
Distinct Schistosoma mansoni-Specific Immunoglobulin Subclasses Are Induced by Different Schistosoma mansoni Stages—A Tool to Decipher Schistosoma mansoni Infection Stages

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
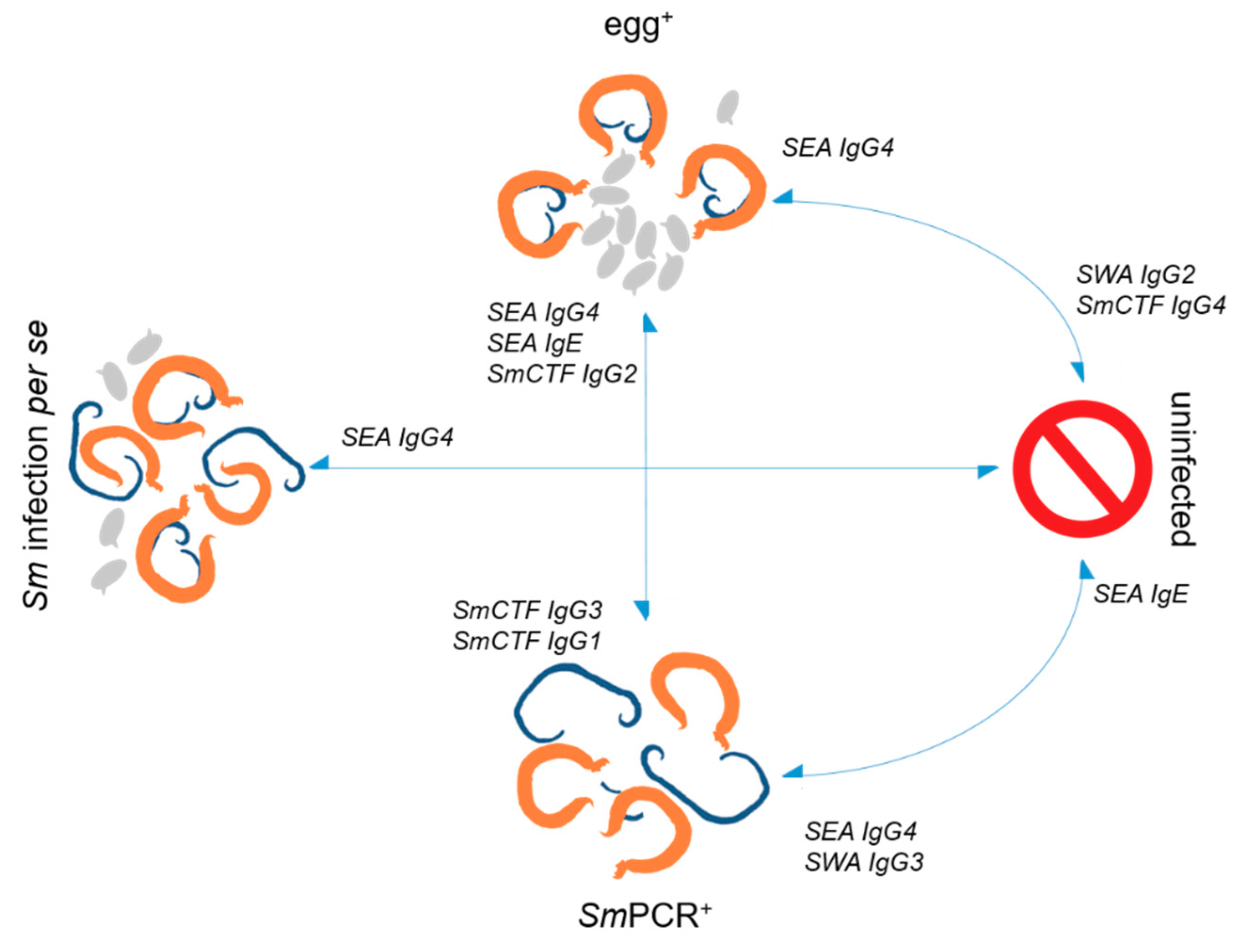
2.1. All Measured SmCTF-specific IgGs Are Elevated in SmPCR+ Individuals
2.2. SmCTF IgG1 Levels Are Associated with SmPCR+ Individuals
2.3. Strong Association of SEA-Specific IgG4 with Infection
3. Discussion
4. Methods
4.1. Epidemiological Profiling and Serum Collection
4.2. Schistosome-Specific Diagnostics and Groupings
4.3. Determination of SmCTF Immunoglobulins
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cha, S.; Elhag, M.S.; Lee, Y.H.; Cho, D.S.; Ismail, H.; Hong, S.T. Epidemiological findings and policy implications from the nationwide schistosomiasis and intestinal helminthiasis survey in sudan. Parasit Vectors 2019, 12, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfaki, T.E.; Arndts, K.; Wiszniewsky, A.; Ritter, M.; Goreish, I.A.; Atti El Mekki, M.E.; Arriens, S.; Pfarr, K.; Fimmers, R.; Doenhoff, M.; et al. Multivariable regression analysis in Schistosoma mansoni-infected individuals in the sudan reveals unique immunoepidemiological profiles in uninfected, egg+ and non-egg+ infected individuals. PLoS Negl. Trop. Dis. 2016, 10, e0004629. [Google Scholar] [CrossRef] [PubMed]
- Arndts, K.; Elfaki, T.E.M.; Jelden, N.; Ritter, M.; Wiszniewsky, A.; Katawa, G.; Goreish, I.A.; El Mekki, M.; Doenhoff, M.J.; Hoerauf, A.; et al. Schistosoma mansoni pcr+ infected individuals in the sudan present elevated systemic levels of chemokines when compared to uninfected and egg+ cohorts. Clin. Exp. Immunol. 2019, 196, 364–373. [Google Scholar] [CrossRef]
- WHO. Sudan Intensifies Its Fight against Schistosomiasis through Large-Scale Treatment of School-Age Children. Available online: https://www.who.int/news/item/02-09-2013-sudan-intensifies-its-fight-against-schistosomiasis-through-large-scale-treatment-of-school-age-children (accessed on 20 December 2021).
- Charani, E.; Cunnington, A.J.; Yousif, A.H.A.; Seed Ahmed, M.; Ahmed, A.E.M.; Babiker, S.; Badri, S.; Buytaert, W.; Crawford, M.A.; Elbashir, M.I.; et al. In transition: Current health challenges and priorities in sudan. BMJ Glob. Health 2019, 4, e001723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhag, M.S.; Jin, Y.; Amin, M.A.; Ismail, H.; Hong, S.T.; Jang, H.I.; Doh, Y.; Cha, S. Cost and logistics implications of a nationwide survey of schistosomiasis and other intestinal helminthiases in sudan: Key activities and cost components. PLoS ONE 2020, 15, e0226586. [Google Scholar] [CrossRef]
- Vale, N.; Gouveia, M.J.; Rinaldi, G.; Brindley, P.J.; Gartner, F.; Correia da Costa, J.M. Praziquantel for schistosomiasis: Single-drug metabolism revisited, mode of action, and resistance. Antimicrob. Agents Chemother 2017, 61, e02582-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergquist, R.; Utzinger, J.; Keiser, J. Controlling schistosomiasis with praziquantel: How much longer without a viable alternative? Infect. Dis. Poverty 2017, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.N. Schistosomiasis. Nat. Rev. Dis. Primers 2018, 4, 13. [Google Scholar] [CrossRef]
- Dunn, D.; Vennerwald, B. Schistosomiasis. In Oxford Textbook of Medicine, 6th ed.; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Grimes, J.E.; Croll, D.; Harrison, W.E.; Utzinger, J.; Freeman, M.C.; Templeton, M.R. The relationship between water, sanitation and schistosomiasis: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2014, 8, e3296. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.; Doenhoff, M.; Aitken, C.; Bailey, W.; Ji, M.; Dawson, E.; Gilis, H.; Spence, G.; Alexander, C.; van Gool, T. Comparison of Schistosoma mansoni soluble cercarial antigens and soluble egg antigens for serodiagnosing schistosome infections. PLoS Negl. Trop. Dis. 2012, 6, e1815. [Google Scholar] [CrossRef]
- Egesa, M.; Lubyayi, L.; Tukahebwa, E.M.; Bagaya, B.S.; Chalmers, I.W.; Wilson, S.; Hokke, C.H.; Hoffmann, K.F.; Dunne, D.W.; Yazdanbakhsh, M.; et al. Schistosoma mansoni schistosomula antigens induce th1/pro-inflammatory cytokine responses. Parasite Immunol. 2018, 40, e12592. [Google Scholar] [CrossRef]
- Hayunga, E.G.; Mollegard, I.; Duncan, J.F., Jr.; Sumner, M.P.; Stek, M., Jr.; Hunter, K.W., Jr. Early diagnosis of Schistosoma mansoni in mice using assays directed against cercarial antigens isolated by hydrophobic chromatography. J. Parasitol. 1987, 73, 351–362. [Google Scholar] [CrossRef]
- Kanamura, H.Y.; Hoshino-Shimizu, S.; da Silva, L.C. Schistosoma mansoni cercaria and schistosomulum antigens in serodiagnosis of schistosomiasis. Bull. Pan. Am. Health Organ. 1992, 26, 220–229. [Google Scholar]
- Winkel, B.M.F.; Dalenberg, M.R.; de Korne, C.M.; Feijt, C.; Langenberg, M.C.C.; Pelgrom, L.; Ganesh, M.S.; Yazdanbakhsh, M.; Smits, H.H.; de Jong, E.C.; et al. Early induction of human regulatory dermal antigen presenting cells by skin-penetrating Schistosoma mansoni cercariae. Front. Immunol. 2018, 9, 2510. [Google Scholar] [CrossRef] [PubMed]
- Anisuzzaman, S.F.; Prodjinotho, U.F.; Bhattacharjee, S.; Verschoor, A.; Prazeres da Costa, C. Host-specific serum factors control the development and survival of Schistosoma mansoni. Front. Immunol. 2021, 12, 635622. [Google Scholar] [CrossRef] [PubMed]
- Hagan, P.; Blumenthal, U.J.; Dunn, D.; Simpson, A.J.; Wilkins, H.A. Human ige, igg4 and resistance to reinfection with Schistosoma haematobium. Nature 1991, 349, 243–245. [Google Scholar] [CrossRef]
- Dunne, D.W.; Butterworth, A.E.; Fulford, A.J.; Kariuki, H.C.; Langley, J.G.; Ouma, J.H.; Capron, A.; Pierce, R.J.; Sturrock, R.F. Immunity after treatment of human schistosomiasis: Association between ige antibodies to adult worm antigens and resistance to reinfection. Eur. J. Immunol. 1992, 22, 1483–1494. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Abubaker, H. Control of schistosomiasis in the gezira irrigation scheme, sudan. J. Biosoc. Sci. 2017, 49, 83–98. [Google Scholar] [CrossRef]
- Sokolow, S. The History of Schistosomiasis in Sudan. Available online: https://schisto.stanford.edu/pdf/Sudan%20and%20South%20Sudan.pdf (accessed on 20 December 2021).
- Fukushige, M.; Mutapi, F.; Woolhouse, M.E.J. Population level changes in schistosome-specific antibody levels following chemotherapy. Parasite Immunol. 2019, 41, e12604. [Google Scholar] [CrossRef] [PubMed]
- Chand, M.A.; Chiodini, P.L.; Doenhoff, M.J. Development of a new assay for the diagnosis of schistosomiasis, using cercarial antigens. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 255–258. [Google Scholar] [CrossRef]
- Katabarwa, M.N.; Eyamba, A.; Nwane, P.; Enyong, P.; Yaya, S.; Baldiagai, J.; Madi, T.K.; Yougouda, A.; Andze, G.O.; Richards, F.O. Seventeen years of annual distribution of ivermectin has not interrupted onchocerciasis transmission in north region, cameroon. Am. J. Trop. Med. Hyg. 2011, 85, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.; Jones, F.M.; Fofana, H.K.; Doucoure, A.; Landoure, A.; Kimani, G.; Mwatha, J.K.; Sacko, M.; Vennervald, B.J.; Dunne, D.W. Rapidly boosted plasma il-5 induced by treatment of human schistosomiasis haematobium is dependent on antigen dose, ige and eosinophils. PLoS Negl. Trop. Dis. 2013, 7, e2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef]
- Rihet, P.; Demeure, C.E.; Bourgois, A.; Prata, A.; Dessein, A.J. Evidence for an association between human resistance to Schistosoma mansoni and high anti-larval ige levels. Eur. J. Immunol. 1991, 21, 2679–2686. [Google Scholar] [CrossRef] [PubMed]
- Coulibaly, J.T.; N’Goran, E.K.; Utzinger, J.; Doenhoff, M.J.; Dawson, E.M. A new rapid diagnostic test for detection of anti-Schistosoma mansoni and anti-schistosoma haematobium antibodies. Parasit Vectors 2013, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nausch, N.; Dawson, E.M.; Midzi, N.; Mduluza, T.; Mutapi, F.; Doenhoff, M.J. Field evaluation of a new antibody-based diagnostic for schistosoma haematobium and s. Mansoni at the point-of-care in northeast zimbabwe. BMC Infect. Dis. 2014, 14, 165. [Google Scholar] [CrossRef] [Green Version]
- Grenfell, R.; Martins, W.; Silva-Moraes, V.; Araujo, N.; Oliveira, E.; Fonseca, C.; Coelho, P.M. The schistosomula tegument antigen as a potential candidate for the early serological diagnosis of schistosomiasis mansoni. Rev. Inst. Med. Trop. Sao Paulo 2013, 55, 75–78. [Google Scholar] [CrossRef]
- Silveira, A.M.; Bethony, J.; Gazzinelli, A.; Kloos, H.; Fraga, L.A.; Alvares, M.C.; Prata, A.; Guerra, H.L.; Loverde, P.T.; Correa-Oliveira, R.; et al. High levels of igg4 to Schistosoma mansoni egg antigens in individuals with periportal fibrosis. Am. J. Trop. Med. Hyg. 2002, 66, 542–549. [Google Scholar] [CrossRef] [Green Version]
- Bonnard, P.; Remoue, F.; Schacht, A.M.; Deuffic-Burban, S.; Dompnier, J.P.; Elguero, E.; Charrier, N.; Cassagnou, M.; Diop, M.; Ly, A.; et al. Specific isotype immune response in the diagnosis of human schistosomiasis pathology? Am. J. Trop. Med. Hyg. 2004, 71, 202–205. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Covariate | OR | Confidence Interval | p Value * | |
|---|---|---|---|---|
| infected versus uninfected | ||||
| not exposed | 0.277 | [0.125–0.612] | 0.002 | |
| age group | 0.001 | |||
| young | 5.511 | [2.095–14.499] | 0.001 | |
| adolescent | 3.812 | [1.682–8.638] | 0.001 | |
| SmCTF IgG4 | 6.571 | [1.245–34.692] | 0.027 | |
| egg+ versus uninfected | ||||
| not exposed | 0.137 | [0.055–0.345] | 0.000 | |
| age group | 0.002 | |||
| young | 5.613 | [1.995–15.794] | 0.001 | |
| adolescent | 3.136 | [1.362–7.220 | 0.007 | |
| SmPCR+ versus uninfected | ||||
| age group | 0.034 | |||
| young | 4.130 | [1.149–14.849] | 0.030 | |
| adolescent | 3.502 | [1.277–9.604] | 0.015 | |
| SmCTF IgG1 × 100 | 1.262 | [1.100–1.448] | 0.001 | |
| egg+ versus SmPCR+ | ||||
| not exposed | 0.204 | [0.081–0.514] | 0.001 | |
| SmCTF IgG1 × 100 | 0.793 | [0.702–0.896] | 0.000 | |
| SmCTF IgG2 × 100 | 1.152 | [1.037–1.280] | 0.008 | |
| SmCTF IgG3 | 0.001 | [0.000–0.061] | 0.002 |
| Covariate | OR | Confidence Interval | p Value * | |
|---|---|---|---|---|
| infected versus uninfected | ||||
| not exposed | 0.282 | [0.113–0.707] | 0.007 | |
| age group | 0.037 | |||
| young | 3.582 | [1.223–10.490] | 0.020 | |
| adolescent | 2.750 | [1.055–7.172] | 0.039 | |
| SEA IgG4 | 14.816 | [5.956–36.885] | 0.000 | |
| egg+ versus uninfected | ||||
| not exposed | 0.185 | [0.065–0.528] | 0.002 | |
| SWA IgG2 | 0.170 | [0.023–1.239] | 0.080 | |
| SmCTF IgG4 | 0.019 | [0.002–0.220] | 0.002 | |
| SEA IgG4 | 49.667 | [12.957–190.384] | 0.000 | |
| SmPCR+ versus uninfected | ||||
| age group | 0.011 | |||
| young | 10.727 | [2.009–57.263] | 0.005 | |
| adolescent | 5.228 | [1.454–18.794] | 0.011 | |
| SEA IgG4 | 28.818 | [6.700–123.964] | 0.000 | |
| SEA IgE | 0.102 | [0.011–0.960] | 0.046 | |
| SWA IgG3 × 100 | 1.418 | [1.176–1.710] | 0.000 | |
| egg+ versus SmPCR+ | ||||
| not exposed | 0.124 | [0.042–0.367] | 0.000 | |
| SEA IgG4 | 3.095 | [1.111–8.619] | 0.031 | |
| SEA IgE | 12.212 | [1.580–94.354] | 0.016 | |
| SmCTF IgG1 × 100 | 0.802 | [0.698–0.921] | 0.002 | |
| SmCTF IgG2 × 100 | 1.150 | [1.028–1.287] | 0.015 | |
| SmCTF IgG3 | 0.000 | [0.000–0.018] | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arndts, K.; Elfaki, T.E.M.; Doenhoff, M.J.; Katawa, G.; Goreish, I.A.; Atti El Mekki, M.E.Y.A.; Hoerauf, A.; Ritter, M.; Layland, L.E. Distinct Schistosoma mansoni-Specific Immunoglobulin Subclasses Are Induced by Different Schistosoma mansoni Stages—A Tool to Decipher Schistosoma mansoni Infection Stages. Pathogens 2022, 11, 19. https://doi.org/10.3390/pathogens11010019
Arndts K, Elfaki TEM, Doenhoff MJ, Katawa G, Goreish IA, Atti El Mekki MEYA, Hoerauf A, Ritter M, Layland LE. Distinct Schistosoma mansoni-Specific Immunoglobulin Subclasses Are Induced by Different Schistosoma mansoni Stages—A Tool to Decipher Schistosoma mansoni Infection Stages. Pathogens. 2022; 11(1):19. https://doi.org/10.3390/pathogens11010019
Chicago/Turabian StyleArndts, Kathrin, Tayseer E. M. Elfaki, Michael J. Doenhoff, Gnatoulma Katawa, Ibtisam A. Goreish, Misk El Yemen A. Atti El Mekki, Achim Hoerauf, Manuel Ritter, and Laura E. Layland. 2022. "Distinct Schistosoma mansoni-Specific Immunoglobulin Subclasses Are Induced by Different Schistosoma mansoni Stages—A Tool to Decipher Schistosoma mansoni Infection Stages" Pathogens 11, no. 1: 19. https://doi.org/10.3390/pathogens11010019
APA StyleArndts, K., Elfaki, T. E. M., Doenhoff, M. J., Katawa, G., Goreish, I. A., Atti El Mekki, M. E. Y. A., Hoerauf, A., Ritter, M., & Layland, L. E. (2022). Distinct Schistosoma mansoni-Specific Immunoglobulin Subclasses Are Induced by Different Schistosoma mansoni Stages—A Tool to Decipher Schistosoma mansoni Infection Stages. Pathogens, 11(1), 19. https://doi.org/10.3390/pathogens11010019