
A Molecular Survey on Neglected Gurltia paralysans and Aelurostrongylus abstrusus Infections in Domestic Cats (Felis catus) from Southern Chile


 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
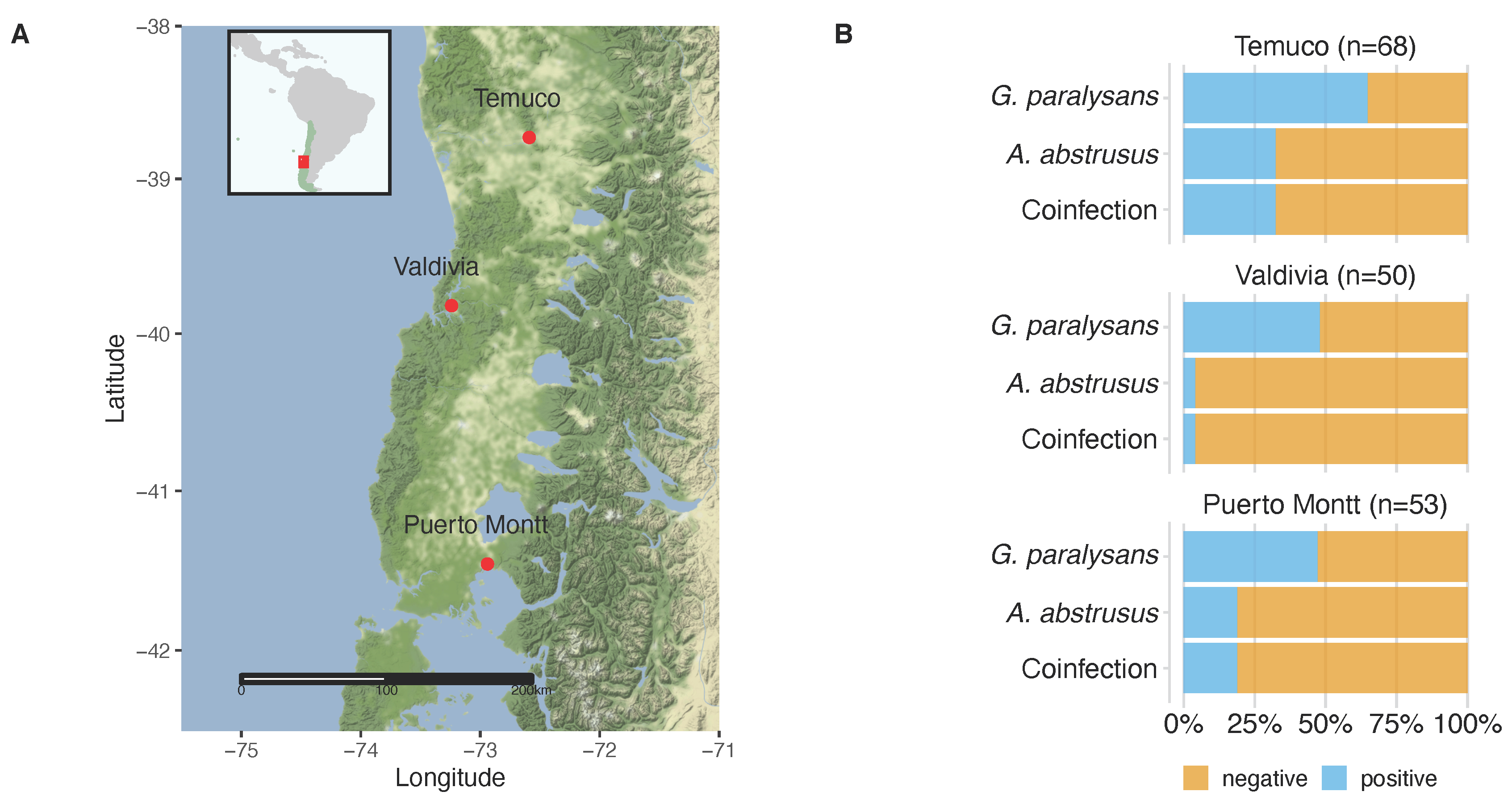
2.1. Animals
2.2. Descriptive Epidemiology
2.3. Epidemiological Factors
2.4. Molecular Detection of G. paralysans and A. abstrusus
3. Discussion
4. Materials and Methods
4.1. Animals and Study Area
4.2. Selection of Participants and Biological Samples
4.3. Extraction and Quantification of G. paralysans and A. abstrusus–Specific DNA
4.4. Amplification of IRBP as an Internal Control by Conventional PCR
4.5. Sequencing Analysis
4.6. Questionnaire for Domestic Cat Owners
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez, M.; Moroni, M.; Muñoz, P.; Taubert, A.; Hermosilla, C.; Hirzmann, J.; Rojas, L. Gurltia paralysans: A neglected parasite of domestic cats. Austral J. Vet. Sci. 2021, 52, 33–45. [Google Scholar] [CrossRef]
- Muñoz, P.; Hirzmann, J.; Rodriguez, E.; Moroni, M.; Taubert, A.; Gibbons, L.; Hermosilla, C.; Gómez, M. Redescription and first molecular characterization of the little known feline neurotropic nematode Gurltia paralysans (Nematoda: Metastrongyloidea). Vet. Parasitol. Reg. Stud. Rep. 2017, 10, 119–125. [Google Scholar]
- Moroni, M.; Muñoz, P.; Gómez, M.; Mieres, M.; Rojas, M.; Lillo, C.; Aguirre, F.; Acosta-Jamett, G.; Kaiser, M.; Lindsay, D. Gurltia paralysans (Wolffhügel, 1933): Description of adults and additional case reports of neurological diseases in three domestic cats from southern Chile. Vet. Parasitol. 2012, 184, 377–380. [Google Scholar] [CrossRef]
- Moroni, M.; Muñoz, P.; Mieres, M.; Gómez, M.; Vera, F. Severe spinal cord thrombophlebitis and meningomyelitis by Gurltia paralysans in a cat. Vet. Rec. Case Rep. 2016, 4, e000327. [Google Scholar] [CrossRef]
- Togni, M.; Panziera, W.; Souza, T.; Oliveira, J.; Mazzanti, A.; Barros, C.; Fighera, R. Epidemiological, clinical and pathological aspects of Gurltia paralysans infection in cats. Pesqui. Veterinária Bras. 2013, 33, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Gómez, M.; Mieres, M.; Moroni, M.; Mora, A.; Barrios, N.; Simeone, C.; Lindsay, D. Meningomyelitis due to nematode infection in four cats. Vet. Parasitol. 2010, 170, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, P.; Gómez, M.; Moroni, M.; Muñoz, P.; Müller, A. Evaluation of terrestrial gastropods as possible intermediate hosts of Gurltia paralysans in southern Chile. Rev. Bras. Parasitol. Vet. 2021, 30, e025020. [Google Scholar] [CrossRef]
- Mieres, M.; Gómez, M.; Lillo, C.; Rojas, M.; Moroni, M.; Muñoz, P.; Acosta-Jamett, G.; Wiegand, R. Clinical, imaging and pathologic characteristics of Gurltia paralysans myelopathy in domestic cats from Chile. Vet. Radiol. Ultrasound 2013, 54, 237–244. [Google Scholar] [CrossRef]
- López-Contreras, F.; Rojas-Barón, L.; Gómez, M.; Morera, F.; Sepúlveda, P.; Moroni, M.; Muñoz, P.; Acosta-Jammett, G.; Mieres, M.; Hirzmann, J.; et al. Molecular Detection of Gurltia paralysans by Semi-Nested PCR in Cerebrospinal Fluid and Serum Samples from Domestic Cats (Felis catus). Animals 2020, 10, 1169. [Google Scholar] [CrossRef]
- Alzate, G.; Aranzazu, D.; Alzate, A.; Chaparro, J. Domestic cat paraplegia compatible with Gurltia paralysans nematode. First cases reported in Colombia. Rev. Colomb. Cienc. Pecu. 2011, 24, 663–669. [Google Scholar]
- Rivero, R.; Matto, C.; Adrien, M.; Nan, F.; Bell, T.; Gardiner, C. Parasite meningomyelitis in cats in Uruguay. Rev. Bras. Parasitol. Vet. 2011, 20, 259–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bono, M.F.; Orcellet, V.; Marengo, R.; Bosio, A.; Junkers, E.; Plaza, D.; Marini, M.R.; Sanchez, A.; Rubio, M.G.; Candiotti, V. A description of three cases of parasitic meningomyelitis in felines from the province of Santa Fé, Argentina. Parasitaria 2016, 74, 1–4. [Google Scholar]
- Udiz-Rodríguez, R.; García-Livia, K.; Valladares-Salmerón, M.; Dorta-Almenar, M.; Martín-Carrillo, N.; Martin-Alonso, A.; Isquierdo-Rodriguez, E.; Feliu, C.; Valladares, B.; Foronda, P. First ocular report of Gurltia paralysans (Wolffhügel, 1993) in cat. Vet. Parasitol. 2018, 255, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; Guglielmini, C. Feline aelurostrongylosis and canine angiostrongylosis: A challenging diagnosis for two emerging verminous pneumonia infections. Vet. Parasitol. 2008, 157, 163–174. [Google Scholar] [CrossRef]
- Traversa, D.; Morelli, S.; Di Cesare, A.; Diakou, A. Felid Cardiopulmonary Nematodes: Dilemmas Solved and New Questions Posed. Pathogens 2021, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Elsheikha, H.; Schnyder, M.; Traversa, D.; Di Cesare, A.; Wright, I.; Lacher, D. Updates on feline aelurostrongylosis and research priorities for de next decade. Parasites Vectors 2016, 9, 389. [Google Scholar] [CrossRef] [Green Version]
- Penagos-Tabares, F.; Lange, M.K.; Vélez, J.; Hirzmann, J.; Gutiérrez-Arboleda, J.; Taubert, A.; Hermosilla, C.; Chaparro Gutiérrez, J.J. The invasive giant African snail Lissachatina fulica as natural intermediate host of Aelurostrongylus abstrusus, Angiostrongylus vasorum, Troglostrongylus brevior, and Crenosoma vulpis in Colombia. PLoS Negl. Trop. Dis. 2019, 13, e0007277. [Google Scholar] [CrossRef]
- Dimzas, D.; Morelli, S.; Traversa, D.; Di Cesare, A.; Van Bourgonie, Y.; Breugelmans, K.; Backeljau, T.; di Regalbono, A.F.; Diakou, A. Intermediate gastropod hosts of major feline cardiopulmonary nematodes in an area of wildcat and domestic cat sympatry in Greece. Parasites Vectors 2020, 13, 345. [Google Scholar] [CrossRef]
- Morelli, S.; Colombo, M.; Diakou, A.; Traversa, D.; Grillini, M.; Frangipane di Regalbono, A.; Di Cesare, A. The Influence of Temperature on the Larval Development of Aelurostrongylus abstrusus in the Land Snail Cornu aspersum. Pathogens 2021, 10, 960. [Google Scholar] [CrossRef]
- Fuehrer, H.P.; Morelli, S.; Bleicher, J.; Brauchart, T.; Edler, M.; Eisschiel, N.; Hering, T.; Lercher, S.; Mohab, K.; Reinelt, S.; et al. Detection of Crenosoma spp., Angiostrongylus vasorum and Aelurostrongylus abstrusus in Gastropods in Eastern Austria. Pathogens 2020, 9, 1046. [Google Scholar] [CrossRef]
- Jezewski, W.; Bunkowska-Gawlik, K.; Hildebrand, J.; Perec-Matysiak, A.; Laskowski, Z. Intermediate and paratenic hosts in the life cycle of Aelurostrongylus abstrusus in natural environment. Vet. Parasitol. 2013, 198, 401–405. [Google Scholar] [CrossRef]
- Morelli, S.; Diakou, A.; Colombo, M.; Di Cesare, A.; Barlaam, A.; Dimzas, D.; Traversa, D. Cat Respiratory Nematodes: Current Knowledge, Novel Data and Warranted Studies on Clinical Features, Treatment and Control. Pathogens 2021, 10, 454. [Google Scholar] [CrossRef]
- Moskvina, T.V. Current knowledge about Aelurostrongylus abstrusus biology and diagnostic. Ann. Parasitol. 2018, 64, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Raue, K.; Raue, J.; Hauck, D.; Söbbeler, F.; Morelli, S.; Traversa, D.; Schnyder, M.; Volk, H.; Strube, C. Do All Roads Lead to Rome? The Potential of Different Approaches to Diagnose Aelurostrongylus abstrusus Infection in Cats. Pathogens 2021, 14, 602. [Google Scholar] [CrossRef]
- Annoscia, G.; Latrofa, M.S.; Campbell, B.E.; Giannelli, A.; Ramos, R.A.; Dantas-Torres, F.; Brianti, E.; Otranto, D. Simultaneous detection of the feline lungworms Troglostrongylus brevior and Aelurostrongylus abstrusus by a newly developed duplex-PCR. Vet. Parasitol. 2014, 199, 172–178. [Google Scholar] [CrossRef]
- daSilvaLima, W.; Ferreira Farago, E.C.; do Nascimento Mesquita, M.; Duarte Pacheco, A.; da Silva Malavazi, P.F.N.; Salvador Oliveira, H.; Morelli, S.; Colombo, M.; Di Cesare, A.; de Souza, S.F. First Case of Clinical Cat Aelurostrongylosis in the Brazilian Amazon: Clinical and Molecular Insights. Pathogens 2021, 10, 595. [Google Scholar] [CrossRef] [PubMed]
- Briggs, K.R.; Yaros, J.P.; Liotta, J.L.; Lucio-Forster, A.; Lee, A.C.; Bowman, D.D. Detecting Aelurostrongylus abstrusus-specific IgG antibody using an immunofluorescence assay. J. Feline Med. Surg. 2013, 15, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Osorio, S.; Navarro-Ruiz, J.L.; Rave, A.; Taubert, A.; Hermosilla, C.; Chaparro-Gutierrez, J.J. Aelurostrongylus abstrusus Infections in Domestic Cats (Felis silvestris catus) from Antioquia, Colombia. Pathogens 2021, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Penagos-Tabares, F.; Lange, M.K.; Chaparro-Gutiérrez, J.J.; Taubert, A.; Hermosilla, C. Angiostrongylus vasorum and Aelurostrongylus abstrusus: Neglected and underestimated parasites in South America. Parasites Vectors 2018, 11, 208. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, I.; Paludi, A.; Saumell, L. Primera Descripción en Argentina de Gurltia Paralysans en un Felino Doméstico. Bachelor’s Thesis, Universidad Nacional del Centro de la Provincia de Buenas Aires, Tandil, Argentina, 2011. Available online: http://www.biblioteca.unlpam.edu.ar/pubpdf/revet/v18n2a05pellegrino.pdf (accessed on 1 June 2021).
- Melo Neto, G.L.; da Silva, R.; Alves, R.C.; Gomes, R.; Medeiros, A.; Araújo, M. Infecção por Gurltia paralysans em gatos domésticos no Eestado de Pernambuco, Brasil. Acta Sci. Vet. 2019, 47, 418. [Google Scholar]
- Maksimov, P.; Hermosilla, C.; Taubert, A.; Staubach, C.; Sauter-Louis, C.; Conraths, F.J.; Vrhovec, M.G.; Pantchev, N. GIS-supported epidemiological analysis on canine Angiostrongylus vasorum and Crenosoma vulpis infections in Germany. Parasites Vectors 2017, 10, 108. [Google Scholar] [CrossRef] [Green Version]
- Segeritz, L.; Cardona, A.; Taubert, A.; Hermosilla, C.; Ruiz, A. Autochthonous Angiostrongylus cantonensis, Angiostrongylus vasorum and Aelurostrongylus abstrusus infections in native terrestrial gastropods from the Macaronesian Archipelago of Spain. Parasitol. Res. 2021, 20, 2671–2680. [Google Scholar] [CrossRef]
- Traversa, D.; Morelli, S.; Cassini, R.; Crisi, P.E.; Russi, I.; Grillotti, E.; Manzocchi, S.; Simonato, G.; Beraldo, P.; Viglietti, A.; et al. Occurrence of canine and feline extra-intestinal nematodes in key endemic regions of Italy. Acta Trop. 2019, 193, 227–235. [Google Scholar] [CrossRef]
- Morelli, S.; Diakou, A.; Di Cesare, A.; Schnyder, M.; Colombo, M.; Strube, C.; Dimzas, D.; Latino, R.; Traversa, D. Feline lungworms in Greece: Copromicroscopic, molecular and serological study. Parasitol. Res. 2020, 119, 2877–2883. [Google Scholar] [CrossRef]
- Györke, A.; Dumitrache, M.O.; Kalmár, Z.; Paştiu, A.I.; Mircean, V. Molecular survey of metastrongyloid lungworms in domestic cats (Felis silvestris catus) from Romania: A retrospective study (2008–2011). Pathogens 2020, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zottler, E.M.; Bieri, M.; Basso, W.; Schnyder, M. Intestinal parasites and lungworms in stray, shelter and privately owned cats of Switzerland. Parasitol. Int. 2019, 69, 75–81. [Google Scholar] [CrossRef]
- Escobar, R.; Illanes, O.; Fuentealba, I.; Cubillos, V. Nematodiasis pulmonar en el gato doméstico. Arch. Med. Vet. 1984, 16, 47–49. [Google Scholar]
- Oyarzún-Cadagán, J.A. Pesquisa de nematodos pulmonares en perros y gatos de las ciudades de Río Bueno y La Unión, Provincia del Ranco, Chile: Universidad Austral de Chile Valdivia. 2013. Available online: http://cybertesis.uach.cl/tesis/uach/2013/fvo.98p/doc/fvo.98p.pdf (accessed on 8 March 2018).
- Trein, E.J. Lesões produzidas por Aelurostrongylus abstrusus (Railliet, 1898) no pulmão do gato doméstico. In Tese Para Inscrição ao Concurso Para a Cadeira de Anatomia Patológica e Técnica de Necropsia; Universidade do Rio Grande do Sul: Porto Alegre, Brazil, 1953. [Google Scholar]
- Mundim, T.; Oliveira Júnior, S.; Rodrigues, D.; Cury, M. Frequency of helminths parasites in cats of Uberlândia, Minas Gerais. Arq. Bras. Med. Vet. Zoo. 2004, 56, 562–563. [Google Scholar] [CrossRef] [Green Version]
- Headley, S.A. Aelurostrongylus abstrusus induced pneumonia in cats: Pathological and epidemiological findings of 38 cases (1987-1996). Semin. Ciências Agrárias 2005, 26, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Ramos, D.G.D.S.; Scheremeta, R.G.A.D.C.; De Oliveira, A.C.S.; Sinkoc, A.L.; Pacheco, R.D.C. Survey of helminth parasites of cats from the metropolitan area of Cuiabá, Mato Grosso, Brazil. Rev. Bras. Parasitol. Vet. 2013, 22, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusma, S.C.; Wrublewski, D.M.; Teixeira, V.N.; Holdefer, D.R. Parasitos intestinais de Leopardus wiedii e Leopardus tigrinus (Felidae) da Floresta Nacional De Três Barras, sc. Luminária 2015, 17, 82–95. [Google Scholar]
- Oliveira, B. Politraumatismo em gato-maracajá (Leopardus wieddi) com infecção por Gurltia paralysans. In Integrando Prácticas e Transvesalizando Saberes; Semana do Conhecimiento UPF: Passo Fundo, Brazil, 2015. [Google Scholar]
- Dazzi, C.C.; Santos, A.; Machado, T.P.; Ataíde, M.W.; Rodriguez, R.; Pereira, A.M.; García, P.S.; Da Motta, A.C. First case report of nematode parasitic myelopathy in a wild feline in Brazil. Braz. J. Vet. Parasitol. 2020, 29, e014619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, O.; Valledor, S.; Crampet, A.; Casás, G. Aporte al conocimiento de los metazoos parásitos del gato doméstico en el Departamento de Montevideo, Uruguay. Veterinaria 2013, 49, 28–37. [Google Scholar]
- Idiart, J.; Martín, A.; Venturini, L.; Ruager, J. Neumonía por Aelurostrongylus abstrusus en gatos. Primeros hallazgos en el gran Buenos Aires y La Plata. Veterinaria Argent. 1986, 23, 229–237. [Google Scholar]
- Martínez, A.; Santa Cruz, A.; Lombardero, O. Histopathological lesions in feline aelurostrongylosis. Rev. Med. Vet. 1990, 71, 260–264. [Google Scholar]
- Cardillo, N.; Clemente, A.; Pasqualetti, M.; Borrás, P.; Rosa, A.; Ribicich, M. First report of Aelurostrongylus abstrusus in domestic land snail Rumina decollata, in the Autonomous city of Buenos Aires. InVet 2014, 16, 15–22. [Google Scholar]
- Barutzki, D.; Schaper, R. Results of parasitological examinations of faecal samples from cats and dogs in Germany between 2003 and 2010. Parasitol. Res. 2011, 109, 45–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferies, R.; Vrhovec, M.G.; Wallner, N.; Catalan, D.R. Aelurostrongylus abstrusus and Troglostrongylus sp. (Nematoda: Metastrongyloidea) infections in cats inhabiting Ibiza, Spain. Vet. Parasitol. 2010, 173, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Gavrilovic, P.; Jovanovic, M.; Gavrilovic, A.; Nesic, S. Fatal aelurostrongylosis in a kitten in Serbia. Acta Parasitol. 2017, 62, 488–491. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.P.; Skarbye, L.K.; Vinther, L.M.; Willesen, J.L.; Pipper, C.B.; Olsen, C.S.; Mejer, H. Occurrence and clinical significance of Aelurostrongylus abstrusus and other endoparasites in Danish cats. Vet. Parasitol. 2017, 234, 31–39. [Google Scholar] [CrossRef]
- Diakou, A.; Di Cesare, A.; Barros, L.A.; Morelli, S.; Halos, L.; Beugnet, F.; Traversa, D. Occurrence of Aelurostrongylus abstrusus and Troglostrongylus brevior in domestic cats in Greece. Parasites Vectors 2015, 8, 590. [Google Scholar] [CrossRef] [Green Version]
- Willard, M.D.; Roberts, R.E.; Allison, N.; Grieve, R.B.; Escher, K. Diagnosis of Aelurostrongylus abstrusus and Dirofilaria immitis infec- tions in cats from a humane shelter. J. Am. Vet. Med. Assoc. 1988, 192, 913–916. [Google Scholar] [PubMed]
- Carruth, A.J.; Buch, J.S.; Braff, J.C.; Chandrashekar, R.; Bowman, D.D. Distribution of the feline lungworm Aelurostrongylus abstrusus in the USA based on fecal testing. J. Feline Med. Surg. Open Rep. 2019, 5. [Google Scholar] [CrossRef]
- Nagamori, Y.; Payton, M.E.; Looper, E.; Apple, H.; Johnson, E.M. Retrospective survey of parasitism identified in feces of client-owned cats in North America from 2007 through 2018. Vet. Parasitol. 2020, 277, 109008. [Google Scholar] [CrossRef]
- Traversa, D.; Iorio, R.; Otranto, D. Diagnostic and clinical implications of a nested PCR specific for ribosomal DNA of the feline lungworm Aelurostrongylus abstrusus (Nematoda, Strongylida). J. Clin. Microbiol. 2008, 46, 1811–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traversa, D.; Di Cesare, A.; Milillo, P.; Iorio, R.; Otranto, D. Aelurostrongylus abstrusus in a feline colony from central Italy: Clinical features, diagnostic procedures and molecular characterization. Parasitol. Res. 2008, 103, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Aguirre, S.; Alegría-Morán, R.A.; Calderón-Amor, J.; Tadich, T.A. Can Responsible Ownership Practices Influence Hunting Behavior of Owned Cats? Results from a Survey of Cat Owners in Chile. Animals 2019, 9, 745. [Google Scholar] [CrossRef] [Green Version]
- Falsone, L.; Colella, V.; Napoli, E.; Brianti, E.; Otranto, D. The cockroach Periplaneta americana as a potential paratenic host of the lungworm Aelurostrongylus abstrusus. Exp. Parasitol. 2017, 182, 54–57. [Google Scholar] [CrossRef]
- Di Cesare, A.; Frangipane di Regalbono, A.; Tessarin, C.; Seghetti, M.; Iorio, R.; Simonato, G.; Traversa, D. Mixed infection by Aelurostrongylus abstrusus and Troglostrongylus brevior in kittens from the same litter in Italy. Parasitol. Res. 2014, 113, 613–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cesare, A.; Veronesi, F.; Grillotti, E.; Manzocchi, S.; Perrucci, S.; Beraldo, P.; Cazzin, S.; De Liberato, C.; Barros, L.A.; Simonato, G.; et al. Respiratory nematodes in cat populations of Italy. Parasitol. Res. 2015, 114, 4463–4469. [Google Scholar] [CrossRef] [Green Version]
- Genchi, M.; Ferrari, N.; Fonti, P.; De Francesco, I.; Piazza, C.; Viglietti, A. Relation between Aelurostrongylus abstrusus larvae excretion, respiratory and radiographic signs in naturally infected cats. Vet. Parasitol. 2014, 206, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Elsheikha, H.M.; Wright, I.; Wang, B.; Schaper, R. Prevalence of feline lungworm Aelurostrongylus abstrusus in England. Vet. Parasitol. Reg. Stud. Rep. 2019, 16, 100271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, E.C.; Gontijo, C.M.; Cruz, I.; Melo, M.N.; Silva, A.M. Alternative PCR protocol using a single primer set for assessing DNA quality in several tissues from a large variety of mammalian species living in áreas endemic for leishmaniasis. Mem. Inst. Oswaldo Cruz. 2010, 105, 895–898. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| G. paralysans | A. abstrusus | A. abstrusus + G. paralysans | ||||
|---|---|---|---|---|---|---|
| City | Positive/Total | % | Positive/Total | % | Positive/Total | % |
| Puerto Montt | 25/53 | 47.2 | 10/53 | 18.9 | 10/53 | 18.9 |
| Temuco | 44/68 | 64.7 | 22/68 | 32.4 | 22/68 | 32.4 |
| Valdivia | 24/50 | 48 | 2/50 | 4.0 | 2/50 | 4.0 |
| Total | 93/171 | 54.4 | 34/171 | 19.9 | 34/171 | 19.9 |
| G. paralysans | A. abstrusus | |||
|---|---|---|---|---|
| Positive/Total | % | Positive/Total | % | |
| Gender | ||||
| female | 41/77 | 46.8 | 15/77 | 19.5 |
| male | 37/69 | 46.4 | 19/69 | 27.5 |
| n/d | 15/25 | 40.0 | ||
| Total | 93/171 | 45.6 | 34/146 | 23.3 |
| Indoor/outdoor | ||||
| indoor | 12/28 | 57.1 | 14/64 | 21.9 |
| outdoor | 33/64 | 48.4 | 7/8 | 25.0 |
| n/d | 48/79 | 39.2 | 13/79 | 16.5 |
| Total | 93/171 | 45.6 | 34/171 | 19.9 |
| Anthelmintic | ||||
| no | 23/55 | 58.2 | 8/55 | 14.5 |
| yes | 23/38 | 39.5 | 13/38 | 34.2 |
| n/d | 47/78 | 39.7 | 13/78 | 16.7 |
| Total | 93/171 | 45.6 | 34/171 | 19.9 |
| Hunting | ||||
| no | 22/50 | 56.0 | 13/50 | 19.0 |
| yes | 23/42 | 45.2 | 8/42 | 26.0 |
| n/d | 48/79 | 39.2 | 13/79 | 16.5 |
| Total | 93/171 | 45.6 | 34/171 | 19.9 |
| Age | ||||
| <1 year | 24/43 | 44.2 | 8/43 | 18.6 |
| ≥1 year | 49/95 | 48.4 | 23/95 | 24.2 |
| n/d | 20/33 | 39.4 | 3/33 | 9.1 |
| Total | 93/171 | 45.6 | 34/171 | 19.9 |
| A. abstrusus | |||
|---|---|---|---|
| Predictors | Odds Ratios | CI | p |
| age [young] | 1.24 | 0.36–4.08 | 0.72 |
| sex [male] | 1.01 | 0.35–2.91 | 0.98 |
| lifestyle [outdoor] | 1.88 | 0.47–8.03 | 0.37 |
| anthelmintic [yes] | 3.92 | 1.33–12.77 | 0.02 |
| hunting [yes] | 0.40 | 0.10–1.59 | 0.19 |
| G. paralysans | |||
|---|---|---|---|
| Predictors | Odds Ratios | CI | p |
| age [young] | 1.55 | 0.57–4.29 | 0.39 |
| sex [male] | 1.28 | 0.53–3.10 | 0.58 |
| lifestyle [outdoor] | 1.64 | 0.52–5.39 | 0.40 |
| anthelmintic [yes] | 2.36 | 0.96–6.03 | 0.06 |
| hunting [yes] | 1.02 | 0.34–3.07 | 0.96 |
| Oligonucleotide | Target | Primer Name | Sequence |
|---|---|---|---|
| Universal | Metastrongyloidea | AaGp28Ss1 | 5’-CGAGTRATATGTATGCCATT-3´ |
| Universal | Metastrongyloidea | AaGp28Sa1 | 5´-AGGCATAGTTCACCATCT-3´ |
| Specific | Gurltia paralysans | Gp28Sa3 | 5´-TCTTGCCGCCATTATAGTAG-3´ |
| Specific | Aelurostrongylus abstrusus | Aa28Ss2 | 5´-CGTTGATGTTGATGAGTATC-3´ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrios, N.; Gómez, M.; Zanelli, M.; Rojas-Barón, L.; Sepúlveda-García, P.; Alabí, A.; Adasme, M.; Müller, A.; Rosenfeld, C.; González-Lagos, C.; et al. A Molecular Survey on Neglected Gurltia paralysans and Aelurostrongylus abstrusus Infections in Domestic Cats (Felis catus) from Southern Chile. Pathogens 2021, 10, 1195. https://doi.org/10.3390/pathogens10091195
Barrios N, Gómez M, Zanelli M, Rojas-Barón L, Sepúlveda-García P, Alabí A, Adasme M, Müller A, Rosenfeld C, González-Lagos C, et al. A Molecular Survey on Neglected Gurltia paralysans and Aelurostrongylus abstrusus Infections in Domestic Cats (Felis catus) from Southern Chile. Pathogens. 2021; 10(9):1195. https://doi.org/10.3390/pathogens10091195
Chicago/Turabian StyleBarrios, Natasha, Marcelo Gómez, Macarena Zanelli, Lisbeth Rojas-Barón, Paulina Sepúlveda-García, Amir Alabí, Melany Adasme, Ananda Müller, Carla Rosenfeld, César González-Lagos, and et al. 2021. "A Molecular Survey on Neglected Gurltia paralysans and Aelurostrongylus abstrusus Infections in Domestic Cats (Felis catus) from Southern Chile" Pathogens 10, no. 9: 1195. https://doi.org/10.3390/pathogens10091195
APA StyleBarrios, N., Gómez, M., Zanelli, M., Rojas-Barón, L., Sepúlveda-García, P., Alabí, A., Adasme, M., Müller, A., Rosenfeld, C., González-Lagos, C., Taubert, A., & Hermosilla, C. (2021). A Molecular Survey on Neglected Gurltia paralysans and Aelurostrongylus abstrusus Infections in Domestic Cats (Felis catus) from Southern Chile. Pathogens, 10(9), 1195. https://doi.org/10.3390/pathogens10091195