
Schistosoma mansoni Adult Worm Protective and Diagnostic Proteins in n-Butanol Extracts Revealed by Proteomic Analysis

 , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Sm-AWBE Proteins Identified from the In-Gel and In-Solution Digestions
2.2. Integrated Analysis of Sm-AWBE Proteins
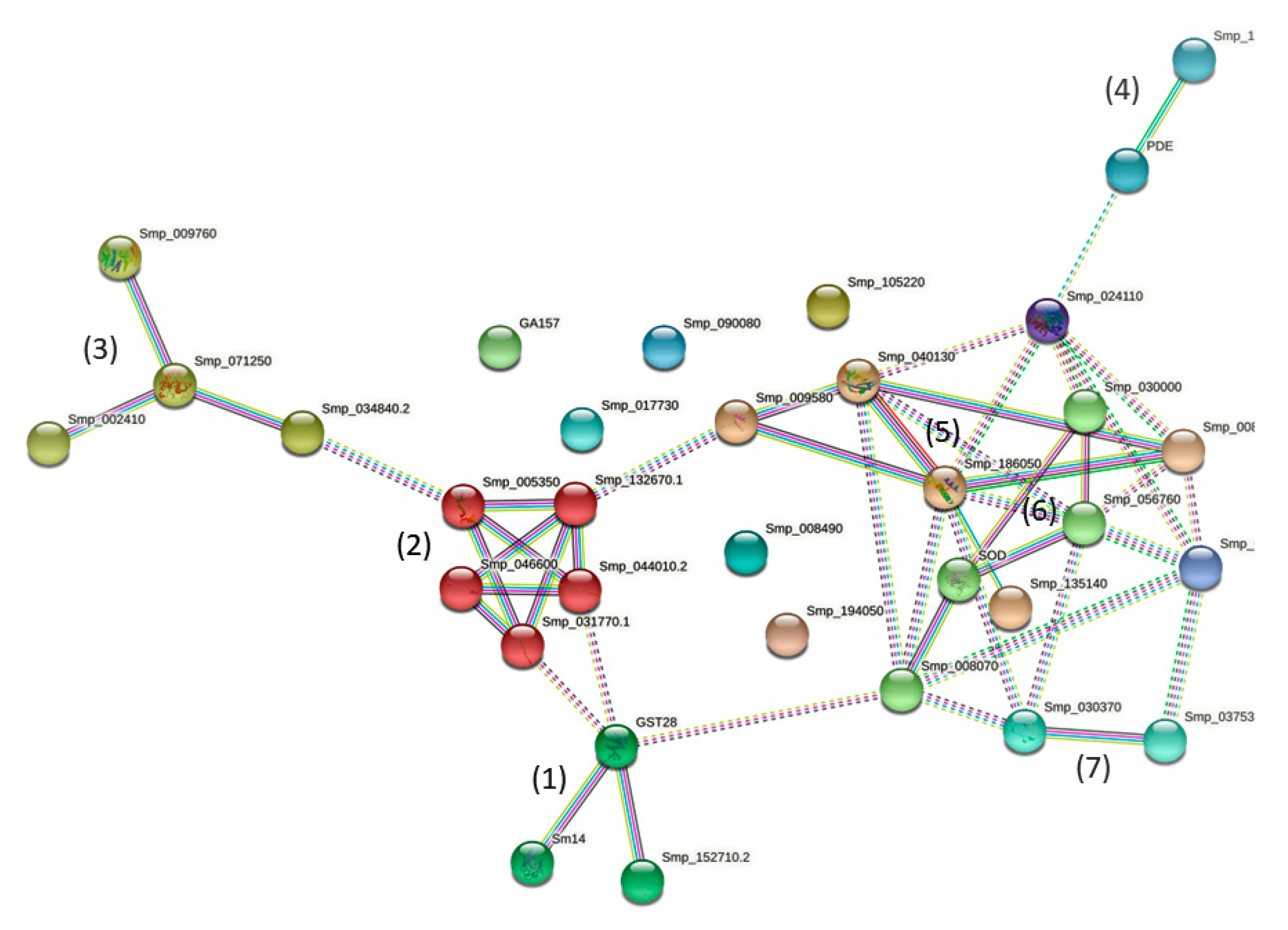
2.3. STRING Analyses of Sm-AWBE Proteins
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Obtention of Adult S. mansoni Worms
5.2. Adult S. mansoni Worms n-Butanol Extract (Sm-AWBE) Preparation
5.3. One-Dimensional Gel Electrophoresis Separation and In-Gel Protein Digestion of Sm-AWBE
5.4. In-Solution Digestion of Sm-AWBE Proteins
5.5. Mass Spectrometry
5.6. Data Analysis and Protein Identification
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sotillo, J.; Pearson, M.S.; Becker, L.; Mulvenna, J.; Loukas, A. A quantitative proteomic analysis of the tegumental proteins from Schistosoma mansoni schistosomula reveals novel potential therapeutic targets. Int. J. Parasitol. 2015, 45, 505–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, C.T.; Figueiredo Carvalho, G.B.; Alves, C.C.; Melo, T.T. Schistosoma tegument proteins in vaccine and diagnosis development: An update. J. Parasitol. Res. 2012, 5, 541268. [Google Scholar] [CrossRef]
- Pujol, F.H.; Alarcón de Noya, B.; Cesari, I.M. Immunodiagnosis of schistosomiasis mansoni with APIA (alkaline phosphatase immunoassay). Immunol. Investig. 1989, 18, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Cesari, I.M.; Ballen, D.E.; Mendoza, L.; Matos, C. Detection of Schistosoma mansoni membrane antigens by immunoblot analysis of sera of patients from low-transmission areas. Clin. Diagn. Lab. Immunol. 2005, 12, 280–286. [Google Scholar] [CrossRef] [Green Version]
- Cesari, I.M.; Ballén, D.E.; Mendoza, L.; Ferrer, A.; Pointier, J.P.; Kombila, M.; Richard-Lenoble, D.; Théron, A. Comparative evaluation of Schistosoma mansoni, Schistosoma intercalatum, and Schistosoma haematobium alkaline phosphatase antigenicity by the alkaline phosphatase immunoassay (APIA). Parasitol. Res. 2014, 113, 1395–1403. [Google Scholar] [CrossRef]
- Alarcón de Noya, B.; Cesari, I.M.; Losada, S.; Colmenares, S.; Balzán, C.; Hoebeke, J.; Noya, O. Evaluation of alkaline phosphatase immunoassay and comparison with other diagnostic methods in areas of low transmission of schistosomiasis. Acta Trop. 1997, 66, 69–78. [Google Scholar] [CrossRef]
- Cesari, I.M.; Bouty, I.; Bout, D.B.; de Noya, B.A.; Hoebeke, J. Parasite enzymes as a tool to investigate immune responses. Mem. Inst. Oswaldo Cruz 1992, 87, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Stepankova, V.; Bidmanova, S.; Koudelakova, T.; Prokop, Z.; Chaloupkova, R.; Damborsky, J. Strategies for stabilization of enzymes in organic solvents. ACS Catal. 2013, 12, 2823–2836. [Google Scholar] [CrossRef]
- Solá, R.J.; Griebenow, K. Effects of glycosylation on the stability of protein pharmaceuticals. J. Pharm. Sci. 2009, 98, 1223–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, E.J.; Sher, A.J. Three major surface antigens of Schistosoma mansoni are linked to the membrane by glycosylphosphatidylinositol. Immunology 1989, 142, 979–984. [Google Scholar]
- Pujol, F.H.; Cesari, I.M. Antigenicity of adult alkaline phosphatase. Parasite Immunol. 1990, 12, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, R.; Skelly, P.J. Characterization of Schistosome tegumental alkaline phosphatase (SmAP). PLoS Negl. Trop. Dis. 2011, 5, e1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffrey, C.R.; McKerrow, J.H.; Salter, J.P.; Sajid, M. Blood ‘n’ guts: An update on schistosome digestive peptidases. Trends Parasitol. 2004, 20, 241–248. [Google Scholar] [CrossRef]
- Cesari, I.M. Schistosoma mansoni: Distribution and characteristics of alkaline and acid phosphatase. Exp. Parasitol. 1974, 36, 405–414. [Google Scholar] [CrossRef]
- Cesari, I.M.; Simpson, A.J.H.; Evans, W.H. Properties of a series of tegumental membrane bound phosphohydrolase activities of Schistosoma mansoni. Biochem. J. 1981, 198, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Araujo-Montoya, B.O.; Rofatto, H.K.; Tararama, C.A.; Farias, L.P.; Oliveira, K.C.; Verjovski-Almeida, S.; Wilson, R.A.; Leite, L.C.C. Schistosoma mansoni: Molecular characterization of alkaline phosphatase and expression patterns across life cycle stages. Exp. Parasitol. 2011, 129, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Sulbarán, G.; Noya, O.; Brito, B.; Ballén, D.E.; Cesari, I.M. Immunoprotection of mice against Schistosomiasis mansoni using solubilized membrane antigens. PLoS Negl. Trop. Dis. 2013, 7, e2254. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.A.; Curwen, R.S.; Braschi, S.; Hall, S.L.; Coulson, P.S.; Ashton, P.D. From genomes to vaccines via the proteome. Mem. Inst. Oswaldo Cruz. 2004, 99, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Hansell, E.; Braschi, S.; Medzihradszky, K.F.; Sajid, M.; Debnath, M.; Ingram, J.; Lim, K.C.; McKerrow, J.H. Proteomic analysis of skin invasion by blood fluke larvae. PLoS Negl. Trop. Dis. 2008, 2, e262. [Google Scholar] [CrossRef] [Green Version]
- Losada, S.; Sabatier, L.; Hammann, P.; Lemaitre-Guillier, C.; Matos, C.; Bermúdez, H.; Lorenzo, M.; Noya, O. A combined proteomic and immunologic approach for the analysis of Schistosoma mansoni cercariae and adult worm protein extracts and the detection of one of the vaccine candidates, Sm28GST, from a Venezuelan parasite isolate. Investig. Clin. 2011, 52, 121–139. [Google Scholar]
- Mutapi, F. Helminth parasite proteomics: From experimental models to human infections. Parasitology 2012, 139, 1195–1204. [Google Scholar] [CrossRef] [Green Version]
- Nahum, L.A.; Mourão, M.M.; Oliveria, G. New frontiers in schistosoma genomics and transcriptomic. J. Parasitol. Res. 2012, 2012, 849132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, G.B.F.; Resende, D.M.; Siqueira, L.M.V.; Lopes, M.D.; Lopes, D.O.; Coelho, P.M.Z.; Texeira-Carvalho, A.; Ruiz, J.; Fonseca, C.T. Selecting targets for the diagnosis of Schistosoma mansoni infection: An integrative approach using multi-omic and immunoinformatics data. PLoS ONE 2017, 12, e0182299. [Google Scholar] [CrossRef] [Green Version]
- Samoil, V.; Dagenais, M.; Ganapathy, V.; Aldridge, J.; Glebov, A.; Jardim, A.; Ribeiro, P. Vesicle-based secretion in schistosomes: Analysis of protein and microRNA (miRNA) content of exosome-like vesicles derived from Schistosoma mansoni. Sci. Rep. 2018, 8, 3286. [Google Scholar] [CrossRef]
- Thiangtrongjit, T.; Adisakwattanab, P.; Limpanontc, Y.; Dekumyoyb, P.; Nuamtanongb, S.; Chusongsang, P.; Chusongsang, Y.; Reamtong, O. Proteomic and immunomic analysis of Schistosoma mekongi egg proteins. Exp. Parasitol. 2018, 191, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.P.; Robinson, M.W.; Hsieh, M.H.; Cody, J.; Le, L.; You, H.; McManus, D.P.; Gobert, G.N. A comparative proteomics analysis of the egg secretions of three major schistosome species. Mol. Biochem. Parasitol. 2020, 19, 111322. [Google Scholar] [CrossRef]
- Afonso, A.J.; Pinto Fonseca, S.; Pinto-Almeida, A.; Mendes, T.; Ferreira, P.; Belo, S.; Anibal, F.; Allegretti, S.; Galinaro, C.A.; Carrilho, E.; et al. Comparative proteomics reveals characteristic proteins on praziquantel-resistance in Schistosoma mansoni. Dryad 2021. [Google Scholar] [CrossRef]
- Farnell, E.J.; Tyagi, N.; Ryan, S.; Chalmers, I.W.; Pinot de Moira, A.; Jones, F.M.; Wawrzyniak, J.; Fitzsimmons, C.M.; Tukahebwa, E.M.; Furnham, N.; et al. Known allergen structures predict Schistosoma mansoni IgE-binding antigens in human infection. Front. Immunol. 2015, 6, 26. [Google Scholar] [CrossRef]
- Floudas, A.; Cluxton, C.D.; Fahel, J.; Khan, A.R.; Saunders, S.P.; Amu, S.; Alcami, A.; Fallon, P.G. Composition of the Schistosoma mansoni worm secretome: Identification of immune modulatory Cyclophilin, A. PLoS Negl. Trop. Dis. 2017, 11, e0006012. [Google Scholar] [CrossRef]
- Bout, D.; Deslée, D.; Capron, A. Antischistosomal effect of cyclosporin A: Cure and prevention of mouse and rat schistosomiasis mansoni. Infect. Immun. 1986, 52, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Balloul, J.M.; Boulanger, D.; Sondermeyer, P.; Dreyer, D.; Capron, M.; Grzych, J.M.; Pierce, R.; Carvallo, D.; Lecocq, J.P.; Capron, A. Vaccination of baboons with a P28 antigen of Schistosoma mansoni expressed in Escherichia coli. In Molecular Paradigms for Eradicating Helminthic Parasites; A. R. Liss: New York, NY, USA, 1987. [Google Scholar]
- Wolowczuk, I.; Auriault, C.; Bossus, M.; Boulanger, D.; Gras-Masse, H.; Mazingue, C.; Pierce, R.; Grezel, D.; Reid, G.D.; Tartar, A. Antigenicity and immunogenicity of a multiple peptidic construction of the Schistosoma mansoni Sm-28 GST antigen in rat, mouse, and monkey. 1. Partial protection of Fischer rat after active immunization. J. Immunol. 1991, 146, 1987–1995. [Google Scholar]
- Riveau, G.; Poulain-Godefroy, O.; Dupré, L.; Remoué, F.; Mielcarek, N.; Locht, C.; Capron, A. Glutathione S-Transferases of 28kDa as major vaccine candidates against Schistosomiasis. Mem. Inst. Oswaldo Cruz. 1998, 93, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girardini, J.; Amirante, A.; Zemzoumi, K.; Serra, E. Characterization of an omega-class glutathione S-transferase from Schistosoma mansoni with glutaredoxin-like dehydroascorbate reductase and thiol transferase activities. Eur. J. Biochem. 2002, 269, 5512–5521. [Google Scholar] [CrossRef]
- Torres-Rivera, A.; Landa, A. Glutathione transferases from parasites: A biochemical view. Acta Trop. 2008, 105, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Driss, V.; El Nady, M.; Delbeke, M.; Rousseaux, C.; Dubuquoy, C.; Sarazin, A.; Gatault, S.; Dendooven, A.; Riveau, G.; Colombel, J.F.; et al. The schistosome glutathione S-transferase P28GST, a unique helminth protein, prevents intestinal inflammation in experimental colitis through a Th2-type response with mucosal eosinophils. Mucosal Immunol. 2016, 9, 322–335. [Google Scholar] [CrossRef]
- Moser, D.; Tendler, M.; Griffiths, G.; Klinkert, M.Q. A 14-kDa Schistosoma mansoni polypeptide is homologous to a gene family of fatty acid binding proteins. J. Biol. Chem. 1991, 266, 8447–8454. [Google Scholar] [CrossRef]
- Tendler, M.; Almeida, M.S.; Vilar, M.M.; Pinto, P.M.; Limaverde-Sousa, G. Current status of the Sm14/GLA-SE schistosomiasis vaccine: Overcoming barriers and paradigms towards the first anti-parasitic human(itarian) vaccine. Trop. Med. Infect. Dis. 2018, 3, 121. [Google Scholar] [CrossRef] [Green Version]
- Eyayu, T.; Zeleke, A.J.; Worku, L. Current status and future prospects of protein vaccine candidates against Schistosoma mansoni infection. Parasite Epidemiol. Control. 2020, 11, e00176. [Google Scholar] [CrossRef]
- Silas, S.; Fitzsimmons, C.M.; Jones, F.M.; Pinto de Moira, A.; Wawrzyuniak, J.; Tukahebwa, E.M.; Dunne, D.W. Human IgE responses to different splice variants of Schistosoma mansoni tropomyosin: Associations with immunity. Int. J. Parasitol. 2014, 44, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Ludolf, F.; Patrocínio, P.R.; Corrêa-Oliveira, R.; Gazzinelli, A.; Falcone, F.H.; Teixeira-Ferreira, A.; Perales, J.; Oliveira, G.C.; Silva-Pereira, R.A. Serological screening of the Schistosoma mansoni adult worm proteome. PLoS Negl. Trop. Dis. 2014, 8, e2745. [Google Scholar] [CrossRef] [Green Version]
- Gnanasekar, M.; Salunkhe, A.M.; Mallia, A.K.; He, Y.X.; Kalyanasundaram, R. Praziquantel affects the regulatory myosin light chain of Schistosoma mansoni. Antimicrob. Agents Chemother. 2009, 53, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Havercroft, J.C.; Huggins, M.C.; Dunne, D.W.; Taylor, D.W. Characterization of Sm20, a 20-kilodalton calcium-binding protein of Schistosoma mansoni. Mol. Biochem. Parasitol. 1990, 38, 211–219. [Google Scholar] [CrossRef]
- Mohamed, M.M.; Shalaby, K.A.; LoVerde, P.T.; Karim, A.M. Characterization of Sm20.8, a member of a family of schistosome tegumental antigens. Mol. Biochem. Parasitol. 1998, 96, 15–25. [Google Scholar] [CrossRef]
- Cohen, C.; Reinhardt, B.; Castellani, L.; Norton, P.K.; Stirewalt, M. Schistosome surface spines are “crystals” of actin. J. Cell Biol. 1982, 195, 987–988. [Google Scholar] [CrossRef] [Green Version]
- Sulbarán, G.; Alamo, L.; Pinto, A.; Marquez, G.; Méndez, F.; Padrón, R.; Craig, R. An invertebrate smooth muscle with striated muscle myosin filaments. Proc. Natl. Acad. Sci. USA 2015, 112, E5660–E5668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.R.; Thulin, E.; Fagan, P.A.; Forsén, S.; Chazin, W.J. The EF-hand domain: A globally cooperative structural unit. Protein Sci. 2002, 11, 198–205. [Google Scholar] [CrossRef]
- Zhang, Y.; Taylor, M.G.; Johansen, M.V.; Bickle, Q.D. Vaccination of mice with a cocktail DNA vaccine induces a Th1-type immune response and partial protection against Schistosoma japonicum infection. Vaccine 2001, 20, 724–730. [Google Scholar] [CrossRef]
- Del Mar Siles-Lucas, M.; Gottstein, B. The 14-3-3 protein: A key molecule in parasites as in other organisms. Trends Parasitol. 2003, 19, 575–581. [Google Scholar] [CrossRef]
- Foote, M.; Zhou, Y. 14-3-3 proteins in neurological disorders. Int. J. Biochem. Mol. Biol. 2012, 3, 152–164. [Google Scholar]
- Carmona, G.; Göttig, S.; Orlandi, A.; Scheele, J.; Baüerle, T.; Jugold, M.; Kiessling, F.; Henschler, R.; Zeiher, A.M.; Dimmeler, S.; et al. Role of the small GTPase Rap1 for integrin activity regulation in endothelial cells and angiogenesis. Blood 2009, 113, 488–497. [Google Scholar] [CrossRef]
- Schechtman, D.; Tarrab-Hazdai, R.; Arnon, R. The 14-3-3 protein as a vaccine candidate against schistosomiasis. Parasite Immunol. 2001, 23, 213–217. [Google Scholar] [CrossRef]
- Elzoheiry, M.; Da’dara, A.A.; Bhardwaj, R.; Wang, Q.; Azab, M.S.; El-Kholy, E.-S.I.; El-Beshbishi, S.N.; Skelly, P.J. Intravascular Schistosoma mansoni cleave the host immune and hemostatic signaling molecule sphingosine-1-phosphate via tegumental alkaline phosphatase. Front. Immunol. 2018, 9, 1746. [Google Scholar] [CrossRef]
- Cesari, I.M.; Pujol, F.H.; Rodríguez, M.; Alarcón de Noya, B. Possible use of Schistosoma mansoni enzymes as antigens for immunodiagnosis. Mem. Inst. Oswaldo Cruz. 1987, 82, 175–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rofatto, H.K.; Araujo-Montoya, B.O.; Miyasato, P.A.; Levano-Garcia, J.; Rodriguez, D.; Nakano, E.; Verjovski-Almeida, S.; Farias, L.P.; Leite, L.C.C. Immunization with tegument nucleotidases associated with a subcurative praziquantel treatment reduces worm burden following Schistosoma mansoni challenge. PeerJ 2013, 1, e58. [Google Scholar] [CrossRef]
- Asseman, C.; Pancré, V.; Delanoye, A.; Capron, A.; Auriault, C.A. Radioimmunoassay for the quantification of human ubiquitin in biological fluids: Application to parasitic and allergic diseases. J. Immunol. Methods 1994, 173, 93–101. [Google Scholar] [CrossRef]
- Majetschak, M. Extracellular ubiquitin: Immune modulator and endogenous opponent of damage-associated molecular pattern molecules. J. Leukoc. Biol. 2011, 89, 205–219. [Google Scholar] [CrossRef]
- Kanamura, H.Y.; Hancock, K.; Rodrigues, V.; Damian, R.T. Schistosoma mansoni heat shock protein 70 p elicits an early humoral immune response in S. mansoni infected Baboons. Mem. Inst. Oswaldo Cruz. 2002, 97, 711–716. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Jin, X.; Chen, X.; Zhu, J.; Xu, Z.; Wang, X.; Liu, F.; Hu, W.; Zhou, L.; Su, C. Heat shock protein 60 in eggs specifically induces Tregs and reduces liver immunopathology in mice with Schistosomiasis japonica. PLoS ONE 2015, 10, e0139133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, W.; Li, Y.; Xu, L.; Zhou, S.; Zhu, J.; Xu, Z.; Liu, F.; Lin, D.; Hu, F.; et al. Elevated serum antibody against Schistosoma japonicum HSP60 as a promising biomarker for liver pathology in schistosomiasis. Sci. Rep. 2017, 7, 7765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador-Recataà, V.; Greenberg, R.M. Calcium channels of schistosomes: Unresolved questions and unexpected answers. WIREs Membr. Transp. Signal. 2012, 1, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, K.A.; Yin, L.; Thakur, A.; Christen, L.; Niles, E.G.; LoVerde, P.T. Protection against Schistosoma mansoni utilizing DNA vaccination with genes encoding Cu/Zn cytosolic superoxide dismutase, signal peptide-containing superoxide dismutase and glutathione peroxidase enzymes. Vaccine 2003, 22, 130–136. [Google Scholar] [CrossRef]
- Nordberg, J.; Arner, E.S.J. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med. 2001, 31, 1287–1312. [Google Scholar] [CrossRef]
- Kuntz, A.N.; Davioud-Charvet, E.; Sayed, A.A.; Califf, L.L.; Dessolin, J.; Arnér, E.S.J.; Williams, D.L. Thioredoxin glutathione reductase from Schistosoma mansoni: An essential parasite enzyme and a key drug target. PLoS Med. 2007, 4, e206. [Google Scholar] [CrossRef]
- Cao, X.; Hong, Y.; Zhang, M.; Han, Y.; Wu, M.; Wang, X.; Guo, X.; Li, C.; Lu, K.; Li, H.; et al. Cloning, expression and characterization of protein disulfide isomerase of Schistosoma japonicum. Exp. Parasitol. 2014, 146, 43–51. [Google Scholar] [CrossRef]
- Damonneville, M.; Auriault, C.; Pierce, R.J.; Capron, A. Antigenic properties of Schistosoma mansoni aminopeptidases: Evolution during the development in mammalian hosts. Mol. Biochem. Parasitol. 1982, 6, 265–275. [Google Scholar] [CrossRef]
- McCarthy, E.; Stack, C.; Donnelly, S.M.; Doyle, S.; Mann, V.H.; Brindley, P.J.; Stewart, M.; Day, T.A.; Maule, A.G.; Dalton, J.P. Leucine aminopeptidase of the human blood flukes, Schistosoma mansoni and Schistosoma japonicum. Int. J. Parasitol. 2004, 34, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Delgado, M.; Cortéz, J.; Sulbarán, G.; Matos, C.; Incani, R.N.; Ballén, D.E.; Cesari, I.M. Differential distribution and biochemical characteristics of hydrolases among developmental stages of Schistosoma mansoni may offer new anti-parasite targets. Parasitol. Int. 2017, 66, 816–820. [Google Scholar] [CrossRef]
- Ma, L.; Li, D.; Yuan, C.; Zhang, X.; Ta, N.; Zhao, X.; Li, Y.; Feng, X. SjCRT, a recombinant Schistosoma japonicum calreticulin, induces maturation of dendritic cells and a Th1-polarized immune response in mice. Parasit. Vectors 2017, 10, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Hosoi, F.; Yamaguchi-Iwai, Y.; Nakamura, H.; Masutani, H.; Ueda, S.; Nishiyama, A.; Takeda, S.; Wada, H.; Spyrou, G.; et al. Thioredoxin-2 (TRX-2) is an essential gene regulating mitochondria-dependent apoptosis. EMBO J. 2002, 21, 1695–1703. [Google Scholar] [CrossRef] [Green Version]
- Abath, F.G.; Xavier, E.M.; Allen, R.; Gomes, Y.M.; Lucena-Silva, N.; Baliza, M.; Simpson, A.J. Characterization of Sm13, a tegumental antigen of Schistosoma mansoni. Parasitol. Res. 2000, 86, 745–752. [Google Scholar] [CrossRef]
- Hall, T.M.; Joseph, G.T.; Strand, M. Schistosoma mansoni: Molecular cloning and sequencing of the 200-kDa chemotherapeutic target antigen. Exp. Parasitol. 1995, 80, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Braschi, S.; Castro-Borges, W.; Wilson, R.A. Proteomic analysis of the schistosome tegument and its surface membranes. Mem. Inst. Oswaldo Cruz 2006, 101, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Borges, W.; Dowle, A.; Curwen, R.S.; Thomas-Oates, J.; Wilson, R.A. Enzymatic shaving of the tegument surface of live schistosomes for proteomic analysis: A rational approach to select vaccine candidates. PLoS Negl. Trop. Dis. 2011, 5, e993. [Google Scholar] [CrossRef]
- Farias, L.P.; Tararam, C.A.; Miyasato, P.A.; Nishiyama Jr, M.Y.; Oliveira, K.C.; Kawano, T.; Verjovski-Almeida, S.; Leite, L.C.C. Screening the Schistosoma mansoni transcriptome for genes differentially expressed in the schistosomulum stage in search for vaccine candidates. Parasitol. Res. 2011, 108, 123–135. [Google Scholar] [CrossRef]
- Chalmers, I.W.; Fitzsimmons, C.M.; Brown, M.; Pierrot, C.; Jones, F.M.; Wawrzyniak, J.M.; Fernandez-Fuente, N.; Tukahebwa, E.M.; Dunne, D.W.; Khalife, J.; et al. Human IgG1 responses to surface localized Schistosoma mansoni Ly6 family members drop following praziquantel treatment. PLoS Negl. Trop. Dis. 2015, 9, e0003920. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Sun, X.; Chen, Y.; Zhan, B.; Zhu, X. Complement evasion: An effective strategy that parasites utilize to survive in the host. Front. Microbiol. 2020, 10, 532. [Google Scholar] [CrossRef] [PubMed]
- Tanigawa, C.; Fujii, Y.; Miura, M.; Nzou, S.M.; Mwangi, A.W.; Nagi, S.; Hamano, S.; Njenga, S.M.; Mbanefo, E.C.; Hirayama, K.; et al. Species-specific serological detection for schistosomiasis by serine protease inhibitor (SERPIN) in multiplex assay. PLoS Negl. Trop. Dis. 2015, 9, e0004021. [Google Scholar] [CrossRef] [Green Version]
- Leontovyč, A.; Ulrychová, L.; O’Donoghue, A.J.; Vondrášek, J.; Marešová, L.; Hubálek, M.; Fajtová, P.; Chanová, M.; Jiang, Z.; Craik, C.S.; et al. SmSP2: A serine protease secreted by the blood fluke pathogen Schistosoma mansoni with anti-hemostatic properties. PLoS Negl. Trop. Dis. 2018, 12, e0006446. [Google Scholar] [CrossRef] [Green Version]
- Simpson, A.J.G.; Rumjanek, F.D.; Payares, G.; Evans, W.H. Glycosyl transferase activities are associated with the surface membrane in adult Schistosoma mansoni. Mol. Biochem. Parasitol. 1981, 4, 107–115. [Google Scholar] [CrossRef]
- Cass, C.L.; Johnson, J.R.; Califf, L.L.; Xu, T.; Hernandez, H.J.; Stadecker, M.J.; Yates, J.R.; Williams, D.L. Proteomic analysis of Schistosoma mansoni egg secretions. Mol. Biochem. Parasitol. 2007, 155, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mebius, M.M.; van Genderen, P.J.J.; Urbanus, R.T.; Tielens, A.G.M.; de Groot, P.G.; van Hellemond, J.J. Interference with the host haemostatic system by schistosomes. PLoS Pathog. 2013, 9, e1003781. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Yu, C.X. Enolase and parasitic infection. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 2014, 26, 445–448. [Google Scholar]
- Figueiredo, B.C.; Da’dara, A.A.; Oliveira, S.C.; Skelly, P.J. Schistosomes enhance plasminogen activation: The role of tegumental enolase. PLoS Pathog. 2015, 11, e1005335. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Xiao, D.; Song, L.; Zhang, W.; Shen, S.; Yin, X.; Wang, J.; Ke, X.; Yu, C.; Zhang, J. Assessment of the diagnostic efficacy of enolase as an indication of active infection of Schistosoma japonicum. Parasitol. Res. 2015, 115, 151–164. [Google Scholar] [CrossRef]
- Onile, O.S.; Calder, B.; Soares, N.C.; Anumudu, C.I.; Blackburn, J.M. Quantitative label-free proteomic analysis of human urine to identify novel candidate protein biomarkers for schistosomiasis. PLoS Negl. Trop. Dis. 2017, 11, e0006045. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.H.; Binder, S.; King, C.H.; Knopp, S.; Rollinson, D.; Person, B.; Webster, B.; Allan, F.; Utzinger, J.; Ame, S.M.; et al. SCORE operational research on moving toward interruption of schistosomiasis transmission. Am. J. Trop. Med. Hyg. 2020, 103, 58–65. [Google Scholar] [CrossRef]
- Tebeje, B.M.; Harvie, M.; You, H.; Loukas, A.; McManus, D.P. Schistosomiasis vaccines: Where do we stand? Parasit. Vectors 2016, 30, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crosnier, C.; Brandt, C.; Rinaldi, G.; McCarthy, C.; Barker, C.; Clare, S.; Berriman, M.; Wright, G.J. Systematic screening of 96 Schistosoma mansoni cell-surface and secreted antigens does not identify any strongly protective vaccine candidates in a mouse model of infection. Wellcome Open Res. 2019, 4, 159. [Google Scholar] [CrossRef] [Green Version]
- Dean, D.A.; Murrell, K.D.; Xu, S.T.; Mangold, B.L. Immunization of mice with ultraviolet-irradiated Schistosoma mansoni cercariae: A re-evaluation. Am. J. Trop. Med. Hyg. 1983, 32, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Fukushige, M.; Mitchell, K.M.; Bourke, C.D.; Woolhouse, M.E.; Mutapi, F. A meta-analysis of experimental studies of attenuated Schistosoma mansoni vaccines in the mouse model. Front. Immunol. 2015, 27, 85. [Google Scholar] [CrossRef] [Green Version]
- Bergquist, N.R.; Colley, D.G. Schistosomiasis vaccines research to development. Parasitology 1998, 14, 99–104. [Google Scholar]
- Wilson, R.A. Proteomics at the schistosome-mammalian host interface: Any prospects for diagnostics or vaccines? Parasitology 2012, 139, 1178–1194. [Google Scholar] [CrossRef]
- Curwen, R.S.; Ashton, P.D.; Johnston, D.A.; Wilson, R.A. The Schistosoma mansoni soluble proteome: A comparison across four life-cycle stages. Mol. Biochem. Parasitol. 2004, 138, 57–66. [Google Scholar] [CrossRef]
- Barratt, M.D. Quantitative structure-activity relationships for skin permeability. Toxicol. Vitr. 1995, 9, 27–37. [Google Scholar] [CrossRef]
- Rutherford, B.J.; Dahl, R.H.; Price, R.E.; Szmidt, H.L.; Benke, P.I.; Mukhopadhyay, A.; Keasling, J.D. Functional genomic study of exogenous n-butanol stress in Escherichia coli. Appl. Environ. Microbiol. 2010, 76, 1935–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, E.; Pililzota, T.; Davies, P.R.; McVey, A.; French, C.E. Characterization of the effects of n-butanol on the cell envelope of E. coli. Appl. Micriobiol. Biotechnol. 2016, 100, 9653–9659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, R.M.; Carvalho-Queiroz, C.; Wilding, G.; LoVerde, P.T. Nucleic acid vaccination with Schistosoma mansoni antioxidant enzyme cytosolic superoxide dismutase and the structural protein filamin confers protection against the adult worm stage. Infect. Immun. 2004, 72, 6112–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, A.A.; Cook, S.K.; Williams, D.L. Redox balance mechanisms in Schistosoma mansoni rely on peroxiredoxins and albumin and implicate peroxiredoxins as novel drug targets. J. Biol. Chem. 2006, 281, 17001–17010. [Google Scholar] [CrossRef] [Green Version]
- Mourão, M.D.M.; Dinguirard, N.; Franco, G.R.; Yoshino, T.P. Role of the endogenous antioxidant system in the protection of Schistosoma mansoni primary sporocysts against exogenous oxidative stress. PLoS Negl. Trop. Dis. 2009, 3, e550. [Google Scholar] [CrossRef] [Green Version]
- Hinks, J.; Wang, Y.; Matysik, A.; Kraut, R.; Kjelleberg, S.; Mu, Y.; Bazan, G.; Wuertz, S.; Seviour, T. Increased microbial butanol tolerance by exogenous membrane insertion molecules. ChemSusChem 2015, 8, 3718–3726. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, S.A.; Gaida, S.M.; Papoutsakis, E.T. A comparative view of metabolite and substrate stress and tolerance in microbial bioprocessing: From biofuels and chemicals, to biocatalysis and bioremediation. Metab. Eng. 2010, 12, 307–331. [Google Scholar] [CrossRef]
- Aragon, A.D.; Imani, R.A.; Blackburn, V.R.; Cupit, P.M.; Melman, S.D.; Goronga, T.; Webb, T.; Loker, E.S.; Cunningham, C. Towards an understanding of the mechanism of action of praziquantel. Mol. Biochem. Parasitol. 2009, 164, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunne, D.W.; Webster, M.; Smith, P.; Langley, J.G.; Richardson, B.A.; Fulford, J.; Butterworth, A.E.; Sturrock, R.F.; Kariuki, H.C.; Ouma, J.H. The isolation of a 22 kDa band after SDS-PAGE of Schistosoma mansoni adult worms and its use to demonstrate that IgE responses against the antigen(s) it contains are associated with human resistance to reinfection. Parasite Immunol. 1997, 19, 79–89. [Google Scholar] [CrossRef]
- Labateya, N.; Thomson, D.M.; Durko, M.; Shenouda, G.; Robb, L.; Scanzano, R. Extraction of human organ-specific cancer neoantigens from cancer cells and plasma membranes with 1-butanol. Cancer Res. 1987, 47, 1058–1064. [Google Scholar]
- Liao, S.K.; Smith, J.W.; Kwong, P.C. Selective extraction by 1-butanol of surface glycoprotein antigens from human melanoma cells. Cancer Immunol. Immunother. 1984, 17, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Akaberi, M.; Emami, S.A.; Vatani, M.; Tayarani-Najaran, Z. Evaluation of antioxidant and anti-melanogenic activity of different extracts of aerial parts of N. sintenisii in murine melanoma B16F10 Cells. Iran. J. Pharm. Res. 2018, 17, 225–235. [Google Scholar]
- Noya, O.; Fermín, Z.; Alarcón de Noya, B.; Losada, S.; Colmenares, C.; Hermoso, T. Humoral immune response of children with chronic schistosomiasis. Isotype recognition of adult worm antigens. Parasite Immunol. 1995, 17, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Bradford, A. A rapid and sensitive method for the quantitation of micrograms quantities of protein utilizing the principle of protein-dye binding. Annal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Accession | Description (Short Names) |
|---|---|
| Smp_002410 | 14-3-3 epsilon 2 |
| Smp_005350 | 20 kDa calcium-binding protein (SM20) |
| Smp_008070 | Thioredoxin (Trx) |
| Smp_008490 | Glycogenin-related |
| Smp_008545 | Heat shock protein 60 (HSP60) |
| Smp_009580 | Ubiquitin (UB) |
| Smp_009760 | 14-3-3 protein homolog 1 |
| Smp_017730 | 200 kDa GPI-anchored surface protein (Sm200) |
| Smp_024110 | Enolase/Phosphopyruvate hydratase (ENO) |
| Smp_030000 | Leucine aminopeptidase (M17)/putative cytosol aminopeptidase (LAP) |
| Smp_030370 | Calreticulin (CALR) |
| Smp_031770 | Tropomyosin-2 (TPM-2) |
| Smp_034840 | 14-3-3 protein homolog 2 (14-3-3 protein epsilon) |
| Smp_037530 | Mitochondrial thioredoxin (Mt Trx) |
| Smp_040130 | Peptidyl-prolyl cis-trans isomerase (CyP A) (PPIA) |
| Smp_044010 | Tropomyosin-1 (TPM-1) |
| Smp_046600 | Actin (s) |
| Smp_046740 | Dihydrolipoyl dehydrogenase (DLD) |
| Smp_054160 | Glutathione S-transferase class-mu 28 kDa isozyme (GST28) |
| Smp_056760 | Protein disulfide-isomerase (PDI) |
| Smp_071250 | Putative rap1 and (Rap1) |
| Smp_090080 | Serpin, putative |
| Smp_095360 | 14 kDa fatty acid-binding protein (FABP/Sm14) |
| Smp_105220 | DIF_5 (CD59-like, SmLy6B) |
| Smp_106930 | Heat shock 70 kDa protein homolog (HSP70) |
| Smp_132670 | Myosin regulatory light chain, putative (MRLC) |
| Smp_135140 | High voltage-activated calcium channel Cav1 |
| Smp_152710 | Glutathione-S-transferase omega, putative (GST omega) |
| Smp_153390 | Nucleotide pyrophosphatase/phosphodiesterase 5 (NPP-5)/Ecto-phosphodiesterase (PDE) |
| Smp_155890 | Alkaline phosphatase (AP) |
| Smp_176200 | Superoxide dismutase (Cu-Zn) (SOD Cu-Zn) |
| Smp_194050 | Clumping factor A (Fibrinogen-binding protein A) (Fibrinogen receptor A), putative (clfA) |
| Smp_195190 | 13 kDa tegumental antigen Sm13/GA157 (Sm13) |
| Extracellular | Signaling | Intracellular | Organelles | Transport/Motion | Regulatory | Cat. | Immunoactive | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession | Proteins Short Names | Av. Mass | Scr | TgS | Rcp | Trd | TrM | Cyk | Str | Cyt | AWE | ER | Mit | Ca-b | Trp | Msl | Reg | RxD | Ch/F | Imd | Inh | Cat. | All | Vac | Ags | PPIs |
| Smp_002410 | 14-3-3 epsilon 2 | 28432 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_005350 | SM20 | 57939 | ● | ● | ● | ● | + | |||||||||||||||||||
| Smp_008070 | Trx | 11924 | ● | ● | ● | ● | ● | + | ||||||||||||||||||
| Smp_008490 | Glycogenin-related | 36511 | ● | ● | ● | - | ||||||||||||||||||||
| Smp_008545 | HSP60 | 59554 | ● | ● | ● | ● | + | |||||||||||||||||||
| Smp_009580 | UB | 18327 | ● | ● | ● | ● | ● | + | ||||||||||||||||||
| Smp_009760 | 14-3-3 protein homolog 1 | 28371 | ● | ● | ● | ● | ● | + | ||||||||||||||||||
| Smp_017730 | Sm200 | 186533 | ● | ● | ● | ● | + | |||||||||||||||||||
| Smp_024110 | ENO | 47006 | ● | ● | ● | ● | ● | ● | ● | + | ||||||||||||||||
| Smp_030000 | LAP | 55676 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_030370 | CALR | 45377 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_031770 | TPM-2 | 32696 | ● | ● | ● | + | ||||||||||||||||||||
| Smp_034840 | 14-3-3 protein epsilon | 28754 | ● | ● | ● | ● | ● | + | ||||||||||||||||||
| Smp_037530 | Mt Trx | 12424 | ● | ● | ● | + | ||||||||||||||||||||
| Smp_040130 | PPIA (CyP A) | 17671 | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | + | ||||||||||||
| Smp_044010 | TPM-1 | 32954 | ● | ● | ● | ● | + | |||||||||||||||||||
| Smp_046600 | Actin (s) | 41731 | ● | ● | ● | ● | ● | ● | ● | ● | ● | + | ||||||||||||||
| Smp_046740 | DLD | 52868 | ● | ● | ● | ● | + | |||||||||||||||||||
| Smp_054160 | GST28 | 23819 | ● | ● | ● | ● | ● | ● | ● | ● | + | |||||||||||||||
| Smp_056760 | PDI | 54160 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_071250 | Rap1 | 20700 | ● | ● | ● | ● | + | |||||||||||||||||||
| Smp_090080 | Serpin, putative | 46005 | ● | ● | ● | ● | ● | ● | - | |||||||||||||||||
| Smp_095360 | FABP/Sm14 | 11923 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_105220 | DIF_5 | 14191 | ● | ● | ● | ● | ● | - | ||||||||||||||||||
| Smp_106930 | HSP70 | 69859 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_132670 | MRLC | 22719 | ● | ● | ● | + | ||||||||||||||||||||
| Smp_135140 | Cav1 | 87668 | ● | ● | ● | - | ||||||||||||||||||||
| Smp_152710 | GST omega | 27532 | ● | ● | ● | + | ||||||||||||||||||||
| Smp_153390 | NPP-5/PDE | 51434 | ● | ● | ● | ● | ● | ● | + | |||||||||||||||||
| Smp_155890 | AP | 46318 | ● | ● | ● | ● | ● | + | ||||||||||||||||||
| Smp_176200 | SOD Cu-Zn | 15910 | ● | ● | ● | ● | ● | ● | ● | ● | + | |||||||||||||||
| Smp_194050 | clfA | 43482 | ● | ● | - | |||||||||||||||||||||
| Smp_195190 | Sm13/GA157 | 11922 | ● | ● | ● | ● | - | |||||||||||||||||||
| n = 33 | Scr | TgS | Rcp | Trd | TrM | Cyk | Str | Cyt | AWE | ER | Mit | Ca-b | Trp | Msl | Reg | RxD | Ch/F | Imd | Inh | Cat. | All | Vac | Ags | |||
| 23 | 10 | 13 | 6 | 10 | 23 | 15 | 27 | |||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulbarán, G.; Verissimo da Costa, G.C.; Losada, S.; Peralta, J.M.; Cesari, I.M. Schistosoma mansoni Adult Worm Protective and Diagnostic Proteins in n-Butanol Extracts Revealed by Proteomic Analysis. Pathogens 2022, 11, 22. https://doi.org/10.3390/pathogens11010022
Sulbarán G, Verissimo da Costa GC, Losada S, Peralta JM, Cesari IM. Schistosoma mansoni Adult Worm Protective and Diagnostic Proteins in n-Butanol Extracts Revealed by Proteomic Analysis. Pathogens. 2022; 11(1):22. https://doi.org/10.3390/pathogens11010022
Chicago/Turabian StyleSulbarán, Guidenn, Giovani C. Verissimo da Costa, Sandra Losada, José M. Peralta, and Italo M. Cesari. 2022. "Schistosoma mansoni Adult Worm Protective and Diagnostic Proteins in n-Butanol Extracts Revealed by Proteomic Analysis" Pathogens 11, no. 1: 22. https://doi.org/10.3390/pathogens11010022
APA StyleSulbarán, G., Verissimo da Costa, G. C., Losada, S., Peralta, J. M., & Cesari, I. M. (2022). Schistosoma mansoni Adult Worm Protective and Diagnostic Proteins in n-Butanol Extracts Revealed by Proteomic Analysis. Pathogens, 11(1), 22. https://doi.org/10.3390/pathogens11010022