

Genome-Wide Searching Single Nucleotide-Polymorphisms (SNPs) and SNPs-Targeting a Multiplex Primer for Identification of Common Salmonella Serotypes

Abstract
:1. Introduction
2. Results
2.1. Acquired Salmonella WGS from GenBank
2.2. Searching SNP Sites from NGS of Salmonella Genome Alignment and Design Primers Based on SNP Sites
2.3. Amplified Target Salmonella Serovars with Newly Designed Encompassing-Primer Sets
2.4. Justification of SNP Sites on Target Salmonella Gene Sequences and Design with Serotype-Specific SNP Primers
2.5. Salmonella Serotype-Specific-SNP Primers Design Based on the Appropriate SNP Sites on the Aligned Gene Sequences
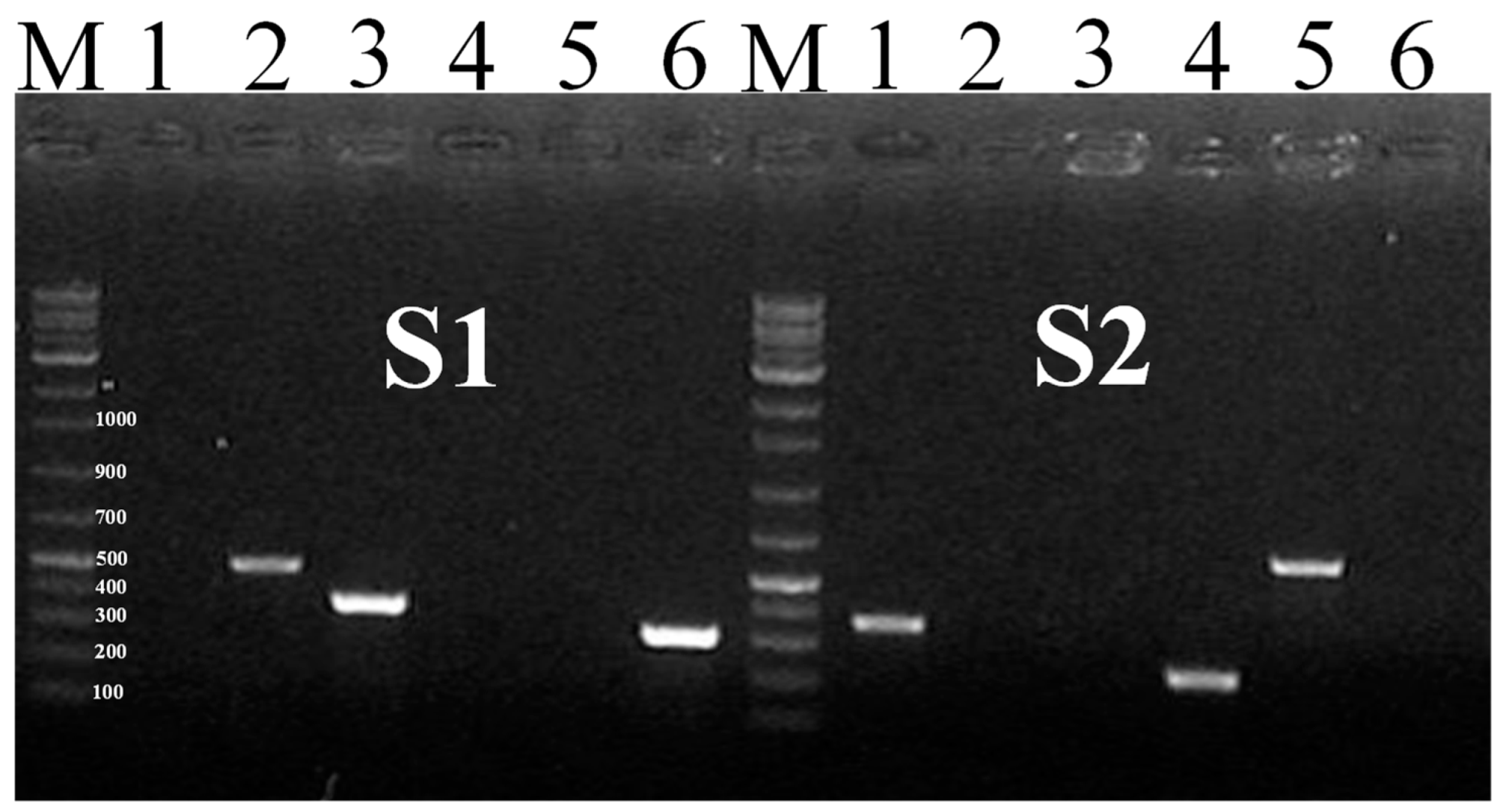
2.6. SNP-Based Multiplex PCR
2.7. Validation of SNP-Based Multiplex Marker with Isolated Salmonella Strains from Wild Animal
3. Discussion
4. Materials and Methods
4.1. Acquired Salmonella Whole-Genome Sequences (WGS) from GenBank, Searched SNPs, and Designed SNPs-Encompassing Primers
4.2. Selection and Isolation of Genomic DNA from Serotype-Specific Target Salmonella Serovar Strains
4.3. Salmonella Serotype-Specific SNP-Based Multiplex-PCR Marker
4.4. Validation of SNP-Based Multiplex Marker with Reference Strains and Laboratory Isolated Salmonella Strains
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- den Bakker, H.C.; Moreno Switt, A.I.; Cummings, C.A.; Hoelzer, K.; Degoricija, L.; Rodriguez-Rivera, L.D.; Wright, E.M.; Fang, R.; Davis, M.; Root, T. A Whole-Genome Single Nucleotide Polymorphism-Based Approach to Trace and Identify Outbreaks Linked to a Common Salmonella Enterica Subsp. Enterica Serovar Montevideo Pulsed-Field Gel Electrophoresis Type. Appl. Environ. Microbiol. 2011, 77, 8648–8655. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Lappi, V.; Wolfgang, W.J.; Lapierre, P.; Palumbo, M.J.; Medus, C.; Boxrud, D. Characterization of Foodborne Outbreaks of Salmonella Enterica Serovar Enteritidis with Whole-Genome Sequencing Single Nucleotide Polymorphism-Based Analysis for Surveillance and Outbreak Detection. J. Clin. Microbiol. 2015, 53, 3334–3340. [Google Scholar] [CrossRef] [PubMed]
- Pightling, A.W.; Pettengill, J.B.; Luo, Y.; Baugher, J.D.; Rand, H.; Strain, E. Interpreting Whole-Genome Sequence Analyses of Foodborne Bacteria for Regulatory Applications and Outbreak Investigations. Front. Microbiol. 2018, 9, 1482. [Google Scholar] [CrossRef] [PubMed]
- Dróżdż, M.; Małaszczuk, M.; Paluch, E.; Pawlak, A. Zoonotic Potential and Prevalence of Salmonella Serovars Isolated from Pets. Infect. Ecol. Epidemiol. 2021, 11, 1975530. [Google Scholar] [CrossRef]
- Dekker, J.P.; Frank, K.M. Salmonella, Shigella, and Yersinia. Clin. Lab. Med. 2015, 35, 225–246. [Google Scholar] [CrossRef]
- Ye, Q.; Shang, Y.; Chen, M.; Pang, R.; Li, F.; Xiang, X.; Wang, C.; Zhou, B.; Zhang, S.; Zhang, J. Identification of Novel Sensitive and Reliable Serovar-Specific Targets for PCR Detection of Salmonella Serovars Hadar and Albany by Pan-Genome Analysis. Front. Microbiol. 2021, 12, 540. [Google Scholar] [CrossRef]
- Branchu, P.; Bawn, M.; Kingsley, R.A. Genome Variation and Molecular Epidemiology of Salmonella Enterica Serovar Typhimurium Pathovariants. Infect. Immun. 2018, 86, e00079-18. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. Multi-Country Outbreak of Salmonella Agona Infections Possibly Linked to Ready-to-Eat Food; Wiley Online Library: Hoboken, NJ, USA, 2018. [Google Scholar]
- Jourdan-da Silva, N.; Fabre, L.; Robinson, E.; Fournet, N.; Nisavanh, A.; Bruyand, M.; Mailles, A.; Serre, E.; Ravel, M.; Guibert, V. Ongoing Nationwide Outbreak of Salmonella Agona Associated with Internationally Distributed Infant Milk Products, France, December 2017. Eurosurveillance 2018, 23, 17–852. [Google Scholar] [CrossRef]
- Marchello, C.S.; Birkhold, M.; Crump, J.A.; Martin, L.B.; Ansah, M.O.; Breghi, G.; Canals, R.; Fiorino, F.; Gordon, M.A.; Kim, J.-H. Complications and Mortality of Non-Typhoidal Salmonella Invasive Disease: A Global Systematic Review and Meta-Analysis. Lancet Infect. Dis. 2022, 22, 692–705. [Google Scholar] [CrossRef]
- Allgayer, M.C.; Lima-Rosa, C.A.V.; Weimer, T.A.; Rodenbusch, C.R.; Pereira, R.A.; Streck, A.F.; Oliveira, S.D.; Canal, C.W. Molecular Diagnosis of Salmonella Species in Captive Psittacine Birds. Vet. Rec. 2008, 162, 816–819. [Google Scholar] [CrossRef]
- de Souza, M.L.; Coelho, M.L.; da Silva, A.O.; da Silva Azuaga, L.B.; Macedo Coutinho Netto, C.R.; Galhardo, J.A.; Brito Leal, C.R.; do Nascimento Ramos, C.A. Salmonella Spp. Infection in Psittacidae at a Wildlife Rehabilitation Center in the State of Mato Grosso Do Sul, Brazil. J. Wildl. Dis. 2020, 56, 288–293. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, E.J.E.; Lopes, A.T.S.; Maciel, B.M. Salmonella in Wild Animals: A Public Health Concern. In Enterobacteria [Working Title]; IntechOpen: London, UK, 2022. [Google Scholar]
- Piras, F.; Spanu, V.; Siddi, G.; Gymoese, P.; Spanu, C.; Cibin, V.; Schjørring, S.; De Santis, E.P.L.; Scarano, C. Whole-Genome Sequencing Analysis of Highly Prevalent Salmonella Serovars in Wild Boars from a National Park in Sardinia. Food Control 2021, 130, 108247. [Google Scholar] [CrossRef]
- Siddiky, N.A.; Sarker, S.; Khan, S.R.; Rahman, T.; Kafi, A.; Samad, M.A. Virulence and Antimicrobial Resistance Profile of Non-Typhoidal Salmonella Enterica Serovars Recovered from Poultry Processing Environments at Wet Markets in Dhaka, Bangladesh. PLoS ONE 2022, 17, e0254465. [Google Scholar] [CrossRef] [PubMed]
- Gutema, F.D.; Agga, G.E.; Abdi, R.D.; De Zutter, L.; Duchateau, L.; Gabriël, S. Prevalence and Serotype Diversity of Salmonella in Apparently Healthy Cattle: Systematic Review and Meta-Analysis of Published Studies, 2000–2017. Front. Vet. Sci. 2019, 6, 102. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hur, S.J.; Yim, D.G. Monitoring of Microbial Contaminants of Beef, Pork, and Chicken in HACCP Implemented Meat Processing Plants of Korea. Korean J. food Sci. Anim. Resour. 2018, 38, 282. [Google Scholar]
- Kim, E.; Park, S.; Cho, S.; Hahn, T.-W.; Yoon, H. Comparative Genomics Approaches to Understanding Virulence and Antimicrobial Resistance of Salmonella Typhimurium ST1539 Isolated from a Poultry Slaughterhouse in Korea. J. Microbiol. Biotechnol. 2019, 29, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Liu, Y.; Qin, X.; Aspridou, Z.; Zheng, J.; Wang, X.; Li, Z.; Dong, Q. The Prevalence and Epidemiology of Salmonella in Retail Raw Poultry Meat in China: A Systematic Review and Meta-Analysis. Foods 2021, 10, 2757. [Google Scholar] [CrossRef]
- Tang, S.; Orsi, R.H.; Luo, H.; Ge, C.; Zhang, G.; Baker, R.C.; Stevenson, A.; Wiedmann, M. Assessment and Comparison of Molecular Subtyping and Characterization Methods for Salmonella. Front. Microbiol. 2019, 10, 1591. [Google Scholar] [CrossRef]
- Ricke, S.C.; Kim, S.A.; Shi, Z.; Park, S.H. Molecular-based Identification and Detection of Salmonella in Food Production Systems: Current Perspectives. J. Appl. Microbiol. 2018, 125, 313–327. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards; Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F. Whole Genome Sequencing and Metagenomics for Outbreak Investigation, Source Attribution and Risk Assessment of Food-borne Microorganisms. EFSA J. 2019, 17, e05898. [Google Scholar]
- Ibrahim, G.M.; Morin, P.M. Salmonella Serotyping Using Whole Genome Sequencing. Front Microbiol. 2018, 9, 2993. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-S.; Besser, T.E.; Hancock, D.D.; Porwollik, S.; McClelland, M.; Call, D.R. Identification of Specific Gene Sequences Conserved in Contemporary Epidemic Strains of Salmonella Enterica. Appl. Environ. Microbiol. 2006, 72, 6938–6947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schut, C.H.; Farzan, A.; Fraser, R.S.; Ainslie-Garcia, M.H.; Friendship, R.M.; Lillie, B.N. Identification of Single-Nucleotide Variants Associated with Susceptibility to Salmonella in Pigs Using a Genome-Wide Association Approach. BMC Vet. Res. 2020, 16, 138. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Davies, R.; Petrovska, L. Identification of Genetic Features for Attenuation of Two Salmonella Enteritidis Vaccine Strains and Differentiation of These from Wildtype Isolates Using Whole Genome Sequencing. Front. Vet. Sci. 2019, 447. [Google Scholar] [CrossRef]
- Zou, Q.-H.; Li, R.-Q.; Wang, Y.-J.; Liu, S.-L. Identification of Genes to Differentiate Closely Related Salmonella Lineages. PLoS ONE 2013, 8, e55988. [Google Scholar] [CrossRef]
- Xin, S.; Zhu, H.; Tao, C.; Zhang, B.; Yao, L.; Zhang, Y.; Afayibo, D.J.A.; Li, T.; Tian, M.; Qi, J. Rapid Detection and Differentiating of the Predominant Salmonella Serovars in Chicken Farm by TaqMan Multiplex Real-Time PCR Assay. Front. Cell. Infect. Microbiol. 2021, 925. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, H.J.; Cho, W.H.; Kim, J.H.; Oh, M.H.; Kim, S.H.; Lee, B.K.; Ricke, S.C.; Kim, H.Y. Identification of Salmonella Enterica Subspecies I, Salmonella Enterica Serovars Typhimurium, Enteritidis and Typhi Using Multiplex PCR. FEMS Microbiol. Lett. 2009, 301, 137–146. [Google Scholar] [CrossRef]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-Analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). National Enteric Disease Surveillance: Salmonella Annual Report, 2016; Centers for Disease Control and Prevention: Atlanta, GA, USA, 28 February 2018.
- Centers for Disease Control and Prevention (CDC). National Enteric Disease Surveillance: Salmonella Annual Summary, 2009; Centers for Disease Control and Prevention: Atlanta, GA, USA, February 2011.
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Lo Fo Wong, D.M.A.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global Monitoring of Salmonella Serovar Distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of Quality Assured Laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef]
- Popa, G.L.; Papa, M.I. Salmonella Spp. Infection-a Continuous Threat Worldwide. Germs 2021, 11, 88. [Google Scholar] [CrossRef]
- Zeinzinger, J.; Pietzka, A.T.; Stöger, A.; Kornschober, C.; Kunert, R.; Allerberger, F.; Mach, R.; Ruppitsch, W. One-Step Triplex High-Resolution Melting Analysis for Rapid Identification and Simultaneous Subtyping of Frequently Isolated Salmonella Serovars. Appl. Environ. Microbiol. 2012, 78, 3352–3360. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.; Baratpour, A.; Khanzade, S.; Jamshidi, A. Salmonella Enteritidis and Salmonella Typhimorium Identification in Poultry Carcasses. Iran. J. Microbiol. 2018, 10, 45–50. [Google Scholar] [PubMed]
- Skov, M.N.; Madsen, J.J.; Rahbek, C.; Lodal, J.; Jespersen, J.B.; Jørgensen, J.C.; Dietz, H.H.; Chriél, M.; Baggesen, D.L. Transmission of Salmonella between Wildlife and Meat-production Animals in Denmark. J. Appl. Microbiol. 2008, 105, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Azad, M.; Kalam, O.; Uddain, J.; Adnan, M.; Ali, M.; Al-Mujahidy, S.K.; Roni, M.; Kadir, Z.; Rahman, M.S. Microbial Quality Assessment and Efficacy of Low-Cost Disinfectants on Fresh Fruits and Vegetables Collected from Urban Areas of Dhaka, Bangladesh. Foods 2021, 10, 1325. [Google Scholar] [CrossRef] [PubMed]
- Ha, A.J.; Perez, L.G.S.; Kim, T.-J.; Mizan, M.F.R.; Nahar, S.; Park, S.-H.; Chun, H.-S.; Ha, S.-D. Research Note: Identification and Characterization of Salmonella Spp. in Mechanically Deboned Chickens Using Pulsed-Field Gel Electrophoresis. Poult. Sci. 2021, 100, 100961. [Google Scholar] [CrossRef] [PubMed]
- Rabsch, W. Salmonella Typhimurium Phage Typing for Pathogens. In Salmonella; Springer: Berlin, Germany, 2007; pp. 177–211. [Google Scholar]
- Alcaine, S.D.; Soyer, Y.; Warnick, L.D.; Su, W.-L.; Sukhnanand, S.; Richards, J.; Fortes, E.D.; McDonough, P.; Root, T.P.; Dumas, N.B. Multilocus Sequence Typing Supports the Hypothesis That Cow-and Human-Associated Salmonella Isolates Represent Distinct and Overlapping Populations. Appl. Environ. Microbiol. 2006, 72, 7575–7585. [Google Scholar] [CrossRef]
- Ben-Darif, E.; Jury, F.; De Pinna, E.; Threlfall, E.J.; Bolton, F.J.; Fox, A.J.; Upton, M. Development of a Multiplex Primer Extension Assay for Rapid Detection of Salmonella Isolates of Diverse Serotypes. J. Clin. Microbiol. 2010, 48, 1055–1060. [Google Scholar] [CrossRef]
- Rahman, M.-M.; Lim, S.; Park, Y.-C. Development of Single Nucleotide Polymorphism (SNP)-Based Triplex PCR Marker for Serotype-Specific Escherichia Coli Detection. Pathogens 2022, 11, 115. [Google Scholar] [CrossRef]
- Kim, T.-W.; Jang, Y.-H.; Jeong, M.K.; Seo, Y.; Park, C.H.; Kang, S.; Lee, Y.J.; Choi, J.-S.; Yoon, S.-S.; Kim, J.M. Single-Nucleotide Polymorphism-Based Epidemiological Analysis of Korean Mycobacterium Bovis Isolates. J. Vet. Sci. 2021, 22, e24. [Google Scholar] [CrossRef]
- Uelze, L.; Grützke, J.; Borowiak, M.; Hammerl, J.A.; Juraschek, K.; Deneke, C.; Tausch, S.H.; Malorny, B. Typing Methods Based on Whole Genome Sequencing Data. One Heal. Outlook 2020, 2, 1–19. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, H.; Chen, J.; Chen, L.; Qi, X.; Zhang, R. Epidemiology of Foodborne Disease Outbreaks Caused by Nontyphoidal Salmonella in Zhejiang Province, China, 2010–2019. Foodborne Pathog. Dis. 2021, 18, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Uelze, L.; Becker, N.; Borowiak, M.; Busch, U.; Dangel, A.; Deneke, C.; Fischer, J.; Flieger, A.; Hepner, S.; Huber, I. Toward an Integrated Genome-Based Surveillance of Salmonella Enterica in Germany. Front. Microbiol. 2021, 12, 200. [Google Scholar] [CrossRef] [PubMed]
- Behl, A.; Nair, A.; Mohagaonkar, S.; Yadav, P.; Gambhir, K.; Tyagi, N.; Sharma, R.K.; Butola, B.S.; Sharma, N. Threat, Challenges, and Preparedness for Future Pandemics: A Descriptive Review of Phylogenetic Analysis Based Predictions. Infect. Genet. Evol. 2022, 98, 105217. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A.; Issenhuth-Jeanjean, S.; Didelot, X.; Thorell, K.; Hale, J.; Parkhill, J.; Thomson, N.R.; Weill, F.-X.; Falush, D.; Brisse, S. The Speciation and Hybridization History of the Genus Salmonella. Microb. Genom. 2019, 5, e000284. [Google Scholar] [CrossRef]
- Guard, J.; Abdo, Z.; Byers, S.O.; Kriebel, P.; Rothrock Jr, M.J. Subtyping of Salmonella Enterica Subspecies I Using Single-Nucleotide Polymorphisms in Adenylate Cyclase. Foodborne Pathog. Dis. 2016, 13, 350–362. [Google Scholar] [CrossRef]
- Roumagnac, P.; Weill, F.-X.; Dolecek, C.; Baker, S.; Brisse, S.; Chinh, N.T.; Le, T.A.H.; Acosta, C.J.; Farrar, J.; Dougan, G. Evolutionary History of Salmonella Typhi. Science 2006, 314, 1301–1304. [Google Scholar] [CrossRef]
- Octavia, S.; Lan, R. Single Nucleotide Polymorphism Typing of Global Salmonella Enterica Serovar Typhi Isolates by Use of a Hairpin Primer Real-Time PCR Assay. J. Clin. Microbiol. 2010, 48, 3504–3509. [Google Scholar] [CrossRef]
- Ogunremi, D.; Kelly, H.; Dupras, A.A.; Belanger, S.; Devenish, J. Development of a New Molecular Subtyping Tool for Salmonella Enterica Serovar Enteritidis Based on Single Nucleotide Polymorphism Genotyping Using PCR. J. Clin. Microbiol. 2014, 52, 4275–4285. [Google Scholar] [CrossRef]
- Gaudet, M.; Fara, A.-G.; Beritognolo, I.; Sabatti, M. Allele-Specific PCR in SNP Genotyping. In Single Nucleotide Polymorphisms; Springer: Berlin, Germany, 2009; pp. 415–424. [Google Scholar]
- Shen, H.; Wen, J.; Liao, X.; Lin, Q.; Zhang, J.; Chen, K.; Wang, S.; Zhang, J. A Sensitive, Highly Specific Novel Isothermal Amplification Method Based on Single-Nucleotide Polymorphism for the Rapid Detection of Salmonella Pullorum. Front. Microbiol. 2020, 11, 560791. [Google Scholar] [CrossRef]
- Shah, D.H.; Park, J.-H.; Cho, M.-R.; Kim, M.-C.; Chae, J.-S. Allele-Specific PCR Method Based on RfbS Sequence for Distinguishing Salmonella Gallinarum from Salmonella Pullorum: Serotype-Specific RfbS Sequence Polymorphism. J. Microbiol. Methods 2005, 60, 169–177. [Google Scholar] [CrossRef]
- Shang, Y.; Ye, Q.; Wu, Q.; Xiang, X.; Zha, F.; Du, M.; Zhang, J. Novel Multiplex PCR Assays for Rapid Identification of Salmonella Serogroups B, C1, C2, D, E, S. Enteritidis, and S. Typhimurium. Anal. Methods 2022, 14, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.R.; Shah, D.H.; Shringi, S.; Lee, M.-J.; Li, Y.-H.; Cho, M.-R.; Park, J.-H.; Eo, S.-K.; Lee, J.-H.; Chae, J.-S. An Allele-Specific PCR Assay for the Rapid and Serotype-Specific Detection of Salmonella Pullorum. Avian Dis. 2005, 49, 558–561. [Google Scholar] [CrossRef] [PubMed]
- Campioni, F.; Cao, G.; Kastanis, G.; Janies, D.A.; Bergamini, A.M.M.; dos Prazeres Rodrigues, D.; Stones, R.; Brown, E.; Allard, M.W.; Falcão, J.P. Changing of the Genomic Pattern of Salmonella Enteritidis Strains Isolated in Brazil over a 48 Year-Period Revealed by Whole Genome SNP Analyses. Sci. Rep. 2018, 8, 10478. [Google Scholar] [CrossRef] [PubMed]
- Kisand, V.; Lettieri, T. Genome Sequencing of Bacteria: Sequencing, de Novo Assembly and Rapid Analysis Using Open Source Tools. BMC Genom. 2013, 14, 211. [Google Scholar] [CrossRef] [PubMed]
- Saltykova, A.; Wuyts, V.; Mattheus, W.; Bertrand, S.; Roosens, N.H.C.; Marchal, K.; De Keersmaecker, S.C.J. Comparison of SNP-Based Subtyping Workflows for Bacterial Isolates Using WGS Data, Applied to Salmonella Enterica Serotype Typhimurium and Serotype 1,4,[5],12:I:-. PLoS One 2018, 13, e0192504. [Google Scholar] [CrossRef] [PubMed]
- Gymoese, P.; Kiil, K.; Torpdahl, M.; Østerlund, M.T.; Sørensen, G.; Olsen, J.E.; Nielsen, E.M.; Litrup, E. WGS Based Study of the Population Structure of Salmonella Enterica Serovar Infantis. BMC Genomics 2019, 20, 870. [Google Scholar] [CrossRef]
- Hu, L.; Cao, G.; Brown, E.W.; Allard, M.W.; Ma, L.M.; Zhang, G. Whole Genome Sequencing and Protein Structure Analyses of Target Genes for the Detection of Salmonella. Sci. Rep. 2021, 11, 20887. [Google Scholar] [CrossRef]
- Den Bakker, H.C.; Allard, M.W.; Bopp, D.; Brown, E.W.; Fontana, J.; Iqbal, Z.; Kinney, A.; Limberger, R.; Musser, K.A.; Shudt, M. Rapid Whole-Genome Sequencing for Surveillance of Salmonella Enterica Serovar Enteritidis. Emerg. Infect. Dis. 2014, 20, 1306. [Google Scholar] [CrossRef]
- Lampel, K.A.; Keasler, S.P.; Hanes, D.E. Specific Detection of Salmonella Enterica Serotype Enteritidis Using the Polymerase Chain Reaction. Epidemiol. Infect. 1996, 116, 137–145. [Google Scholar] [CrossRef]
- Moorhead, S.M.; Dykes, G.A.; Cursons, R.T. An SNP-Based PCR Assay to Differentiate between Listeria Monocytogenes Lineages Derived from Phylogenetic Analysis of the SigB Gene. J. Microbiol. Methods 2003, 55, 425–432. [Google Scholar] [CrossRef]
- Chen, X.; Sullivan, P.F. Single Nucleotide Polymorphism Genotyping: Biochemistry, Protocol, Cost and Throughput. Pharmacogenom. J. 2003, 3, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Andrews, W.H.; Jacobson, A.; Hammack, T. Bacteriological Analytical Manual (BAM) Chapter 5: Salmonella; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2018.
- Bäumler, A.J.; Heffron, F.; Reissbrodt, R. Rapid Detection of Salmonella Enterica with Primers Specific for IroB. J. Clin. Microbiol. 1997, 35, 1224–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferretti, R.; Mannazzu, I.; Cocolin, L.; Comi, G.; Clementi, F. Twelve-Hour PCR-Based Method for Detection of Salmonella Spp. in Food. Appl. Environ. Microbiol. 2001, 67, 977–978. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. c | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon Size (bp) | Gene (Position of SNP: Nucleotide), Flanking Sequences in between Ambiguous Code b | One-Letter Amino Acid (a.a) Code of Comparing Salmonella bongori Strains RKS3044/N268-08 -a.a Position of Ref. S. bongori NCTC 12419-a.a Code (Mutation Types) a | Respective Genes |
|---|---|---|---|---|---|---|
| 01 | GGGGAAATGTTGGCGGGA | TTATGCCCGGTGCCATGG | 735 | SBG_RS00105 (20978: G) CAACCTGCCDACCCCGATGAG | K/Y-145-D (nonsynonymous) | Conserved hypothetical protein (SBG) |
| 04 | TTGCTGGTCGCCTTCCTG | CGTATCGCGTGGCAAGGA | 863 | dedA (104317: G) TCTGGCTGGHGCCGCTATTGA | G/G-163-G (Synonymous) | DedA family integral membrane (dedA) |
| 06 | CGCGTGATGGAGCAGGAT | CCTCACACAGGCGCTGAA | 674 | yacG (146357: T) CCAGACGACBGCTTTACCACA | A/P-15-A (Synonymous) | Conserved hypothetical protein (yacG) |
| 09 | GGCGTTGAAGAAGCAGCG | ACGGCCTACCCAGGTGAT | 799 | mrcB (210347: C) CGCCAGCGGBGGAAATCGCGC. | G/G-626-G (Synonymous) | Penicillin Binding protein (mrcB) |
| 11 | TTCTGGCCAGCGACCTTG | TGCCAGTTTCAGCCACCC | 714 | mesJ (263017: G) TGAACTGCGBCAACCGCGCGC. | R/R-349-R (Synonymous) | tRNA (Ile)-lysidine synthase (mesJ) |
| 12 | ATTGGCACGCTGTCAGCT | TGCCGGTAAAAGCACGCT | 681 | metN (270836: G) CGGATCGAGGBCGCTGGTCGC. | A/A-169-A(Synonymous) | Methionine import ATP-binding protein (metN) |
| 13 | GCTGTACCTGCCGACTGG | GTTCCCCACGGGCTATGG | 797 | rihB (576128: T) GCGTATGACDCTGCAGTACG. | T/T-69-T (Synonymous) | Pyrimidine-specific ribonucleoside hydrolase (rihB) |
| 14 | TCCCCTGTGTTTCGACGC | ACGCCGGATAAGACGCTG | 682 | rihA (626519: G) CTCGGCAGCBGCGTCCAGTT. | P/P-167-P (Synonymous) | Pyrimidine-specific ribonucleoside hydrolase(rihA) |
| 15 | GCGGGAAACTCCTGTGCT | CAACACCCGGCAGCAAAC | 766 | modA (683698: C) ACTACACCGVCGCTTCATGG. | R/R-104-R (Synonymous) | Molybdate-binding periplasmic protein (modA) |
| 16 | ACGGTCTGGGTGAGGTGT | CCACCGCATCAGAACCGT | 836 | modA (747388: T) TGCGGCGGADTATAAAAAAGA. | D/D-47-D (Synonymous) | Molybdate-binding periplasmic protein (modA) |
| 18 | GCATCTGGATCTGCGCCA | TCGGCGACAAAGGTTCCC | 751 | hutG (766287: G) AATGCCGGCBTTTCCGCCCC. | A/A-252-A (Synonymous) | Formimidoylglutamase (hutG) |
| 19 | TCACGGCGGGTAAGAGGA | ATGAGATTCGCCAGGCCG | 666 | yehX (792736: T) GGCTTTGCCDAGCTGACTTT. | S/S-397-S (Synonymous) | Hypothetical ABC transporter ATP-binding (yehX) |
| 21 | CTGCTTAAACGGCGCGTC | TGGTGCGGCATGATCCTG | 738 | ybiY (827585: T) TGAGCCAGGHTGGAAAAATGG | N/Y-87-S (Nonsynonymous) | pyruvate formate-lyase 3-activating enzyme (ybiY) |
| 22 | CCGAACAGACGGCTCAGG | CCGGACATCAAGGGTCGC | 681 | moeB (828767: C) GACGCCTGCVCCGGCCAGATA | G/G-52-G (Synonymous) | Molybdopterin biosynthesis MoeB protein (moeB) |
| 24 | GTAGTGTGGCGGGCTGAG | CTGGTAAGCGTGCTGGCC | 801 | sopA (1032435: T) CTCATAAAGHGCCGCGGCTTT | A/A-494-A (Synonymous) | Candidate type three secretion system effector protein (sopA) |
| Gene | Primer | Sequence (5′-3′) # | (Mer bp) | Size (bp) | Salmonella Serovar Strains | m-PCR |
|---|---|---|---|---|---|---|
| RNA polymerase sigma factor FliA (fliA) | SBG-2F | TTACCAGGAAGAGCTCGAC | 19 | 498 | Salmonella Enteritidis | S1 |
| SBG-2R | CGGTGCCATGGCTCATCTCG | 20 | ||||
| Molybdate-binding periplasmic protein (modA) | ModA-3-F | TCGCAGGGGCGACATTATCTTCCA | 24 | 373 | Salmonella Agona | |
| ModA-3-R | AGACGAATCCAGTCCGTTTTGCTA | 24 | ||||
| E3 ubiquitin-protein ligase sopA (sopA) | SBG (2)-6F | GCTGGTTCAGCTCCCCATTA | 20 | 300 | Salmonella Abony | |
| SBG (2)-6R | CGGACTGGACAACCCGCTCC | 20 | ||||
| Penicillin binding protein (mrcB) | mrcB-1-F | TGGCGTTAGGTCTACCGTCA | 20 | 363 | Salmonella Typhi | S2 |
| mrcB-1-R | TTGTCGTCCCGGTTTTATCG | 20 | ||||
| Molybdate-binding periplasmic protein (modA) | ModA-4-F | TTACGCCTGGTCGCAGGGACA | 21 | 242 | Salmonella enterica | |
| ModA-4-R | CATTTCTGATCAGCAGAGATGGAG | 24 | ||||
| Penicillin binding protein (mrcB) | mrcB-5-F | GGCGGAGCCGCAGTATACT | 19 | 637 | Salmonella Typhimurium | |
| mrcB-5-R | TGTCGTCCCGGTTTTACTCA | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.-M.; Lim, S.-J.; Park, Y.-C. Genome-Wide Searching Single Nucleotide-Polymorphisms (SNPs) and SNPs-Targeting a Multiplex Primer for Identification of Common Salmonella Serotypes. Pathogens 2022, 11, 1075. https://doi.org/10.3390/pathogens11101075
Rahman M-M, Lim S-J, Park Y-C. Genome-Wide Searching Single Nucleotide-Polymorphisms (SNPs) and SNPs-Targeting a Multiplex Primer for Identification of Common Salmonella Serotypes. Pathogens. 2022; 11(10):1075. https://doi.org/10.3390/pathogens11101075
Chicago/Turabian StyleRahman, Md-Mafizur, Sang-Jin Lim, and Yung-Chul Park. 2022. "Genome-Wide Searching Single Nucleotide-Polymorphisms (SNPs) and SNPs-Targeting a Multiplex Primer for Identification of Common Salmonella Serotypes" Pathogens 11, no. 10: 1075. https://doi.org/10.3390/pathogens11101075
APA StyleRahman, M. -M., Lim, S. -J., & Park, Y. -C. (2022). Genome-Wide Searching Single Nucleotide-Polymorphisms (SNPs) and SNPs-Targeting a Multiplex Primer for Identification of Common Salmonella Serotypes. Pathogens, 11(10), 1075. https://doi.org/10.3390/pathogens11101075