Gnanakaran, S. Machine Learning Algorithm Identifies an Antibiotic Vocabulary for Permeating Gram-Negative Bacteria. J. Chem. Inf. Model. 2020, 60, 2838–2847. [Google Scholar] [CrossRef]
- Saha, P.; Sikdar, S.; Krishnamoorthy, G.; Zgurskaya, H.I.; Rybenkov, V.V. Drug Permeation against Efflux by Two Transporters. ACS Infect. Dis. 2020, 6, 747–758. [Google Scholar] [CrossRef]
- Vergalli, J.; Atzori, A.; Pajovic, J.; Dumont, E.; Malloci, G.; Masi, M.; Vargiu, A.V.; Winterhalter, M.; Refregiers, M.; Ruggerone, P.; et al. The Challenge of Intracellular Antibiotic Accumulation, a Function of Fluoroquinolone Influx versus Bacterial Efflux. Commun. Biol. 2020, 3, 198. [Google Scholar] [CrossRef]
- Westfall, D.A.; Krishnamoorthy, G.; Wolloscheck, D.; Sarkar, R.; Zgurskaya, H.I.; Rybenkov, V.V. Bifurcation Kinetics of Drug Uptake by Gram-Negative Bacteria. PLoS ONE 2017, 12, e0184671. [Google Scholar] [CrossRef] [Green Version]
- Zgurskaya, H.I.; Rybenkov, V.V. Permeability Barriers of Gram-Negative Pathogens. Ann. N. Y. Acad. Sci. 2020, 1459, 5–18. [Google Scholar] [CrossRef]
- El Zahed, S.S.; French, S.; Farha, M.A.; Kumar, G.; Brown, E.D. Physicochemical and Structural Parameters Contributing to the Antibacterial Activity and Efflux Susceptibility of Small Molecule Inhibitors of Escherichia Coli. Antimicrob. Agents Chemother. 2021, 65, e01925-20. [Google Scholar] [CrossRef] [PubMed]
- Tommasi, R.; Iyer, R.; Miller, A.A. Antibacterial Drug Discovery: Some Assembly Required. ACS Infect. Dis. 2018, 4, 686–695. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. Host-Pathogen Interactions: Redefining the Basic Concepts of Virulence and Pathogenicity. Infect. Immun. 1999, 67, 3703–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassenaar, T.M.; Gaastra, W. Bacterial Virulence: Can We Draw the Line? FEMS Microbiol. Lett. 2001, 201, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Wang, Y.; Lilburn, T. A Comparative Genomics, Network-Based Approach to Understanding Virulence in Vibrio Cholerae. J. Bacteriol. 2009, 191, 6262–6272. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. Vfdb: A Reference Database for Bacterial Virulence Factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Xiong, Z.; Sun, L.; Yang, J.; Jin, Q. Vfdb 2012 Update: Toward the Genetic Diversity and Molecular Evolution of Bacterial Virulence Factors. Nucleic Acids Res 2012, 40, D641–D645. [Google Scholar] [CrossRef]
- Zhou, C.E.; Smith, J.; Lam, M.; Zemla, A.; Dyer, M.D.; Slezak, T. Mvirdb—A Microbial Database of Protein Toxins, Virulence Factors and Antibiotic Resistance Genes for Bio-Defence Applications. Nucleic Acids Res. 2007, 35, D391–D394. [Google Scholar] [CrossRef]
- Zheng, L.-L.; Li, Y.-X.; Ding, J.; Guo, X.-K.; Feng, K.-Y.; Wang, Y.-J.; Hu, L.-L.; Cai, Y.-D.; Hao, P.; Chou, K.-C. A Comparison of Computational Methods for Identifying Virulence Factors. PLoS ONE 2012, 7, e42517. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.C.; Coleman, R.G.; Smyth, K.T.; Cao, Q.; Soulard, P.; Caffrey, D.R.; Salzberg, A.C.; Huang, E.S. Structure-Based Maximal Affinity Model Predicts Small-Molecule Druggability. Nat. Biotechnol. 2007, 25, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Desaphy, J.; Azdimousa, K.; Kellenberger, E.; Rognan, D. Comparison and Druggability Prediction of Protein-Ligand Binding Sites from Pharmacophore-Annotated Cavity Shapes. J. Chem. Inf. Model. 2012, 52, 2287–2299. [Google Scholar] [CrossRef]
- Fauman, E.B.; Rai, B.K.; Huang, E.S. Structure-Based Druggability Assessment--Identifying Suitable Targets for Small Molecule Therapeutics. Curr. Opin. Chem. Biol. 2011, 15, 463–468. [Google Scholar] [CrossRef]
- Hajduk, P.J.; Huth, J.R.; Fesik, S.W. Druggability Indices for Protein Targets Derived from Nmr-Based Screening Data. J. Med. Chem. 2005, 48, 2518–2525. [Google Scholar] [CrossRef]
- Halgren, T.A. Identifying and Characterizing Binding Sites and Assessing Druggability. J. Chem. Inf. Model. 2009, 49, 377–389. [Google Scholar] [CrossRef]
- Henrich, S.; Salo-Ahen, O.M.; Huang, B.; Rippmann, F.F.; Cruciani, G.; Wade, R.C. Computational Approaches to Identifying and Characterizing Protein Binding Sites for Ligand Design. J. Mol. Recognit. 2010, 23, 209–219. [Google Scholar] [CrossRef]
- Keller, T.H.; Pichota, A.; Yin, Z. A Practical View of ‘Druggability’. Curr. Opin. Chem. Biol. 2006, 10, 357–361. [Google Scholar] [CrossRef]
- Schmidtke, P.; Barril, X. Understanding and Predicting Druggability. A High-Throughput Method for Detection of Drug Binding Sites. J. Med. Chem. 2010, 53, 5858–5867. [Google Scholar] [CrossRef] [PubMed]
- Seco, J.; Luque, F.J.; Barril, X. Binding Site Detection and Druggability Index from First Principles. J. Med. Chem. 2009, 52, 2363–2371. [Google Scholar] [CrossRef]
- Volkamer, A.; Kuhn, D.; Grombacher, T.; Rippmann, F.; Rarey, M. Combining Global and Local Measures for Structure-Based Druggability Predictions. J. Chem. Inf. Model. 2012, 52, 360–372. [Google Scholar] [CrossRef]
- Volkamer, A.; Rarey, M. Exploiting Structural Information for Drug-Target Assessment. Future Med. Chem. 2014, 6, 319–331. [Google Scholar] [CrossRef]
- Krasowski, A.; Muthas, D.; Sarkar, A.; Schmitt, S.; Brenk, R. Drugpred: A Structure-Based Approach to Predict Protein Druggability Developed Using an Extensive Nonredundant Data Set. J. Chem. Inf. Model. 2011, 51, 2829–2842. [Google Scholar] [CrossRef]
- Sarkar, A.; Brenk, R. To Hit or Not to Hit, That Is the Question—Genome-Wide Structure-Based Druggability Predictions for Pseudomonas Aeruginosa Proteins. PLoS ONE 2015, 10, e0137279. [Google Scholar]
- Thurlow, L.R.; Joshi, G.S.; Richardson, A.R. Virulence Strategies of the Dominant Usa300 Lineage of Community-Associated Methicillin-Resistant Staphylococcus Aureus (Ca-Mrsa). FEMS Immunol. Med. Microbiol. 2012, 65, 5–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, A.; Vranken, T.; Malhotra, A.; Arts, J.J.C.; Habibovic, P. In Vitro Antimicrobial Susceptibility Testing Methods: Agar Dilution to 3d Tissue-Engineered Models. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 187–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, S.L.; Mekalanos, J.J.; Holden, D.W. In Vivo Genetic Analysis of Bacterial Virulence. Annu. Rev. Microbiol. 1999, 53, 129–154. [Google Scholar] [CrossRef] [Green Version]
- Totsika, M. Benefits and Challenges of Antivirulence Antimicrobials at the Dawn of the Post-Antibiotic Era. Curr. Med. Chem. 2016, 6, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Duraiswamy, S.; Chee, J.L.Y.; Chen, S.; Yang, E.; Lees, K.; Chen, S.L. Purification of Intracellular Bacterial Communities during Experimental Urinary Tract Infection Reveals an Abundant and Viable Bacterial Reservoir. Infect. Immun. 2018, 86, e00740-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhede, M.; Kragh, K.N.; Qvortrup, K.; Allesen-Holm, M.; van Gennip, M.; Christensen, L.D.; Jensen, P.Ø.; Nielsen, A.K.; Parsek, M.; Wozniak, D.; et al. Phenotypes of Non-Attached Pseudomonas Aeruginosa Aggregates Resemble Surface Attached Biofilm. PLoS ONE 2011, 6, e27943. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.E.; Kragh, K.N.; Bjarnsholt, T.; Diggle, S.P. The Limitations of in Vitro Experimentation in Understanding Biofilms and Chronic Infection. J. Mol. Biol. 2015, 427, 3646–3661. [Google Scholar] [CrossRef]
- Shi, D.; Mi, G.; Wang, M.; Webster, T. In Vitro and Ex Vivo Systems at the Forefront of Infection Modeling and Drug Discovery. Biomaterials 2018, 198, 228–249. [Google Scholar] [CrossRef] [PubMed]
- Begun, J.; Gaiani, J.M.; Rohde, H.; Mack, D.; Calderwood, S.B.; Ausubel, F.M.; Sifri, C.D. Staphylococcal Biofilm Exopolysaccharide Protects against Caenorhabditis Elegans Immune Defenses. PLoS Pathog. 2007, 3, e57. [Google Scholar] [CrossRef] [Green Version]
- Sifri, C.D.; Baresch-Bernal, A.; Calderwood, S.B.; von Eiff, C. Virulence of Staphylococcus Aureus Small Colony Variants in the Caenorhabditis Elegans Infection Model. Infect. Immun. 2006, 74, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The Worm Has Turned--Microbial Virulence Modeled in Caenorhabditis Elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar] [CrossRef]
- Sifri, C.D.; Begun, J.; Ausubel, F.M.; Calderwood, S.B. Caenorhabditis Elegans as a Model Host for Staphylococcus Aureus Pathogenesis Caenorhabditis Elegans as a Model Host for Staphylococcus Aureus Pathogenesis. Infect. Immun. 2003, 71, 2208–2217. [Google Scholar] [CrossRef] [Green Version]
- García-Lara, J.; Needham, A.J.; Foster, S.J. Invertebrates as Animal Models for Staphylococcus Aureus Pathogenesis: A Window into Host–Pathogen Interaction. FEMS Immunol. Med. Microbiol. 2005, 43, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, S.; Matsumoto, Y.; Sekimizu, K.; Kaito, C. Evaluation of Staphylococcus Aureus Virulence Factors Using a Silkworm Model. FEMS Microbiol. Lett. 2012, 326, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, S.; Maculloch, B.; Batz, M. Economic Burden of Major Foodborne Illnesses Acquired in the United States, Eib-140; Economic Research Service U.S. Department of Agriculture: Washington, DC, USA, 2015. [Google Scholar]
- WHO. 2007–2015, Foodborne Disease Burden Epidemiology Reference Group. In Estimates of the Global Burden of Foodborne Diseases; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- World Health Organization. Global Incidence and Prevalence of Selected Curable Sexually Transmitted Infections-2008; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Simmering, J.E.; Tang, F.; Cavanaugh, J.E.; Polgreen, L.A.; Polgreen, P.M. The Increase in Hospitalizations for Urinary Tract Infections and the Associated Costs in the United States, 1998–2011. Open Forum Infect. Dis. 2017, 4, ofw281. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.Y.; Jiang, W.; Mojica, N.; Tseng, K.K.; McNeill, R.; Cosgrove, S.E.; Perl, T.M. National Costs Associated with Methicillin-Susceptible and Methicillin-Resistant Staphylococcus Aureus Hospitalizations in the United States, 2010–2014. Clin. Infect. Dis. 2019, 68, 22–28. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent Bacterial Infections and Persister Cells. Nat. Rev. Microbiol. 2017, 15, 453. [Google Scholar] [CrossRef] [PubMed]
- Ray, K.; Marteyn, B.; Sansonetti, P.J.; Tang, C.M. Life on the Inside: The Intracellular Lifestyle of Cytosolic Bacteria. Nat. Rev. Microbiol. 2009, 9, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Chai, Q.; Wang, L.; Liu, C.H.; Ge, B. New Insights into the Evasion of Host Innate Immunity by Mycobacterium Tuberculosis. Cell Mol. Immunol. 2020, 17, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Robino, L.; Scavone, P.; Araujo, L.; Algorta, G.; Zunino, P.; Pírez, M.C.; Vignoli, R. Intracellular Bacteria in the Pathogenesis of Escherichia Coli Urinary Tract Infection in Children. Clin. Infect. Dis. 2014, 75, e158–e164. [Google Scholar] [CrossRef] [Green Version]
- Freitag, N.E.; Port, G.C.; Miner, M.D. Listeria Monocytogenes—From Saprophyte to Intracellular Pathogen. Nat. Rev. Microbiol. 2009, 7, 623–628. [Google Scholar] [CrossRef]
- Nagamatsu, K.; Hannan, T.J.; Guest, R.L.; Kostakioti, M.; Hadjifrangiskou, M.; Binkley, J.; Dodson, K.; Raivio, T.L.; Hultgren, S.J. Dysregulation of Escherichia Coli A-Hemolysin Expression Alters the Course of Acute and Persistent Urinary Tract Infection. Proc. Natl. Acad. Sci. USA 2015, 75, E871–E880. [Google Scholar] [CrossRef] [Green Version]
- Carlson, J.H.; Whitmire, W.M.; Crane, D.D.; Wicke, L.; Virtaneva, K.; Sturdevant, D.E.; Kupko, J.J., 3rd; Porcella, S.F.; Martinez-Orengo, N.; Heinzen, R.A.; et al. The Chlamydia Trachomatis Plasmid Is a Transcriptional Regulator of Chromosomal Genes and a Virulence Factor. Infect. Immun. 2008, 76, 2273–2283. [Google Scholar] [CrossRef] [Green Version]
- Heuer, D.; Lipinski, A.R.; Machuy, N.; Karlas, A.; Wehrens, A.; Siedler, F.; Brinkmann, V.; Meyer, T.F. Chlamydia Causes Fragmentation of the Golgi Compartment to Ensure Reproduction. Nature 2009, 457, 731–735. [Google Scholar] [CrossRef]
- Tullius, M.V.; Harth, G.; Horwitz, M.A. Glutamine Synthetase Glna1 Is Essential for Growth of Mycobacterium Tuberculosis in Human Thp-1 Macrophages and Guinea Pigs. Infect. Immun. 2003, 71, 3927–3936. [Google Scholar] [CrossRef] [Green Version]
- Loisel-Meyer, S.; de Bagüés, M.P.J.; Bassères, E.; Dornand, J.; Köhler, S.; Liautard, J.-P.; Jubier-Maurin, V. Requirement of Nord for Brucella Suis Virulence in a Murine Model of in Vitro and in Vivo Infection. Infect. Immun. 2006, 75, 1973–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ammendola, S.; Pasquali, P.; Pistoia, C.; Petrucci, P.; Petrarca, P.; Rotilio, G.; Battistoni, A. High-Affinity Zn2+ Uptake System Znuabc Is Required for Bacterial Zinc Homeostasis in Intracellular Environments and Contributes to the Virulence of Salmonella Enterica. Infect. Immun. 2007, 75, 5867–5876. [Google Scholar] [CrossRef] [Green Version]
- Vitoriano, I.; Saraiva-Pava, K.D.; Rocha-Gonçalves, A.; Santos, A.; Lopes, A.I.; Oleastro, M.; Roxo-Rosa, M. Ulcerogenic Helicobacter Pylori Strains Isolated from Children: A Contribution to Get Insight into the Virulence of the Bacteria. PLoS ONE 2011, 6, e26265. [Google Scholar] [CrossRef] [Green Version]
- Mirrashidi, K.M.; Elwell, C.A.; Verschueren, E.; Johnson, J.R.; Frando, A.; von Dollen, J.; Rosenberg, O.; Gulbahce, N.; Jang, G.; Johnson, T.; et al. Global Mapping of the Inc-Human Interactome Reveals That Retromer Restricts Chlamydia Infection. Cell Host Microbe 2015, 18, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Purschke, F.G.; Hiller, E.; Trick, I.; Rupp, S. Flexible Survival Strategies of Pseudomonas Aeruginosa in Biofilms Result in Increased Fitness Compared with Candida Albicans. Mol. Cell. Proteom. MCP 2012, 12, 1652–1669. [Google Scholar] [CrossRef] [Green Version]
- Chey, W.D.; Leontiadis, G.I.; Howden, C.W.; Moss, S.F. Acg Clinical Guideline: Treatment of Helicobacter Pylori Infection. Am. J. Gastroenterol. 2017, 112, 212–239. [Google Scholar] [CrossRef]
- Metlay, J.P.; Waterer, G.W.; Long, A.C.; Anzueto, A.; Brozek, J.; Crothers, K.; Cooley, L.A.; Dean, N.C.; Fine, M.J.; Flanders, S.A.; et al. Diagnosis and Treatment of Adults with Community-Acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 2019, 200, e45–e67. [Google Scholar] [CrossRef]
- Fournier, P.E.; Thuny, F.; Richet, H.; Lepidi, H.; Casalta, J.P.; Arzouni, J.P.; Maurin, M.; Celard, M.; Mainardi, J.L.; Caus, T.; et al. Comprehensive Diagnostic Strategy for Blood Culture-Negative Endocarditis: A Prospective Study of 819 New Cases. Clin. Infect. Dis. 2010, 51, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Cyr, S.; Barbee, L.; Workowski, K.A.; Bachmann, L.H.; Pham, C.; Schlanger, K.; Torrone, E.; Weinstock, H.; Kersh, E.N.; Thorpe, P. Update to Cdc’s Treatment Guidelines for Gonococcal Infection, 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 50, 1911–1916. [Google Scholar] [CrossRef] [PubMed]
- Baddour, L.M.; Wilson, W.R.; Bayer, A.S.; Fowler, V.G., Jr.; Tleyjeh, I.M.; Rybak, M.J.; Barsic, B.; Lockhart, P.B.; Gewitz, M.H.; Levison, M.E.; et al. Infective Endocarditis in Adults: Diagnosis, Antimicrobial Therapy, and Management of Complications: A Scientific Statement for Healthcare Professionals from the American Heart Association. Circulation 2015, 15, 1435–1486. [Google Scholar]
- Osmon, D.R.; Berbari, E.F.; Berendt, A.R.; Lew, D.; Zimmerli, W.; Steckelberg, J.M.; Rao, N.; Hanssen, A.; Wilson, W.R. Diagnosis and Management of Prosthetic Joint Infection: Clinical Practice Guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, e1–e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tucker, A.N.; Carlson, T.J.; Sarkar, A. Challenges in Drug Discovery for Intracellular Bacteria. Pathogens 2021, 10, 1172. https://doi.org/10.3390/pathogens10091172
Tucker AN, Carlson TJ, Sarkar A. Challenges in Drug Discovery for Intracellular Bacteria. Pathogens. 2021; 10(9):1172. https://doi.org/10.3390/pathogens10091172
Chicago/Turabian StyleTucker, Allison N., Travis J. Carlson, and Aurijit Sarkar. 2021. "Challenges in Drug Discovery for Intracellular Bacteria" Pathogens 10, no. 9: 1172. https://doi.org/10.3390/pathogens10091172
APA StyleTucker, A. N., Carlson, T. J., & Sarkar, A. (2021). Challenges in Drug Discovery for Intracellular Bacteria. Pathogens, 10(9), 1172. https://doi.org/10.3390/pathogens10091172