
Antibiofilm Activity of β-Lactam/β-Lactamase Inhibitor Combination against Multidrug-Resistant Salmonella Typhimurium



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Antibiotic Susceptibility of Salmonella Typhimurium Treated with Sulbactam
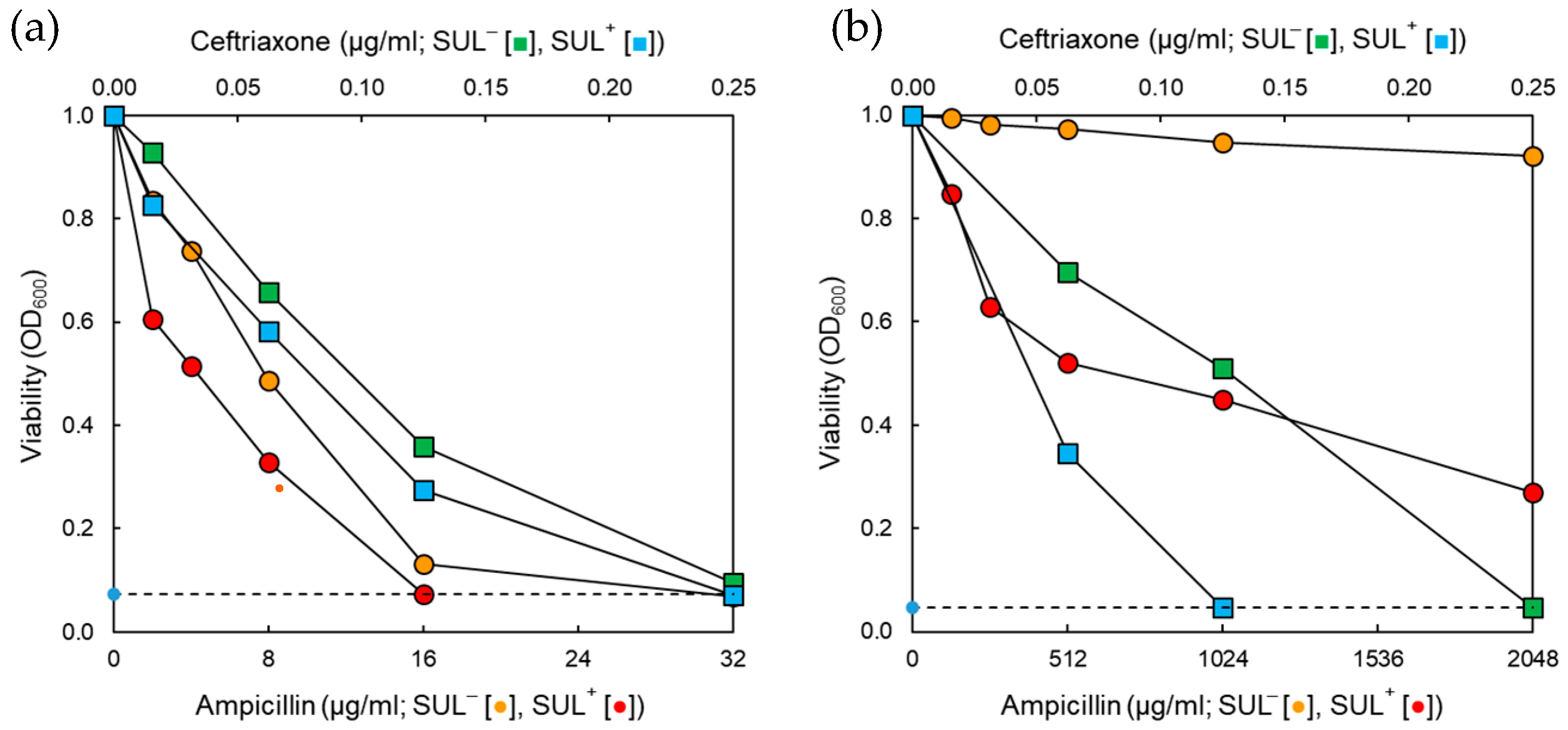
2.2. Synergistic Effect of Antibiotic and β-Lactamase Inhibitor Combinations on the Growth of Salmonella Typhimurium
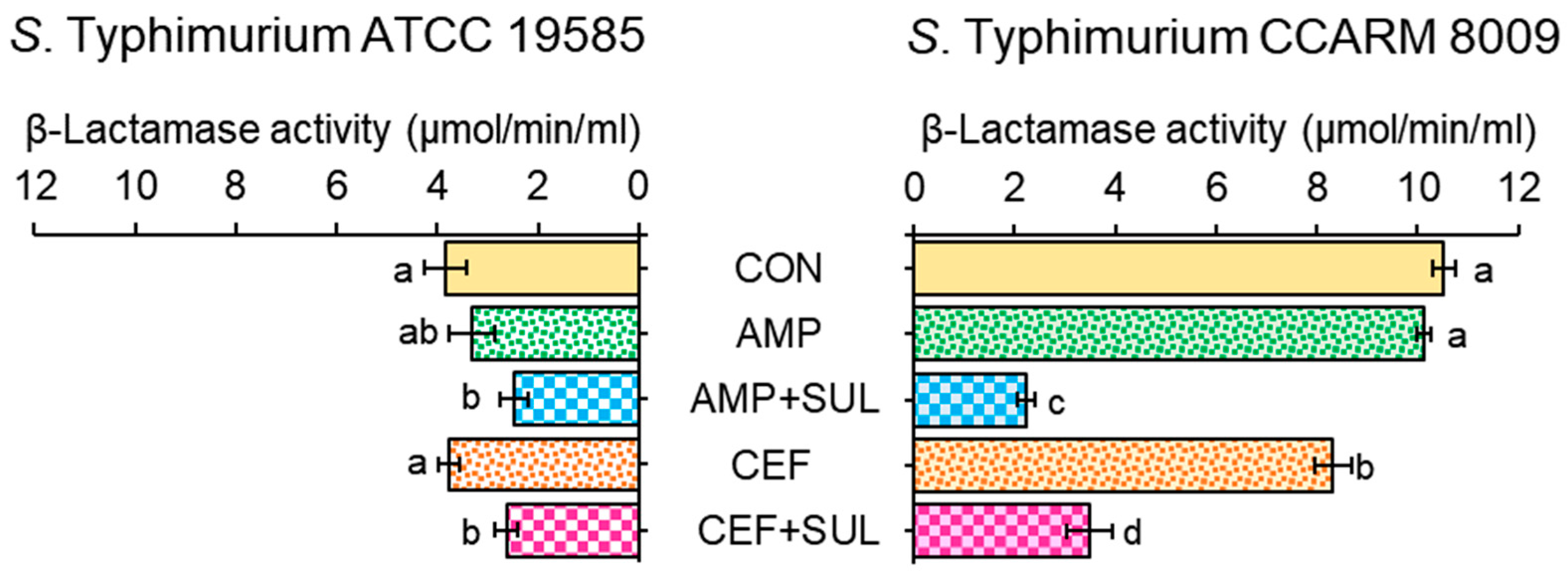
2.3. Effect of β-Lactam/β-Lactamase Inhibitor Combinations on the β-Lactamase Activity of Salmonella Typhimurium
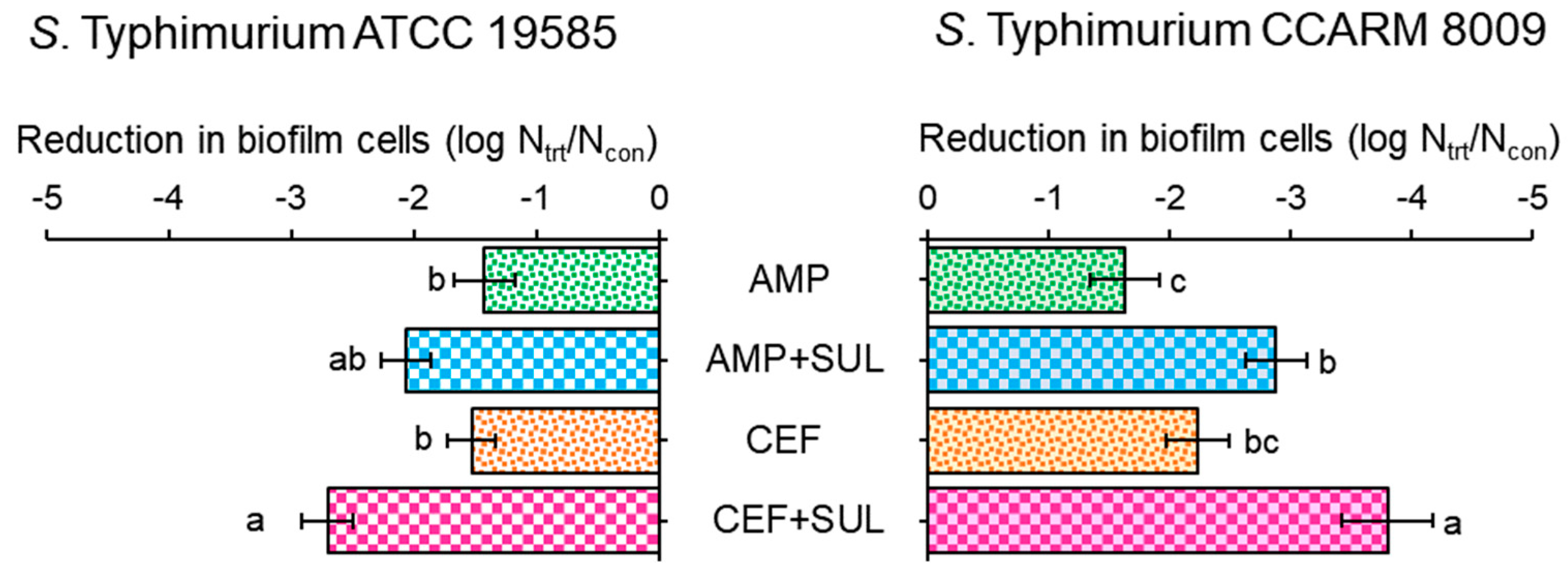
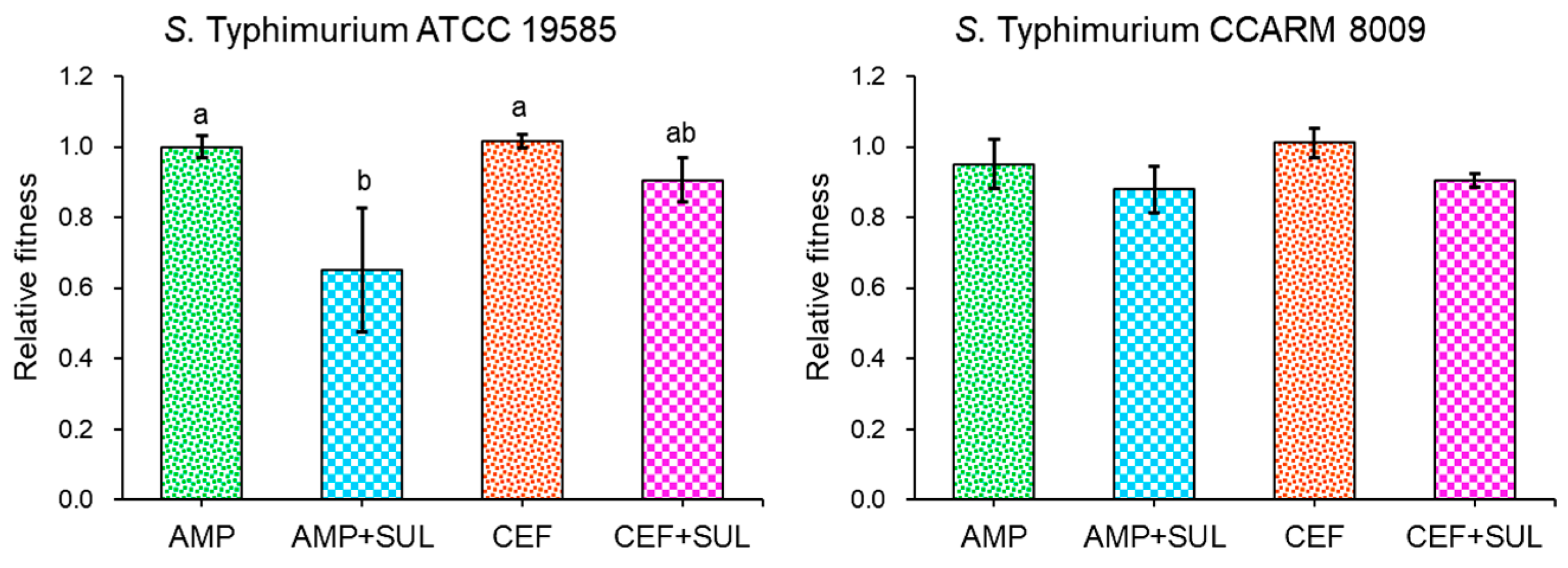
2.4. Role of β-Lactamase Inhibitor in Biofilm-Forming Ability and Relative Fitness of Salmonella Typhimurium
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Antibiotic Susceptibility Assay
4.3. β-Lactamase Activity Assay
4.4. Biofilm-Forming Ability Assay
4.5. Estimation of Relative Fitness
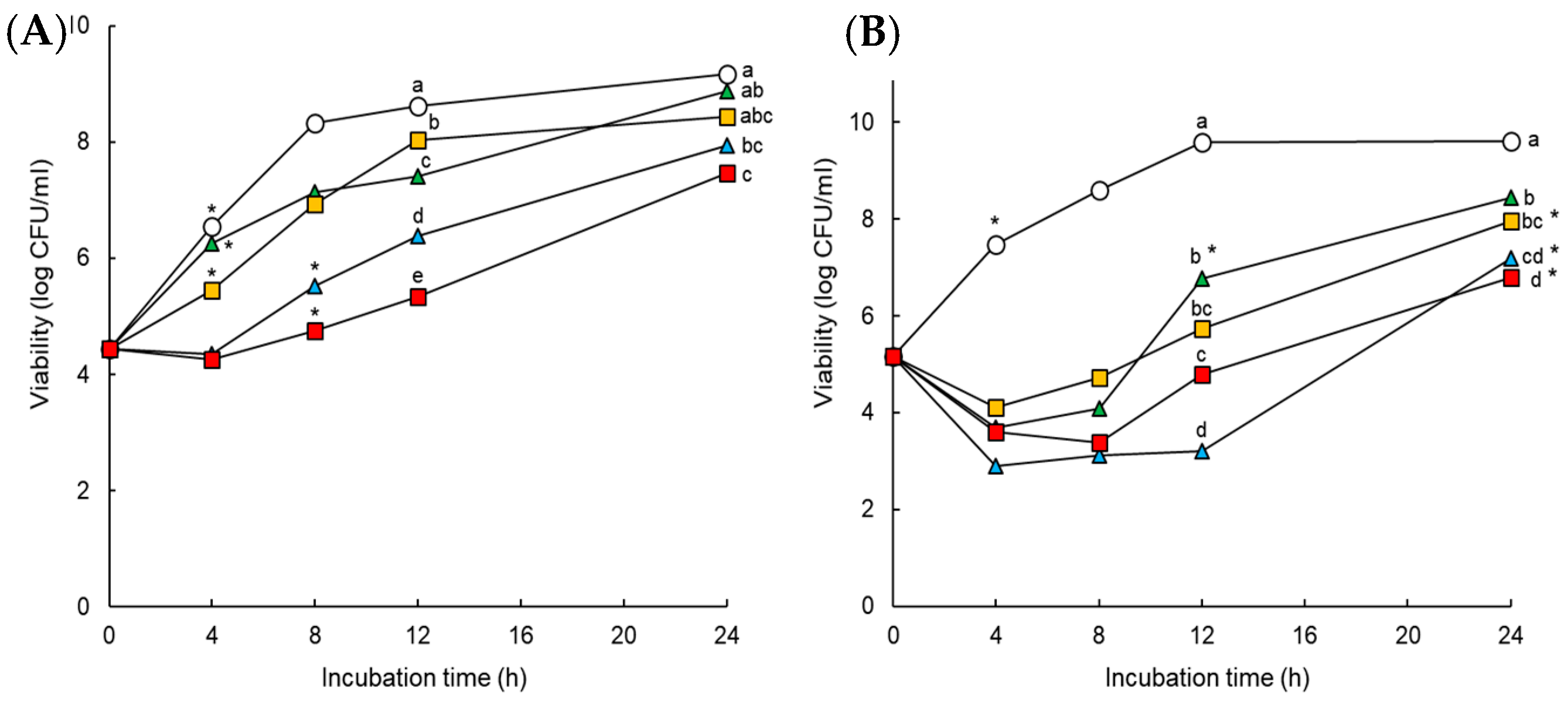
4.6. In Vitro Time-Kill Assay
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Capoor, M.R.; Nair, D. Quinolone and cephalosporin resistance in enteric fever. J. Global Infect. Dis. 2010, 2, 258. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Li, F.; Dong, N.; Tian, S.; Zhang, H.; Du, X.; Zhou, X.; Xu, X.; Yang, H.; Xie, J.; et al. Investigation of a salmonellosis outbreak caused by multidrug resistant Salmonella Typhimurium in China. Front. Microbiol. 2020, 11, 801. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Puchol, S.; Riveros, M.; Ruidias, K.; Granda, A.; Ruiz-Roldán, L.; Zapata-Cachay, C.; Ochoa, T.J.; Pons, M.J.; Ruiz, J. Dissemination of a multidrug resistant CTX-M-65 producer Salmonella enterica serovar Infantis clone between marketed chicken meat and children. Int. J. Food Microbiol. 2021, 344, 109109. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, H.; Zhang, T.; Chu, Y.; Zuo, J.; Chen, D. Study on antibiotic susceptibility of Salmonella Typhimurium L forms to the third and forth generation cephalosporins. Sci. Rep. 2020, 10, 3042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Uehara, T.; Bernhardt, T.G. Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery. Cell 2014, 159, 1300–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everaert, A.; Coenye, T. Effect of β-lactamase inhibitors on in vitro activity of β-lactam antibiotics against Burkholderia cepacia complex species. Antimicrob. Resist. Infect. Control. 2016, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Lin, J. Beta-lactamase induction and cell wall metabolism in Gram-negative bacteria. Front. Microbiol. 2013, 4, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watkins, R.; Papp-Wallace, K.; Drawz, S.; Bonomo, R. Novel β-lactamase inhibitors: A therapeutic hope against the scourge of multidrug resistance. Front. Microbiol. 2013, 4, 392. [Google Scholar] [CrossRef] [PubMed]
- Morroni, G.; Bressan, R.; Fioriti, S.; D’Achille, G.; Mingoia, M.; Cirioni, O.; Di Bella, S.; Piazza, A.; Comandatore, F.; Mauri, C. Antimicrobial activity of aztreonam in combination with old and new β-lactamase inhibitors against MBL and ESBL co-producing Gram-negative clinical isolates: Possible options for the treatment of complicated infections. Antibiotics 2021, 10, 1341. [Google Scholar] [CrossRef] [PubMed]
- Tehrani, K.H.; Martin, N.I. β-lactam/β-lactamase inhibitor combinations: An update. Medchemcomm 2018, 9, 1439–1456. [Google Scholar] [CrossRef] [PubMed]
- Drawz, S.M.; Bonomo, R.A. Three decades of β-lactamase inhibitors. Clin. Microbiol. Rev. 2010, 23, 160–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akova, M. Sulbactam-containing β-lactamase inhibitor combinations. Clin. Microbiol. Infect. 2008, 14, 185–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Bello, C.; Rodríguez, D.; Pernas, M.; Rodríguez, Á.; Colchón, E. β-lactamase inhibitors to restore the efficacy of antibiotics against superbugs. J. Med. Chem. 2019, 63, 1859–1881. [Google Scholar] [CrossRef] [PubMed]
- Mallik, D.; Pal, S.; Ghosh, A.S. Involvement of AmpG in mediating a dynamic relationship between serine beta-lactamase induction and biofilm-forming ability of Escherichia coli. FEMS Microbiol. Lett. 2018, 365, fny065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Chen, Y.; Han, R.; Huang, Z.; Zhang, X.; Hu, F.; Yang, F. Sulbactam enhances in vitro activity of β-lactam antibiotics against Acinetobacter baumannii. Infect. Drug Resist. 2021, 14, 3971. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.-K.; Pusparajah, P.; Ab Mutalib, N.-S.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Al kraiem, A.A.; Yang, G.; Al kraiem, F.; Chen, T. Challenges associated with ceftriaxone resistance in Salmonella. Front. Life Sci. 2018, 11, 26–34. [Google Scholar] [CrossRef]
- Liu, X.; Zheng, H.; Zhang, W.; Shen, Z.; Zhao, M.; Chen, Y.; Sun, L.; Shi, J.; Zhang, J. Tracking cefoperazone/sulbactam resistance development in vivo in A. baumannii isolated from a patient with hospital-acquired pneumonia by whole-genome sequencing. Front. Microbiol. 2016, 7, 1268. [Google Scholar] [CrossRef] [PubMed]
- Penwell, W.F.; Shapiro, A.B.; Giacobbe, R.A.; Gu, R.F.; Gao, N.; Thresher, J.; McLaughlin, R.E.; Huband, M.D.; DeJonge, B.L.; Ehmann, D.E.; et al. Molecular mechanisms of sulbactam antibacterial activity and resistance determinants in Acinetobacter baumannii. Antimicrob Agents Chemother 2015, 59, 1680–1689. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.-D.; Lin, M.-L.; Lee, W.-S.; Liu, C.-Y. In vitro activities of β-lactam antibiotics alone and in combination with sulbactam against Gram-negative bacteria. Int. J. Antimicrob. Agents 2004, 23, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Selmer, M.; Andersson, D.I. Resistance to β-lactam antibiotics conferred by point mutations in penicillin-binding proteins PBP3, PBP4 and PBP6 in Salmonella enterica. PLoS ONE 2014, 9, e97202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Carcione, D.; Siracusa, C.; Sulejmani, A.; Leoni, V.; Intra, J. Old and new beta-lactamase inhibitors: Molecular structure, mechanism of action, and clinical Use. Antibiotics 2021, 10, 995. [Google Scholar] [CrossRef] [PubMed]
- Vivas, M.C.; Caicedo, L.D.; Castillo, J.E. Occurrence of β-lactamase-producing Gram-negative bacterial isolates in water sources in Cali City, Colombia. Int. J. Microbiol. 2019, 2019, 1375060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Chen, Q.; Yu, X.; Li, Q.; Ding, B.; Yang, L.; Chen, C.; Qin, Z.; Parsons, C.; Zhang, X. High prevalence of extended-spectrum beta lactamases among Salmonella enterica Typhimurium isolates from pediatric patients with diarrhea in China. PLoS ONE 2011, 6, e16801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguale, T.; Birungi, J.; Asrat, D.; Njahira, M.N.; Njuguna, J.; Gebreyes, W.A.; Gunn, J.S.; Djikeng, A.; Engidawork, E. Genetic markers associated with resistance to beta-lactam and quinolone antimicrobials in non-typhoidal Salmonella isolates from humans and animals in central Ethiopia. Antimicrob. Resist. Infect. Control. 2017, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, A.B. Kinetics of sulbactam hydrolysis by β-lactamases, and kinetics of β-lactamase inhibition by sulbactam. Antimicrob. Agents Chemother. 2017, 61, e01612–e01617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, M.; McCusker, M.; Amaral, L.; Fanning, S. Mechanisms of antibiotic resistance in Salmonella: Efflux pumps, genetics, quorum sensing and biofilm formation. Lett. Drug Des. Discov. 2011, 8, 114–123. [Google Scholar] [CrossRef]
- Jing, W.; Liu, J.; Wu, S.; Li, X.; Liu, Y. Role of cpxA mutations in the resistance to aminoglycosides and β-lactams in Salmonella enterica serovar Typhimurium. Front. Microbiol. 2021, 12, 604079. [Google Scholar] [CrossRef] [PubMed]
- Kakasis, A.; Panitsa, G. Bacteriophage therapy as an alternative treatment for human infections. A comprehensive review. Int. J. Antimicrob. Agents 2019, 53, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Wenjie, Y.; Ping, L.; Na, W.; Haixia, R.; Xuequn, Z. Antibiotic resistance of Klebsiella pneumoniae through β-arrestin recruitment-induced β-lactamase signaling pathway. Exp. Ther. Med. 2018, 15, 2247–2254. [Google Scholar] [PubMed]
- Neupane, S.; Pant, N.D.; Khatiwada, S.; Chaudhary, R.; Banjara, M.R. Correlation between biofilm formation and resistance toward different commonly used antibiotics along with extended spectrum beta lactamase production in uropathogenic Escherichia coli isolated from the patients suspected of urinary tract infections visiting Shree Birendra Hospital, Chhauni, Kathmandu, Nepal. Antimicrob. Resist. Infect. Control. 2016, 5, 1–5. [Google Scholar]
- Kaplan, J.B.; Izano, E.A.; Gopal, P.; Karwacki, M.T.; Kim, S.; Bose, J.L.; Bayles, K.W.; Horswill, A.R. Low levels of β-lactam antibiotics induce extracellular DNA release and biofilm formation in Staphylococcus aureus. MBio 2012, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peppoloni, S.; Pericolini, E.; Colombari, B.; Pinetti, D.; Cermelli, C.; Fini, F.; Prati, F.; Caselli, E.; Blasi, E. The β-lactamase inhibitor boronic acid derivative SM23 as a new anti-Pseudomonas aeruginosa biofilm. Front. Microbiol. 2020, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunai, A.; Spohn, R.; Farkas, Z.; Lázár, V.; Györkei, Á.; Apjok, G.; Boross, G.; Szappanos, B.; Grézal, G.; Faragó, A. Rapid decline of bacterial drug-resistance in an antibiotic-free environment through phenotypic reversion. Elife 2019, 8, e47088. [Google Scholar] [CrossRef]
- Dawan, J.; Ahn, J. Effectiveness of antibiotic combination treatments to control heteroresistant Salmonella Typhimurium. Microb. Drug Resist. 2021, 27, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Besier, S.; Ludwig, A.; Brade, V.; Wichelhaus, T.A. Compensatory adaptation to the loss of biological fitness associated with acquisition of fusidic acid resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- O’Regan, E.; Quinn, T.; Frye, J.G.; Pagès, J.-M.; Porwollik, S.; Fedorka-Cray, P.J.; McClelland, M.; Fanning, S. Fitness costs and stability of a high-level ciprofloxacin resistance phenotype in Salmonella enterica serotype enteritidis: Reduced infectivity associated with decreased expression of Salmonella pathogenicity island 1 genes. Antimicrob. Agents Chemother. 2010, 54, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Starikova, I.; Harms, K.; Haugen, P.; Lunde, T.T.; Primicerio, R.; Samuelsen, Ø.; Nielsen, K.M.; Johnsen, P.J. A trade-off between the fitness cost of functional integrases and long-term stability of integrons. PLoS Pathog. 2012, 8, e1003043. [Google Scholar] [CrossRef] [Green Version]
- Maisnier-Patin, S.; Andersson, D.I. Adaptation to the deleterious effects of antimicrobial drug resistance mutations by compensatory evolution. Res. Microbiol. 2004, 155, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Ramnani, P.; Virdi, J.S. Detection and assay of β-lactamases in clinical and non-clinical strains of Yersinia enterocolitica biovar 1A. J. Antimicrob. Chemother. 2004, 54, 401–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laure, N.N.; Ahn, J. Antibiofilm Activity of β-Lactam/β-Lactamase Inhibitor Combination against Multidrug-Resistant Salmonella Typhimurium. Pathogens 2022, 11, 349. https://doi.org/10.3390/pathogens11030349
Laure NN, Ahn J. Antibiofilm Activity of β-Lactam/β-Lactamase Inhibitor Combination against Multidrug-Resistant Salmonella Typhimurium. Pathogens. 2022; 11(3):349. https://doi.org/10.3390/pathogens11030349
Chicago/Turabian StyleLaure, Nana Nguefang, and Juhee Ahn. 2022. "Antibiofilm Activity of β-Lactam/β-Lactamase Inhibitor Combination against Multidrug-Resistant Salmonella Typhimurium" Pathogens 11, no. 3: 349. https://doi.org/10.3390/pathogens11030349
APA StyleLaure, N. N., & Ahn, J. (2022). Antibiofilm Activity of β-Lactam/β-Lactamase Inhibitor Combination against Multidrug-Resistant Salmonella Typhimurium. Pathogens, 11(3), 349. https://doi.org/10.3390/pathogens11030349