
RT-PCR Testing of Organ Culture Medium for Corneal Storage Fails to Detect SARS-CoV-2 Infection Due to Lack of Viral Replication

 , , ,
, , ,
Abstract
:1. Introduction
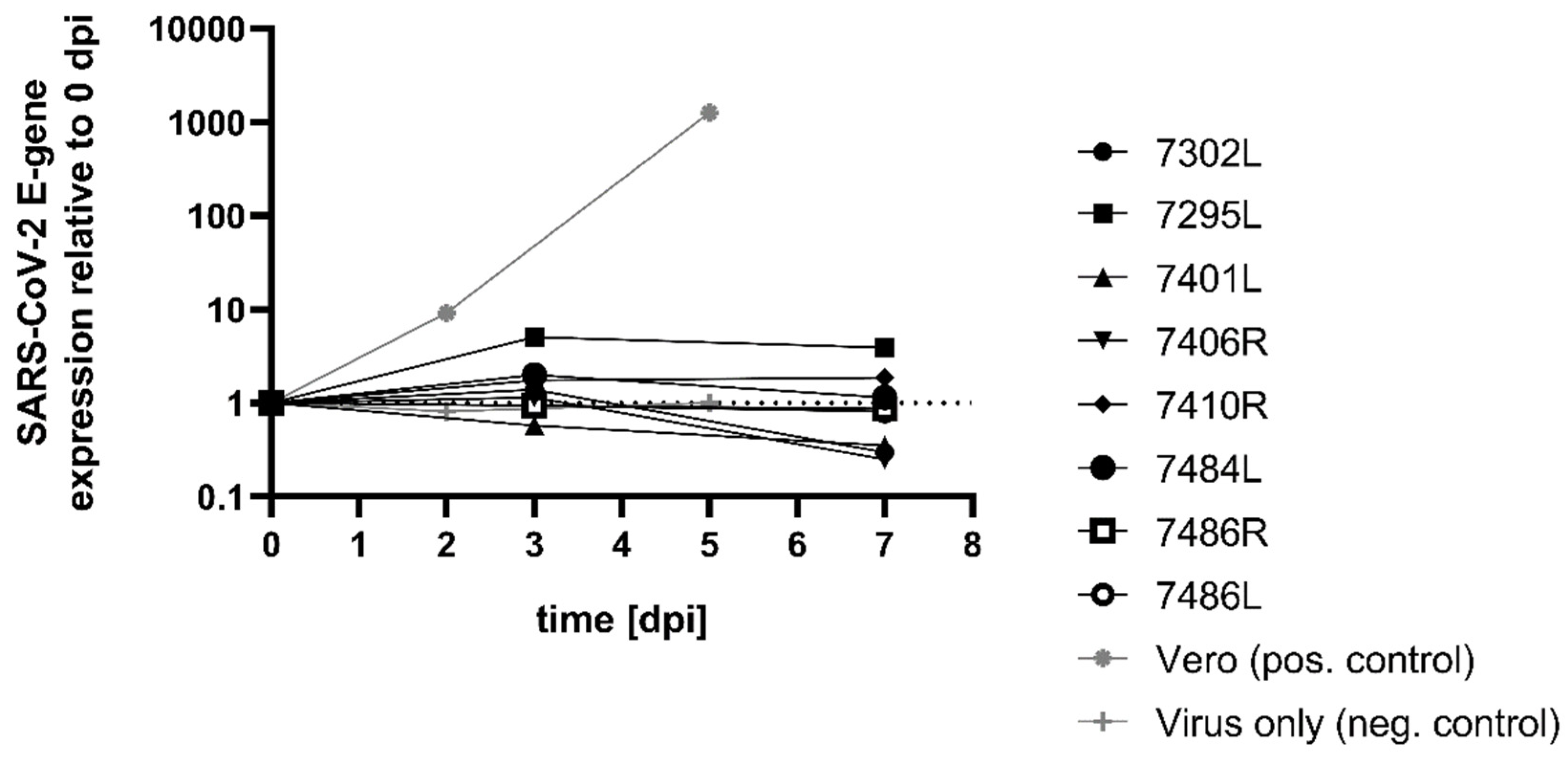
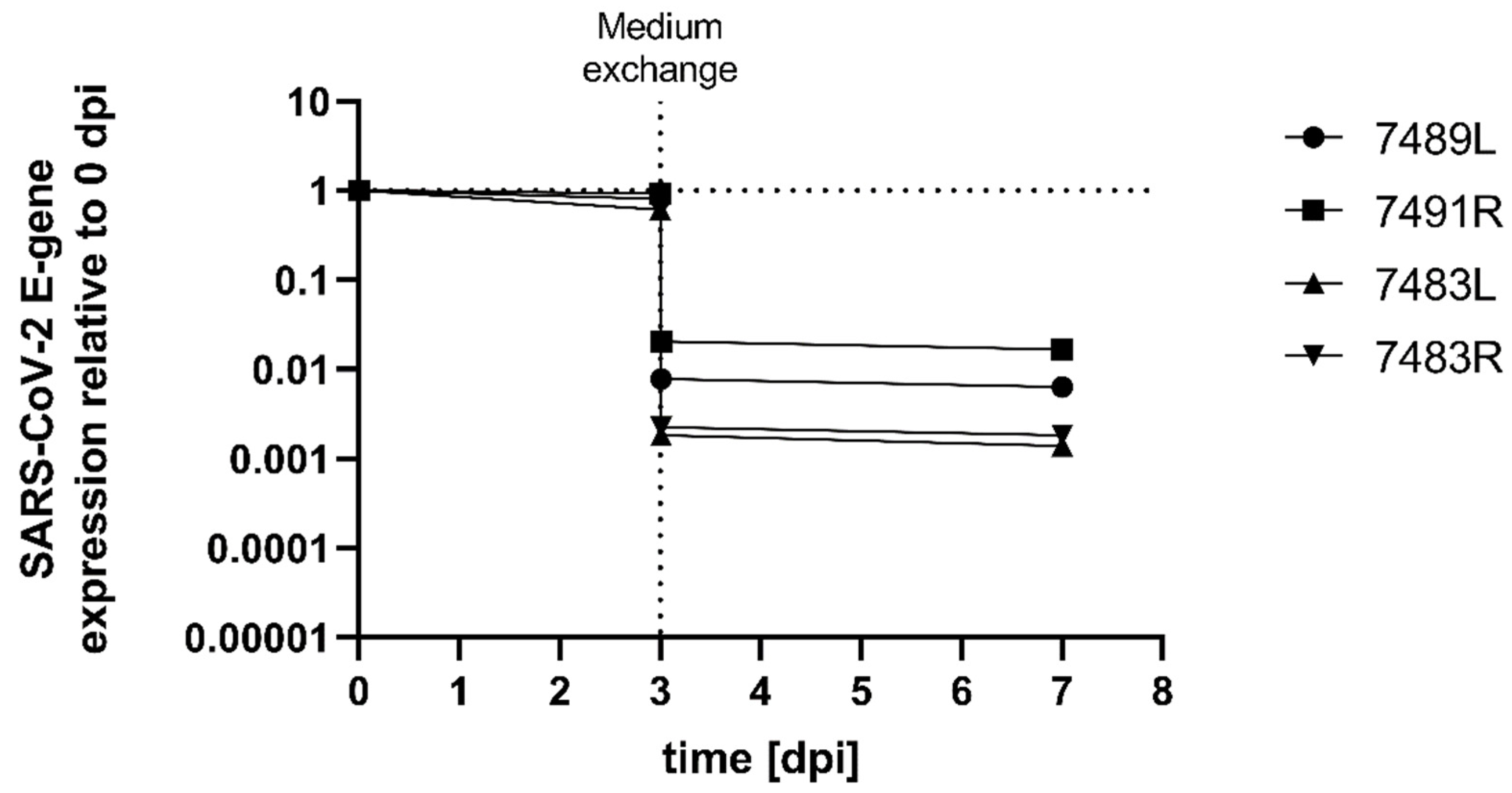
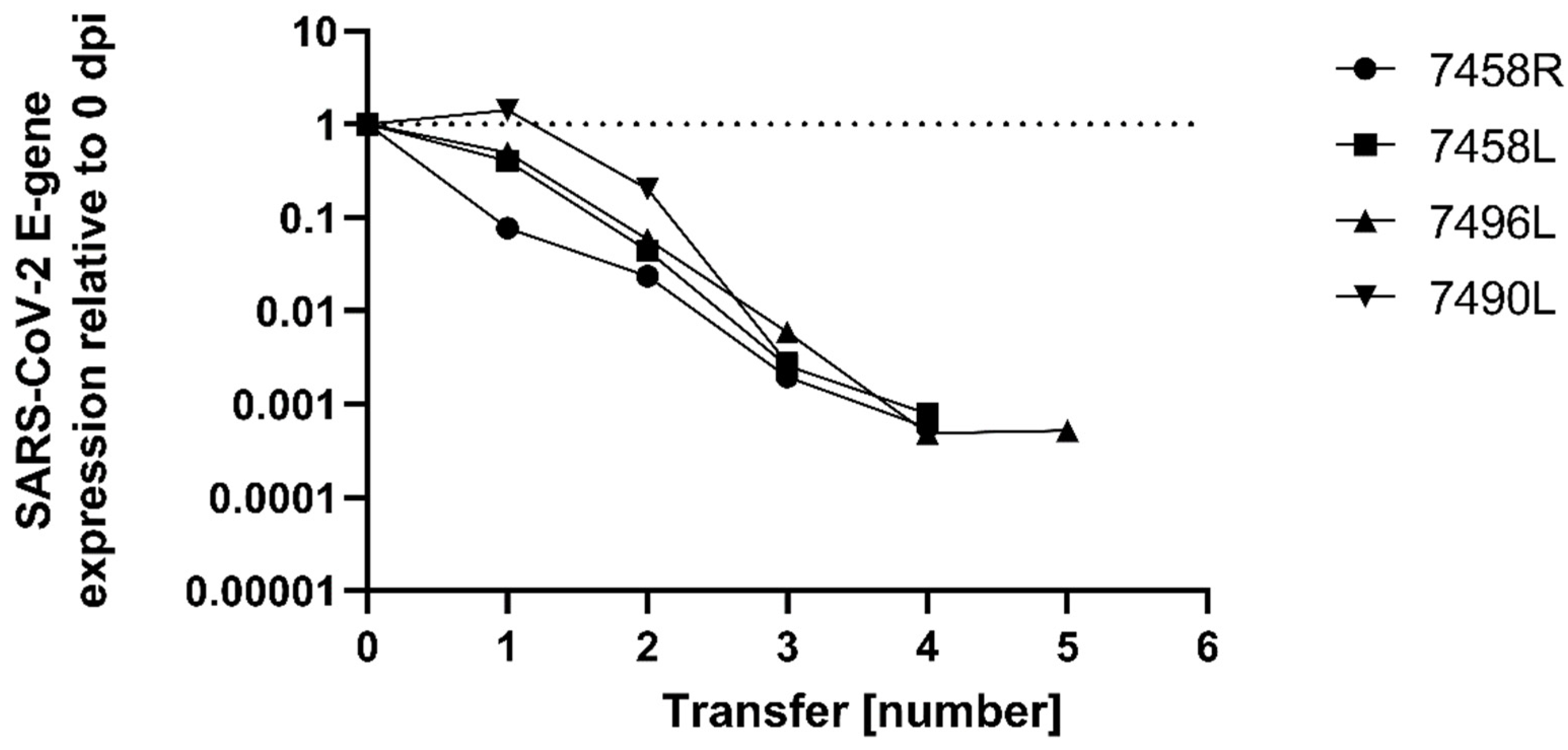
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Virus Inoculation
4.3. Supernatant Samples
4.4. Tissue Samples
4.5. qRT-PCR
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pormohammad, A.; Ghorbani, S.; Khatami, A.; Farzi, R.; Baradaran, B.; Turner, D.L.; Turner, R.J.; Bahr, N.C.; Idrovo, J.P. Comparison of confirmed COVID-19 with SARS and MERS cases—Clinical characteristics, laboratory findings, radiographic signs and outcomes: A systematic review and meta-analysis. Rev. Med. Virol. 2020, 30, e2112. [Google Scholar] [CrossRef] [PubMed]
- Keshta, A.S.; Mallah, S.I.; Al Zubaidi, K.; Ghorab, O.K.; Keshta, M.S.; Alarabi, D.; Abousaleh, M.A.; Salman, M.T.; Taha, O.E.; Zeidan, A.A.; et al. COVID-19 versus SARS: A comparative review. J. Infect. Public Health 2021, 14, 967–977. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Abdelghany, T.M.; Ganash, M.; Bakri, M.M.; Qanash, H.; Al-Rajhi, A.M.H.; Elhussieny, N.I. SARS-CoV-2, the other face to SARS-CoV and MERS-CoV: Future predictions. Biomed. J. 2021, 44, 86–93. [Google Scholar] [CrossRef]
- Ibrahim, B.; Dawson, R.; Chandler, J.A.; Goldberg, A.; Hartell, D.; Hornby, L.; Simpson, C.; Weiss, M.J.; Wilson, L.C.; Wilson, T.M.; et al. The COVID-19 pandemic and organ donation and transplantation: Ethical issues. BMC Med. Ethics 2021, 22, 142. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Roehrich, H.; Yuan, C.; Hou, J.H. Immunohistochemical Study of SARS-CoV-2 Viral Entry Factors in the Cornea and Ocular Surface. Cornea 2020, 39, 1556–1562. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, Z.; Castiglione, G.M.; Soiberman, U.S.; Eberhart, C.G.; Duh, E.J. ACE2 and TMPRSS2 are expressed on the human ocular surface, suggesting susceptibility to SARS-CoV-2 infection. Ocul. Surf. 2020, 18, 537–544. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Desautels, J.D.; Moshirfar, M.; Martheswaran, T.; Shmunes, K.M.; Ronquillo, Y.C. Risks Posed to Corneal Transplant Recipients by COVID-19-Affected Donors. Ophthalmol. Ther. 2020, 9, 371–379. [Google Scholar] [CrossRef]
- Fuest, M.; Boor, P.; Knuechel, R.; The DeRegCOVID; Walter, P.; Salla, S. Postmortem conjunctival and nasopharyngeal swabs in SARS-CoV-2 infected and uninfected patients. Acta Ophthalmol. 2021, 99, e615–e617. [Google Scholar] [CrossRef]
- Casagrande, M.; Fitzek, A.; Spitzer, M.S.; Puschel, K.; Glatzel, M.; Krasemann, S.; Norz, D.; Lutgehetmann, M.; Pfefferle, S.; Schultheiss, M. Presence of SARS-CoV-2 RNA in the Cornea of Viremic Patients with COVID-19. JAMA Ophthalmol. 2021, 139, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Sawant, O.B.; Singh, S.; Wright, R.E., 3rd; Jones, K.M.; Titus, M.S.; Dennis, E.; Hicks, E.; Majmudar, P.A.; Kumar, A.; Mian, S.I. Prevalence of SARS-CoV-2 in human post-mortem ocular tissues. Ocul. Surf. 2021, 19, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Thuret, G.; Courrier, E.; Poinard, S.; Gain, P.; Baud’Huin, M.; Martinache, I.; Cursiefen, C.; Maier, P.; Hjortdal, J.; Sanchez Ibanez, J.; et al. One threat, different answers: The impact of COVID-19 pandemic on cornea donation and donor selection across Europe. Br. J. Ophthalmol. 2020, 0, 1–7. [Google Scholar] [CrossRef]
- Trigaux, C.; Salla, S.; Schroeter, J.; Tourtas, T.; Thomasen, H.; Maier, P.; Hellwinkel, O.J.C.; Wittmershaus, I.; Merz, P.R.; Seitz, B.; et al. SARS-CoV-2: Impact on, Risk Assessment and Countermeasures in German Eye Banks. Curr. Eye Res. 2021, 46, 666–671. [Google Scholar] [CrossRef]
- dell’Omo, R.; Filippelli, M.; Virgili, G.; Bandello, F.; Querques, G.; Lanzetta, P.; Avitabile, T.; Viola, F.; Reibaldi, M.; Semeraro, F.; et al. Effect of COVID-19-related lockdown on ophthalmic practice in Italy: A report from 39 institutional centers. Eur. J. Ophthalmol. 2021, 32, 695–703. [Google Scholar] [CrossRef]
- Aiello, F.; Genzano Besso, F.; Pocobelli, G.; Gallo Afflitto, G.; Colabelli Gisoldi, R.A.M.; Nucci, C.; Ponzin, D.; Italian Society Eye Bank, G. Corneal transplant during COVID-19 pandemic: The Italian Eye Bank national report. Cell Tissue Bank. 2021, 22, 697–702. [Google Scholar] [CrossRef]
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaler, S.; Schindler, M.; Iftner, T.; Bartz-Schmidt, K.U.; Bayyoud, T. Importance of corneal organ culture in donors with possible SARS-CoV-2 infections. Ophthalmologe 2020, 117, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, F.U.; Cichutek, K.; Scriba, P.C.; Reinhard, T. Richtlinie zur Gewinnung von Spenderhornhäuten und zum Führen einer Augenhornhautbank, Erste Fortschreibung. Dtsch. Arztebl. 2018, 115, A1–A18. [Google Scholar] [CrossRef]
- Anderson, D.E.; Sivalingam, V.; Kang, A.E.Z.; Ananthanarayanan, A.; Arumugam, H.; Jenkins, T.M.; Hadjiat, Y.; Eggers, M. Povidone-Iodine Demonstrates Rapid In Vitro Virucidal Activity Against SARS-CoV-2, The Virus Causing COVID-19 Disease. Infect. Dis. Ther. 2020, 9, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Bayyoud, T.; Iftner, T.; Bartz-Schmidt, K.U.; Rohrbach, J.M.; Ueffing, M.; Schindler, M.; Thaler, S. First results of investigations of SARS-CoV2 RNA in human corneal tissue. Ophthalmologe 2020, 117, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Werner, J.; Kronberg, R.M.; Stachura, P.; Ostermann, P.N.; Muller, L.; Schaal, H.; Bhatia, S.; Kather, J.N.; Borkhardt, A.; Pandyra, A.A.; et al. Deep Transfer Learning Approach for Automatic Recognition of Drug Toxicity and Inhibition of SARS-CoV-2. Viruses 2021, 13, 610. [Google Scholar] [CrossRef]
- Ramani, A.; Muller, L.; Ostermann, P.N.; Gabriel, E.; Abida-Islam, P.; Muller-Schiffmann, A.; Mariappan, A.; Goureau, O.; Gruell, H.; Walker, A.; et al. SARS-CoV-2 targets neurons of 3D human brain organoids. EMBO J. 2020, 39, e106230. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.J.; Platt, D.J.; Ghaznavi, C.M.; Chandra, P.; Santeford, A.; Menos, A.M.; Dong, Z.; Wang, E.R.; Qian, W.; Karozichian, E.S.; et al. HSV-1 and Zika Virus but Not SARS-CoV-2 Replicate in the Human Cornea and Are Restricted by Corneal Type III Interferon. Cell Rep. 2020, 33, 108339. [Google Scholar] [CrossRef] [PubMed]
- Armitage, W.J. Preservation of Human Cornea. Transfus. Med. Hemother. 2011, 38, 143–147. [Google Scholar] [CrossRef]
- Pedrosa, C.; Goto-Silva, L.; Temerozo, J.R.; Souza, L.R.Q.; Vitoria, G.; Ornelas, I.M.; Karmirian, K.; Mendes, M.A.; Gomes, I.C.; Sacramento, C.Q.; et al. Non-permissive SARS-CoV-2 infection in human neurospheres. Stem. Cell Res. 2021, 54, 102436. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Wolf, J.; Auw-Haedrich, C.; Schlecht, A.; Boneva, S.; Lapp, T.; Horres, R.; Agostini, H.; Martin, G.; Reinhard, T.; et al. Expression of the COVID-19 receptor ACE2 in the human conjunctiva. J. Med. Virol. 2020, 92, 2081–2086. [Google Scholar] [CrossRef]
- Hui, K.P.Y.; Cheung, M.C.; Perera, R.; Ng, K.C.; Bui, C.H.T.; Ho, J.C.W.; Ng, M.M.T.; Kuok, D.I.T.; Shih, K.C.; Tsao, S.W.; et al. Tropism, replication competence, and innate immune responses of the coronavirus SARS-CoV-2 in human respiratory tract and conjunctiva: An analysis in ex-vivo and in-vitro cultures. Lancet. Respir. Med. 2020, 8, 687–695. [Google Scholar] [CrossRef]
- Kaya, H.; Caliskan, A.; Okul, M.; Sari, T.; Akbudak, I.H. Detection of SARS-CoV-2 in the tears and conjunctival secretions of Coronavirus disease 2019 patients. J. Infect. Dev. Ctries. 2020, 14, 977–981. [Google Scholar] [CrossRef]
- Xia, J.; Tong, J.; Liu, M.; Shen, Y.; Guo, D. Evaluation of coronavirus in tears and conjunctival secretions of patients with SARS-CoV-2 infection. J. Med. Virol. 2020, 92, 589–594. [Google Scholar] [CrossRef] [Green Version]
- Inomata, T.; Kitazawa, K.; Kuno, T.; Sung, J.; Nakamura, M.; Iwagami, M.; Takagi, H.; Midorikawa-Inomata, A.; Zhu, J.; Fujimoto, K.; et al. Clinical and Prodromal Ocular Symptoms in Coronavirus Disease: A Systematic Review and Meta-Analysis. Investig. Ophthalmol. Vis. Sci. 2020, 61, 29. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, C.B. Conjunctiva is not a preferred gateway of entry for SARS-CoV-2 to infect respiratory tract. J. Med. Virol. 2020, 92, 1410–1412. [Google Scholar] [CrossRef] [Green Version]
- Bhadani, P.; Agarwal, B.; Kalita, S.; Bhattacharjee, H.; Das, D.R.; Mohapatra, S.S.D. Letter to the Editor: Immunohistochemical Study of SARS-CoV-2 Viral Entry Factors in the Cornea and Ocular Surface. Cornea 2021, 40, e17–e18. [Google Scholar] [CrossRef] [PubMed]
- Aldave, A.J.; DeMatteo, J.; Chamberlain, W.D.; Philippy, B.; Farooq, A.V.; Buckman, N.; Crosson, A.; Li, J.; Meinecke, E.; Kaufman, A.H. COVID and the Cornea: From Controversies to Consensus: Report of the Eye Bank Association of America Medical Advisory Board Policy and Position Review Subcommittee. Cornea 2021, 40, 809–816. [Google Scholar] [CrossRef]
- Bayyoud, T.; Iftner, A.; Iftner, T.; Bartz-Schmidt, K.U.; Rohrbach, J.M.; Ueffing, M.; Schindler, M.; Thaler, S. Absence of Severe Acute Respiratory Syndrome-Coronavirus-2 RNA in Human Corneal Tissues. Cornea 2021, 40, 342–347. [Google Scholar] [CrossRef]
- Aiello, F.; Ciotti, M.; Gallo Afflitto, G.; Rapanotti, M.C.; Caggiano, B.; Treglia, M.; Grelli, S.; Bernardini, S.; Mauriello, S.; Nucci, C.; et al. Post-Mortem RT-PCR Assay for SARS-CoV-2 RNA in COVID-19 Patients’ Corneal Epithelium, Conjunctival and Nasopharyngeal Swabs. J. Clin. Med. 2021, 10, 4256. [Google Scholar] [CrossRef]
- Qu, J.H.; Peng, R.M.; Xiao, G.G.; Qu, H.Q.; Yu, T.; Zhang, S.; Hong, J. The incidence and influence of the donor corneas positive for herpesviridae DNA in keratoplasty. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Holland, E.J.; Bennett, S.R.; Brannian, R.; Osborne, J.C.; Goeken, J.A.; Krachmer, J.H. The risk of cytomegalovirus transmission by penetrating keratoplasty. Am. J. Ophthalmol. 1988, 105, 357–360. [Google Scholar] [CrossRef]
- Cordoba, A.; Mejia, L.F.; Mannis, M.J.; Navas, A.; Madrigal-Bustamante, J.A.; Graue-Hernandez, E.O. Current Global Bioethical Dilemmas in Corneal Transplantation. Cornea 2020, 39, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brunink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, B.; Mohammed, I.; Hopkinson, A.; Dua, H.S. Validation of endogenous control genes for gene expression studies on human ocular surface epithelium. PLoS ONE 2011, 6, e22301. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | Tissue ct-Value | Supernatant ct-Value (Inoculation) |
|---|---|---|
| 7489R | 24,0 | 25,3 |
| 7491L | 24,5 | 25,6 |
| 7401L | 30,4 | 33,0 |
| 7484L | 18,3 | 20,9 |
| 7486R | 24,8 | 21,8 |
| 7486L | 23,6 | 21,9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, L.; Ostermann, P.N.; Schaal, H.; Salla, S.; Timm, J.; Geerling, G.; Menzel-Severing, J. RT-PCR Testing of Organ Culture Medium for Corneal Storage Fails to Detect SARS-CoV-2 Infection Due to Lack of Viral Replication. Pathogens 2022, 11, 133. https://doi.org/10.3390/pathogens11020133
Müller L, Ostermann PN, Schaal H, Salla S, Timm J, Geerling G, Menzel-Severing J. RT-PCR Testing of Organ Culture Medium for Corneal Storage Fails to Detect SARS-CoV-2 Infection Due to Lack of Viral Replication. Pathogens. 2022; 11(2):133. https://doi.org/10.3390/pathogens11020133
Chicago/Turabian StyleMüller, Lisa, Philipp Niklas Ostermann, Heiner Schaal, Sabine Salla, Jörg Timm, Gerd Geerling, and Johannes Menzel-Severing. 2022. "RT-PCR Testing of Organ Culture Medium for Corneal Storage Fails to Detect SARS-CoV-2 Infection Due to Lack of Viral Replication" Pathogens 11, no. 2: 133. https://doi.org/10.3390/pathogens11020133
APA StyleMüller, L., Ostermann, P. N., Schaal, H., Salla, S., Timm, J., Geerling, G., & Menzel-Severing, J. (2022). RT-PCR Testing of Organ Culture Medium for Corneal Storage Fails to Detect SARS-CoV-2 Infection Due to Lack of Viral Replication. Pathogens, 11(2), 133. https://doi.org/10.3390/pathogens11020133