
Spectrum of Atazanavir-Selected Protease Inhibitor-Resistance Mutations

 ,
,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Studies
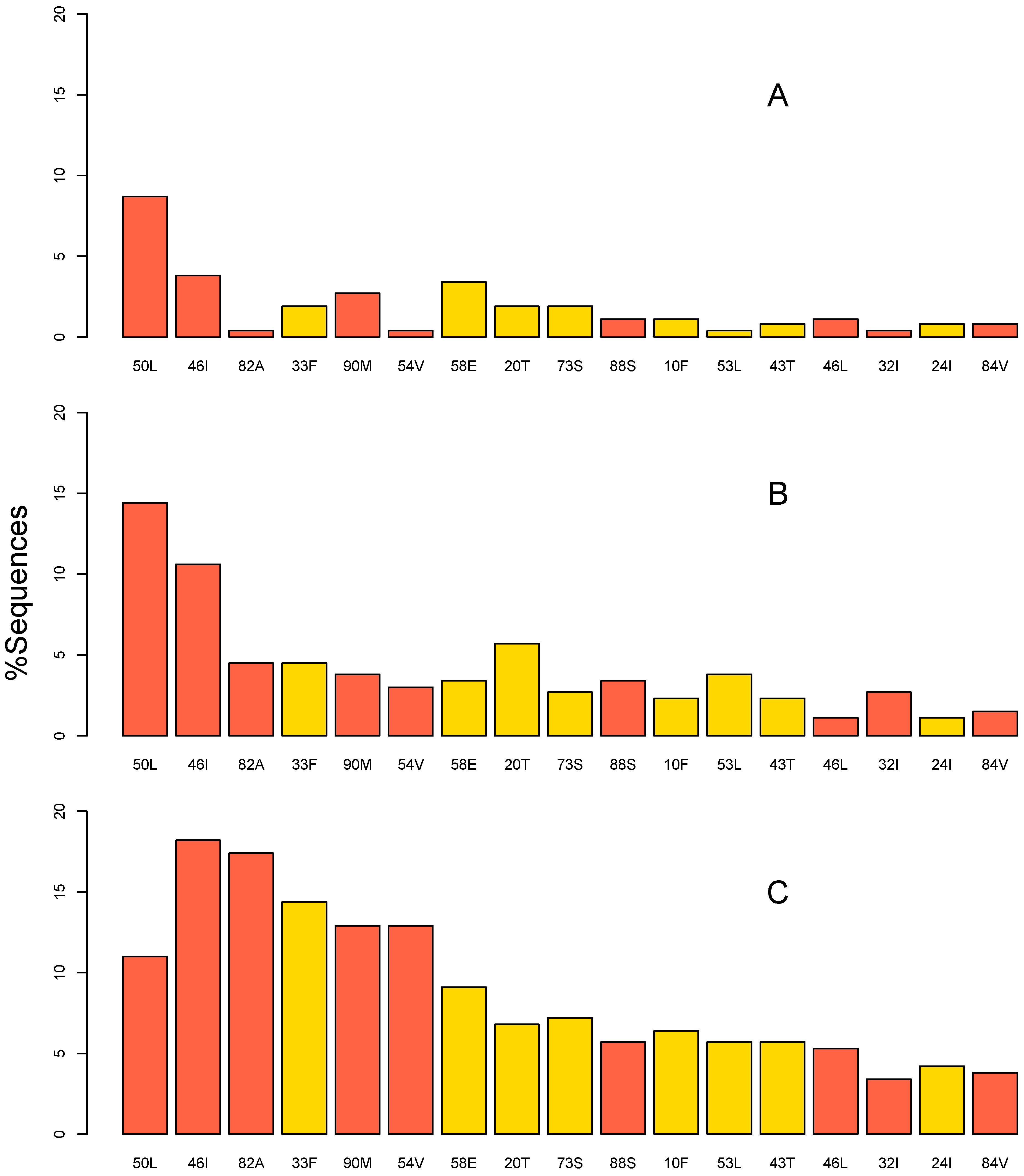
2.2. Mutation Prevalence
2.3. Unboosted versus Boosted Atazanavir
2.4. Subtypes
2.5. ART Experience
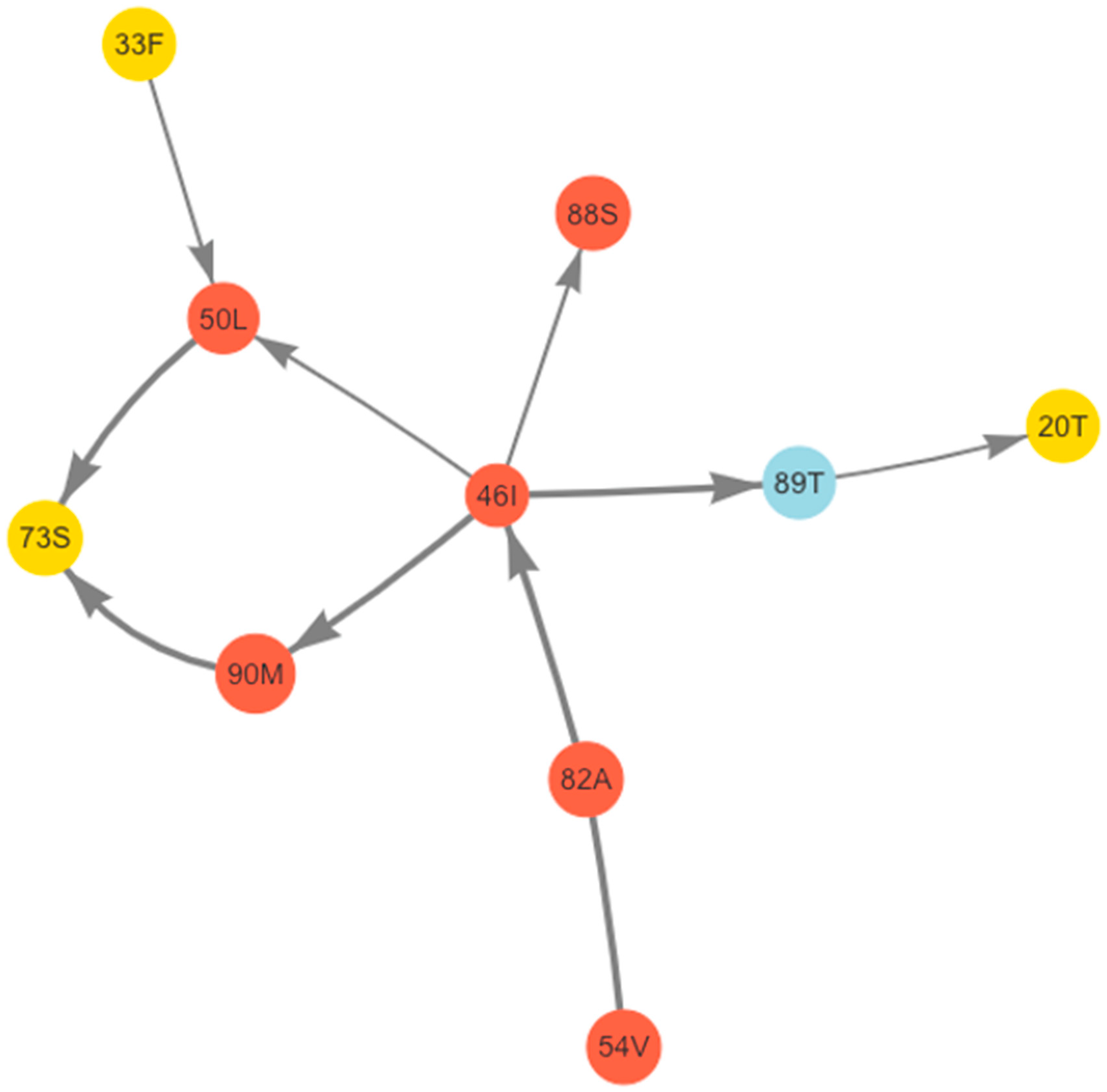
2.6. Bayesian Network Analysis of Correlated Mutations
2.7. Estimated Cross Resistance to LPV/r and DRV/r
2.8. Virological Failure with Resistance
2.9. Studies Not Included in the Analysis
3. Discussion
4. Materials and Methods
4.1. Study Selection Criteria
4.2. Mutations
4.3. Analyses
4.4. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection. 2016. Available online: https://www.who.int/publications-detail-redirect/9789241549684 (accessed on 10 February 2022).
- Laker, E.A.O.; Nabaggala, M.S.; Kaimal, A.; Nalwanga, D.; Castelnuovo, B.; Musubire, A.; Kiragga, A.; Lamorde, M.; Ratanshi, R.P. An Observational Study in an Urban Ugandan Clinic Comparing Virological Outcomes of Patients Switched from First-Line Antiretroviral Regimens to Second-Line Regimens Containing Ritonavir-Boosted Atazanavir or Ritonavir-Boosted Lopinavir. BMC Infect. Dis. 2019, 19, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engamba, D.C.; Wester, C.W.; Mutinta, M.M.; Kumar, A.P.; Chirwa, B.; Phiri, G.; Sinkala, E.; Kampamba, D.; Mpanji, S.; Mbewe, N.; et al. Predictors of Viral Suppression Following Enhanced Adherence Counseling: VISEND Trial. In Proceedings of the Conference on Retroviruses and Opportunistic Infections, Virtual, 12–16 February 2022; p. 490. [Google Scholar]
- Barber, T.J.; Harrison, L.; Asboe, D.; Williams, I.; Kirk, S.; Gilson, R.; Bansi, L.; Pillay, D.; Dunn, D.; UK HIV Drug Resistance Database and UK Collaborative HIV Cohort (UK CHIC) Study Steering Committees. Frequency and Patterns of Protease Gene Resistance Mutations in HIV-Infected Patients Treated with Lopinavir/Ritonavir as Their First Protease Inhibitor. J. Antimicrob. Chemother. 2012, 67, 995–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Zyl, G.U.; Liu, T.F.; Claassen, M.; Engelbrecht, S.; de Oliveira, T.; Preiser, W.; Wood, N.T.; Travers, S.; Shafer, R.W. Trends in Genotypic HIV-1 Antiretroviral Resistance between 2006 and 2012 in South African Patients Receiving First- and Second-Line Antiretroviral Treatment Regimens. PLoS ONE 2013, 8, e67188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawizza, H.E.; Chaplin, B.; Meloni, S.T.; Darin, K.M.; Olaitan, O.; Scarsi, K.K.; Onwuamah, C.K.; Audu, R.A.; Chebu, P.R.; Imade, G.E.; et al. Accumulation of Protease Mutations among Patients Failing Second-Line Antiretroviral Therapy and Response to Salvage Therapy in Nigeria. PLoS ONE 2013, 8, e73582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerrienet, E. HIV-1 Protease Inhibitors Resistance Profiles in Patients with Virological Failure on LPV/r-Based 2nd Line Regimen in Cambodia. J. AIDS Clin. Res. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Grossman, Z.; Schapiro, J.M.; Levy, I.; Elbirt, D.; Chowers, M.; Riesenberg, K.; Olstein-Pops, K.; Shahar, E.; Istomin, V.; Asher, I.; et al. Comparable Long-Term Efficacy of Lopinavir/Ritonavir and Similar Drug-Resistance Profiles in Different HIV-1 Subtypes. PLoS ONE 2014, 9, e86239. [Google Scholar] [CrossRef]
- Steegen, K.; Bronze, M.; Papathanasopoulos, M.A.; van Zyl, G.; Goedhals, D.; Van Vuuren, C.; Macleod, W.; Sanne, I.; Stevens, W.S.; Carmona, S.C. Prevalence of Antiretroviral Drug Resistance in Patients Who Are Not Responding to Protease Inhibitor-Based Treatment: Results From the First National Survey in South Africa. J. Infect. Dis. 2016, 214, 1826–1830. [Google Scholar] [CrossRef]
- De Faria Romero Soldi, G.; Ribeiro, I.C.; Ahagon, C.M.; Coelho, L.P.O.; Cabral, G.B.; Lopes, G.I.S.L.; de Paula Ferreira, J.L.; de Macedo Brígido, L.F.; Salvage Therapy Working Group. Major Drug Resistance Mutations to HIV-1 Protease Inhibitors (PI) among Patients Exposed to PI Class Failing Antiretroviral Therapy in São Paulo State, Brazil. PLoS ONE 2019, 14, e0223210. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.A.; Kityo, C.; Dunn, D.; Hoppe, A.; Ndashimye, E.; Hakim, J.; Kambugu, A.; van Oosterhout, J.J.; Arribas, J.; Mugyenyi, P.; et al. Evolution of Protease Inhibitor Resistance in Human Immunodeficiency Virus Type 1 Infected Patients Failing Protease Inhibitor Monotherapy as Second-Line Therapy in Low-Income Countries: An Observational Analysis Within the EARNEST Randomized Trial. Clin. Infect. Dis. 2019, 68, 1184–1192. [Google Scholar] [CrossRef]
- Posada-Céspedes, S.; Van Zyl, G.; Montazeri, H.; Kuipers, J.; Rhee, S.-Y.; Kouyos, R.; Günthard, H.F.; Beerenwinkel, N. Comparing Mutational Pathways to Lopinavir Resistance in HIV-1 Subtypes B versus C. PLoS Comput. Biol. 2021, 17, e1008363. [Google Scholar] [CrossRef]
- Rhee, S.-Y.; Gonzales, M.J.; Kantor, R.; Betts, B.J.; Ravela, J.; Shafer, R.W. Human Immunodeficiency Virus Reverse Transcriptase and Protease Sequence Database. Nucleic Acids Res. 2003, 31, 298–303. [Google Scholar] [CrossRef] [Green Version]
- EuResist Network|Research in HIV|HIV Resistance Database. Available online: https://www.euresist.org (accessed on 1 March 2022).
- Tarasova, O.; Rudik, A.; Kireev, D.; Poroikov, V. RHIVDB: A Freely Accessible Database of HIV Amino Acid Sequences and Clinical Data of Infected Patients. Front. Genet. 2021, 12, 679029. [Google Scholar] [CrossRef] [PubMed]
- Mollan, K.; Daar, E.S.; Sax, P.E.; Balamane, M.; Collier, A.C.; Fischl, M.A.; Lalama, C.M.; Bosch, R.J.; Tierney, C.; Katzenstein, D.; et al. HIV-1 Amino Acid Changes among Participants with Virologic Failure: Associations with First-Line Efavirenz or Atazanavir plus Ritonavir and Disease Status. J. Infect. Dis. 2012, 206, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Kantor, R.; Smeaton, L.; Vardhanabhuti, S.; Hudelson, S.E.; Wallis, C.L.; Tripathy, S.; Morgado, M.G.; Saravanan, S.; Balakrishnan, P.; Reitsma, M.; et al. Pretreatment HIV Drug Resistance and HIV-1 Subtype C Are Independently Associated with Virologic Failure: Results from the Multinational PEARLS (ACTG A5175) Clinical Trial. Clin. Infect. Dis. 2015, 60, 1541–1549. [Google Scholar] [CrossRef] [Green Version]
- Lennox, J.L.; Landovitz, R.J.; Ribaudo, H.J.; Ofotokun, I.; Na, L.H.; Godfrey, C.; Kuritzkes, D.R.; Sagar, M.; Brown, T.T.; Cohn, S.E.; et al. Efficacy and Tolerability of 3 Nonnucleoside Reverse Transcriptase Inhibitor-Sparing Antiretroviral Regimens for Treatment-Naive Volunteers Infected with HIV-1: A Randomized, Controlled Equivalence Trial. Ann. Intern. Med. 2014, 161, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Kouamou, V.; Manasa, J.; Katzenstein, D.; McGregor, A.M.; Ndhlovu, C.E.; Makadzange, A.T. Drug Resistance and Optimizing Dolutegravir Regimens for Adolescents and Young Adults Failing Antiretroviral Therapy. AIDS 2019, 33, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Lima, E.N.; Lima, R.S.A.; Piqueira, J.R.C.; Sucupira, M.C.; Camargo, M.; Galinskas, J.; Diaz, R.S. Evidence of Genomic Information and Structural Restrictions of HIV-1 PR and RT Gene Regions from Individuals Experiencing Antiretroviral Virologic Failure. Infect. Genet. Evol. 2020, 78, 104134. [Google Scholar] [CrossRef] [PubMed]
- Acharya, A.; Vaniawala, S.; Shah, P.; Misra, R.N.; Wani, M.; Mukhopadhyaya, P.N. Development, Validation and Clinical Evaluation of a Low Cost in-House HIV-1 Drug Resistance Genotyping Assay for Indian Patients. PLoS ONE 2014, 9, e105790. [Google Scholar] [CrossRef] [Green Version]
- Ndashimye, E.; Avino, M.; Kyeyune, F.; Nankya, I.; Gibson, R.M.; Nabulime, E.; Poon, A.F.Y.; Kityo, C.; Mugyenyi, P.; Quiñones-Mateu, M.E.; et al. Absence of HIV-1 Drug Resistance Mutations Supports the Use of Dolutegravir in Uganda. AIDS Res. Hum. Retrovir. 2018, 34, 404–414. [Google Scholar] [CrossRef]
- Gulick, R.M.; Ribaudo, H.J.; Shikuma, C.M.; Lustgarten, S.; Squires, K.E.; Meyer, W.A.; Acosta, E.P.; Schackman, B.R.; Pilcher, C.D.; Murphy, R.L.; et al. Triple-Nucleoside Regimens versus Efavirenz-Containing Regimens for the Initial Treatment of HIV-1 Infection. N. Engl. J. Med. 2004, 350, 1850–1861. [Google Scholar] [CrossRef]
- Colonno, R.; Rose, R.; McLaren, C.; Thiry, A.; Parkin, N.; Friborg, J. Identification of I50L as the Signature Atazanavir (ATV)-Resistance Mutation in Treatment-Naive HIV-1-Infected Patients Receiving ATV-Containing Regimens. J. Infect. Dis. 2004, 189, 1802–1810. [Google Scholar] [CrossRef] [PubMed]
- Chimukangara, B.; Varyani, B.; Shamu, T.; Mutsvangwa, J.; Manasa, J.; White, E.; Chimbetete, C.; Luethy, R.; Katzenstein, D. HIV Drug Resistance Testing among Patients Failing Second Line Antiretroviral Therapy. Comparison of in-House and Commercial Sequencing. J. Virol. Methods 2017, 243, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makwaga, O.; Adhiambo, M.; Mulama, D.H.; Muoma, J.; Adungo, F.; Wanjiku, H.; Ongaya, A.; Maitha, G.M.; Mwau, M. Prevalence of Human Immunodeficiency Virus-1 Drug-Resistant Mutations among Adults on First- and Second-Line Antiretroviral Therapy in a Resource-Limited Health Facility in Busia County, Kenya. Pan Afr. Med. J. 2020, 37, 311. [Google Scholar] [CrossRef] [PubMed]
- De Sa-Filho, D.J.; da Silva Soares, M.; Candido, V.; Gagliani, L.H.; Cavaliere, E.; Diaz, R.S.; Caseiro, M.M. HIV Type 1 Pol Gene Diversity and Antiretroviral Drug Resistance Mutations in Santos, Brazil. AIDS Res. Hum. Retrovir. 2008, 24, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Kolomeets, A.N.; Varghese, V.; Lemey, P.; Bobkova, M.R.; Shafer, R.W. A Uniquely Prevalent Nonnucleoside Reverse Transcriptase Inhibitor Resistance Mutation in Russian Subtype A HIV-1 Viruses. AIDS 2014, 28, F1–F8. [Google Scholar] [CrossRef] [Green Version]
- Alves, B.M.; Siqueira, J.D.; Prellwitz, I.M.; Botelho, O.M.; Da Hora, V.P.; Sanabani, S.; Recordon-Pinson, P.; Fleury, H.; Soares, E.A.; Soares, M.A. Estimating HIV-1 Genetic Diversity in Brazil through Next-Generation Sequencing. Front. Microbiol. 2019, 10, 749. [Google Scholar] [CrossRef]
- Kim, M.H.; Song, J.E.; Ahn, J.Y.; Kim, Y.C.; Oh, D.H.; Choi, H.; Ann, H.W.; Kim, J.K.; Kim, S.B.; Jeong, S.J.; et al. HIV Antiretroviral Resistance Mutations among Antiretroviral Treatment-Naive and -Experienced Patients in South Korea. AIDS Res. Hum. Retrovir. 2013, 29, 1617–1620. [Google Scholar] [CrossRef] [Green Version]
- Karkashadze, E.; Dvali, N.; Bolokadze, N.; Sharvadze, L.; Gabunia, P.; Karchava, M.; Tchelidze, T.; Tsertsvadze, T.; DeHovitz, J.; Del Rio, C.; et al. Epidemiology of Human Immunodeficiency Virus (HIV) Drug Resistance in HIV Patients with Virologic Failure of First-Line Therapy in the Country of Georgia. J. Med. Virol. 2019, 91, 235–240. [Google Scholar] [CrossRef]
- Armenia, D.; Bouba, Y.; Gagliardini, R.; Gori, C.; Bertoli, A.; Borghi, V.; Gennari, W.; Micheli, V.; Callegaro, A.P.; Gazzola, L.; et al. Evaluation of Virological Response and Resistance Profile in HIV-1 Infected Patients Starting a First-Line Integrase Inhibitor-Based Regimen in Clinical Settings. J. Clin. Virol. 2020, 130, 104534. [Google Scholar] [CrossRef]
- El-Khatib, Z.; Ekstrom, A.M.; Ledwaba, J.; Mohapi, L.; Laher, F.; Karstaedt, A.; Charalambous, S.; Petzold, M.; Katzenstein, D.; Morris, L. Viremia and Drug Resistance among HIV-1 Patients on Antiretroviral Treatment: A Cross-Sectional Study in Soweto, South Africa. AIDS 2010, 24, 1679–1687. [Google Scholar] [CrossRef]
- Hoffmann, C.J.; Ledwaba, J.; Li, J.-F.; Johnston, V.; Hunt, G.; Fielding, K.L.; Chaisson, R.E.; Churchyard, G.J.; Grant, A.D.; Johnson, J.A.; et al. Resistance to Tenofovir-Based Regimens during Treatment Failure of Subtype C HIV-1 in South Africa. Antivir. Ther. 2013, 18, 915–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mziray, S.R.; Kumburu, H.H.; Assey, H.B.; Sonda, T.B.; Mahande, M.J.; Msuya, S.E.; Kiwelu, I.E. Patterns of Acquired HIV-1 Drug Resistance Mutations and Predictors of Virological Failure in Moshi, Northern Tanzania. PLoS ONE 2020, 15, e0232649. [Google Scholar] [CrossRef] [PubMed]
- Neogi, U.; Engelbrecht, S.; Claassen, M.; Jacobs, G.B.; van Zyl, G.; Preiser, W.; Sonnerborg, A. Mutational Heterogeneity in P6 Gag Late Assembly (L) Domains in HIV-1 Subtype C Viruses from South Africa. AIDS Res. Hum. Retrovir. 2016, 32, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Riddler, S.A.; Haubrich, R.; DiRienzo, A.G.; Peeples, L.; Powderly, W.G.; Klingman, K.L.; Garren, K.W.; George, T.; Rooney, J.F.; Brizz, B.; et al. Class-Sparing Regimens for Initial Treatment of HIV-1 Infection. N. Engl. J. Med. 2008, 358, 2095–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen-Zvi, M.; Altmann, A.; Prosperi, M.; Aharoni, E.; Neuvirth, H.; Sönnerborg, A.; Schülter, E.; Struck, D.; Peres, Y.; Incardona, F.; et al. Selecting Anti-HIV Therapies Based on a Variety of Genomic and Clinical Factors. Bioinformatics 2008, 24, i399–i406. [Google Scholar] [CrossRef]
- Svärd, J.; Mugusi, S.; Mloka, D.; Neogi, U.; Meini, G.; Mugusi, F.; Incardona, F.; Zazzi, M.; Sönnerborg, A. Drug Resistance Testing through Remote Genotyping and Predicted Treatment Options in Human Immunodeficiency Virus Type 1 Infected Tanzanian Subjects Failing First or Second Line Antiretroviral Therapy. PLoS ONE 2017, 12, e0178942. [Google Scholar] [CrossRef] [Green Version]
- Vergani, B.; Cicero, M.L.; Vigano’, O.; Sirianni, F.; Ferramosca, S.; Vitiello, P.; Di Vincenzo, P.; Pia De Pasquale, M.; Galli, M.; Rusconi, S. Evolution of the HIV-1 Protease Region in Heavily Pretreated HIV-1 Infected Patients Receiving Atazanavir. J. Clin. Virol. 2008, 41, 154–159. [Google Scholar] [CrossRef]
- Abecasis, A.B.; Deforche, K.; Snoeck, J.; Bacheler, L.T.; McKenna, P.; Carvalho, A.P.; Gomes, P.; Camacho, R.J.; Vandamme, A.-M. Protease Mutation M89I/V Is Linked to Therapy Failure in Patients Infected with the HIV-1 Non-B Subtypes C, F or G. AIDS 2005, 19, 1799–1806. [Google Scholar] [CrossRef]
- Gong, Y.F.; Robinson, B.S.; Rose, R.E.; Deminie, C.; Spicer, T.P.; Stock, D.; Colonno, R.J.; Lin, P.F. In Vitro Resistance Profile of the Human Immunodeficiency Virus Type 1 Protease Inhibitor BMS-232632. Antimicrob. Agents Chemother. 2000, 44, 2319–2326. [Google Scholar] [CrossRef] [Green Version]
- Malan, D.R.N.; Krantz, E.; David, N.; Yang, R.; Mathew, M.; Iloeje, U.H.; Su, J.; McGrath, D.; 089 Stuy Group. 96-Week Efficacy and Safety of Atazanavir, with and without Ritonavir, in a HAART Regimen in Treatment-Naive Patients. J. Int. Assoc. Physicians AIDS Care 2010, 9, 34–42. [Google Scholar] [CrossRef]
- Molina, J.-M.; Andrade-Villanueva, J.; Echevarria, J.; Chetchotisakd, P.; Corral, J.; David, N.; Moyle, G.; Mancini, M.; Percival, L.; Yang, R.; et al. Once-Daily Atazanavir/Ritonavir versus Twice-Daily Lopinavir/Ritonavir, Each in Combination with Tenofovir and Emtricitabine, for Management of Antiretroviral-Naive HIV-1-Infected Patients: 48 Week Efficacy and Safety Results of the CASTLE Study. Lancet 2008, 372, 646–655. [Google Scholar] [CrossRef]
- Rosenbloom, D.I.S.; Hill, A.L.; Rabi, S.A.; Siliciano, R.F.; Nowak, M.A. Antiretroviral Dynamics Determines HIV Evolution and Predicts Therapy Outcome. Nat. Med. 2012, 18, 1378–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockdale, A.J.; Saunders, M.J.; Boyd, M.A.; Bonnett, L.J.; Johnston, V.; Wandeler, G.; Schoffelen, A.F.; Ciaffi, L.; Stafford, K.; Collier, A.C.; et al. Effectiveness of Protease Inhibitor/Nucleos(t)Ide Reverse Transcriptase Inhibitor-Based Second-Line Antiretroviral Therapy for the Treatment of Human Immunodeficiency Virus Type 1 Infection in Sub-Saharan Africa: A Systematic Review and Meta-Analysis. Clin. Infect. Dis. 2018, 66, 1846–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colonno, R.J.; Thiry, A.; Limoli, K.; Parkin, N. Activities of Atazanavir (BMS-232632) against a Large Panel of Human Immunodeficiency Virus Type 1 Clinical Isolates Resistant to One or More Approved Protease Inhibitors. Antimicrob. Agents Chemother. 2003, 47, 1324–1333. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.-Y.; Taylor, J.; Fessel, W.J.; Kaufman, D.; Towner, W.; Troia, P.; Ruane, P.; Hellinger, J.; Shirvani, V.; Zolopa, A.; et al. HIV-1 Protease Mutations and Protease Inhibitor Cross-Resistance. Antimicrob. Agents Chemother. 2010, 54, 4253–4261. [Google Scholar] [CrossRef] [Green Version]
- Yanchunas, J.; Langley, D.R.; Tao, L.; Rose, R.E.; Friborg, J.; Colonno, R.J.; Doyle, M.L. Molecular Basis for Increased Susceptibility of Isolates with Atazanavir Resistance-Conferring Substitution I50L to Other Protease Inhibitors. Antimicrob. Agents Chemother. 2005, 49, 3825–3832. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.-Y.; Shafer, R.W. Geographically-Stratified HIV-1 Group M Pol Subtype and Circulating Recombinant Form Sequences. Sci. Data 2018, 5, 180148. [Google Scholar] [CrossRef] [Green Version]
- Stanford HIV Drug Resistance Database HIV Drug Resistance Database. Available online: https://hivdb.stanford.edu/pages/documentPage/PI_mutationClassification.html (accessed on 2 March 2022).
- Rhee, S.-Y.; Sankaran, K.; Varghese, V.; Winters, M.A.; Hurt, C.B.; Eron, J.J.; Parkin, N.; Holmes, S.P.; Holodniy, M.; Shafer, R.W. HIV-1 Protease, Reverse Transcriptase, and Integrase Variation. J. Virol. 2016, 90, 6058–6070. [Google Scholar] [CrossRef] [Green Version]
- Holm, S. A Simple Sequentially Rejective Multiple Test Procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Rhee, S.-Y.; Liu, T.F.; Holmes, S.P.; Shafer, R.W. HIV-1 Subtype B Protease and Reverse Transcriptase Amino Acid Covariation. PLoS Comput. Biol. 2007, 3, e87. [Google Scholar] [CrossRef] [Green Version]
- Scutari, M. Learning Bayesian Networks with the Bnlearn R Package. J. Stat. Soft. 2010, 35, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Almende, B.V.; Benoit, T.; Titouan, R. VisNetwork: Network Visualization Using “vis.Js” Library. Available online: https://CRAN.R-project.org/package=visNetwork (accessed on 2 March 2022).



{kind=link}
{kind=link}
| AuthorYr | Study Type | # Total ATV | # bATV | # ATV | % DRMs 1 | Median Year | Country | Subtypes (%) 2 |
|---|---|---|---|---|---|---|---|---|
| Large clinical trials and cohorts for which genotypic resistance testing was routinely available at virological failure | ||||||||
| EuResist Network [14] | Cohort | 562 | 286 | 276 | 10.3 | 2012 | Europe | B (57.8), G (16.2), 02_AG (12) |
| Stanford University Hospital | Cohort | 152 | 142 | 10 | 9.2 | 2010 | U.S. | B (96.7) |
| Mollan12 [16] | ACTG A5202 | 137 | 137 | 0 | 5.8 | 2006 | U.S. | B (97.1) |
| Kantor15 [17] | ACTG A5175 | 117 | 19 | 98 | 14.5 | 2006 | Multi-continents | C (55.6), B (41.9) |
| Lennox14 [18] | ACTG A5257 | 69 | 69 | 0 | 2.9 | 2010 | U.S. | B (97.1) |
| Case series and cohorts for which genotypic resistance testing may not have been routinely available at virological failure | ||||||||
| Soldi19 [10] | Cohort | 149 | 81 | 68 | 30.2 | 2015 | Brazil | B (75.8), F (12.8) |
| Tarasova21 [15] | Cohort | 43 | 16 | 27 | 37.2 | 2017 | Russia | A (90.7) |
| Kouamou19 [19] | Cohort | 40 | 40 | 0 | 12.5 | 2017 | Zimbabwe | C (100) |
| de Carvalho Lima20 [20] | Cohort | 37 | 28 | 9 | 54.1 | 2010 | Brazil | B (81.1), F (16.2) |
| Acharya14 [21] | Cohort | 35 | 35 | 0 | 48.6 | 2013 | India | C (80), A (20) |
| Ndashimye18 [22] | Cohort | 33 | 33 | 0 | 42.4 | 2016 | Uganda | A (57.6), D (24.2), B (15.2) |
| Gulick04 [23] | ACTG A5095 | 24 | 1 | 23 | 8.3 | 2003 | U.S. | B (100) |
| Colonno04 [24] | Case series from clinical trials 3 | 21 | 0 | 21 | 100 | 2000 | Multi-continents | B (71.4), C (28.6) |
| Chimukangara16 [25] | Cohort | 17 | 17 | 0 | 29.4 | 2015 | Zimbabwe | C (100) |
| Posada Cespedes21 [12] | Cohort | 13 | 7 | 6 | 7.7 | 2015 | South Africa | C (100) |
| Makwaga20 [26] | Cohort | 11 | 11 | 0 | 36.4 | 2020 | Kenya | A (63.6), B (18.2), D (18.2) |
| de Sa Filho08 [27] | Cohort | 10 | 8 | 2 | 80 | 2006 | Brazil | B (80), F (20) |
| Kolomeets14 [28] | Cohort | 10 | 0 | 10 | 30 | 2012 | Russia | A (70), 02_AG (30) |
| Alves19 [29] | Cohort | 3 | 2 | 1 | 0 | 2017 | Brazil | C (66.7), B (33.3) |
| Kim13 [30] | Cohort | 3 | 1 | 2 | 33.3 | 2011 | Korea | B (100) |
| Karkashadze19 [31] | Cohort | 2 | 0 | 2 | 100 | 2015 | Republic Of Georgia | A (50), B (50) |
| Armenia20 [32] | Cohort | 1 | 1 | 0 | 0 | 2012 | Italy | B (100) |
| El-Khatib10 [33] | Cohort | 1 | 1 | 0 | 100 | 2008 | South Africa | C (100) |
| Hoffmann13 [34] | Cohort | 1 | 0 | 1 | 0 | 2010 | South Africa | C (100) |
| Mziray20 [35] | Cohort | 1 | 1 | 0 | 0 | 2018 | Tanzania | C (100) |
| Neogi16 [36] | Cohort | 1 | 0 | 1 | 0 | 2013 | South Africa | C (100) |
| Riddler08 [37] | ACTG A5142 | 1 | 1 | 0 | 0 | 2004 | U.S. | D (100) |
| Rosen-Zvi08 [38] | Cohort | 1 | 1 | 0 | 0 | 2006 | Germany | B (100) |
| Svard17 [39] | Cohort | 1 | 1 | 0 | 0 | 2013 | Tanzania | A (100) |
| Vergani08 [40] | Cohort | 1 | 0 | 1 | 0 | 2006 | Italy | B (100) |
| # Patients, (% of Total; n = 1437) | # Patients with ≥1 DRMs 1, (% of Row Total) | Median # DRMs in Patients with ≥1 DRM (IQR) | |
|---|---|---|---|
| Unboosted vs. boosted | |||
| Unboosted | 558 (37.3) | 117 (21.0) | 3.0 (1.0–4.0) |
| Boosted | 939 (62.7) | 147 (15.7) | 2.0 (1.0–4.0) |
| Subtype B vs. non-subtype B | |||
| Subtype B | 570 (38.1) | 150 (16.2) | 3.0 (1.0–4.0) |
| Non-subtype B | 927 (61.9) | 114 (20.0) | 3.0 (1.0–4.0) |
| ART-naïve vs. ART-experienced | |||
| ART-naïve | 907 (60.6) | 136 (15.0) | 3.0 (1.0–4.0) |
| ART-experienced | 590 (39.4) | 128 (21.7) | 2.0 (1.0–4.0) |
| Year of ART initiation2 | |||
| 1993–2004 | 134 (11.9) | 24 (17.9) | 2.0 (1.0–2.1) |
| 2005–2006 | 362 (32.1) | 44 (12.1) | 1.0 (1.0–2.4) |
| 2007–2009 | 316 (28.0) | 26 (8.2) | 2.0 (1.0–2.8) |
| 2010–2018 | 315 (28.0) | 29 (9.2) | 2.0 (1.0–2.3) |
| DRM 1 | Classification 2 | % in the 264 Patients with a PI-Associated DRM | Median # Co-Occurring DRMs (IQR) |
|---|---|---|---|
| I50L | Major | 34.1 | 2 (0.2–3) |
| M46I | Major | 32.6 | 3 (2–5) |
| V82A | Major | 22.3 | 4 (3–5) |
| L90M | Major | 19.3 | 3 (2–4.5) |
| I54V | Major | 16.3 | 4 (3–5) |
| N88S | Major | 10.2 | 3 (2–4) |
| M46L | Major | 7.6 | 3 (2–4) |
| V32I | Major | 6.4 | 3 (2–5) |
| I84V | Major | 6.1 | 3 (2–5) |
| I54L | Major | 4.2 | 3 (3–4.5) |
| G48V | Major | 3.4 | 3 (2–3) |
| I47V | Major | 2.7 | 5 (4.5–7) |
| I50V | Major | 2.3 | 4 (3–5) |
| L76V | Major | 2.3 | 5.5 (4.2–6) |
| I47A | Major | 1.5 | 3.5 (2–5) |
| V82M | Major | 1.5 | 2 (1.7–3) |
| V82T | Major | 1.5 | 4.5 (3.5–5.5) |
| D30N | Major | 1.1 | 4 (3.5–7) |
| G48A | Major | 1.1 | 6 (4.5–6.5) |
| V82F | Major | 1.1 | 6 (5–6.5) |
| V82L | Major | 1.1 | 3 (1.5–4.5) |
| I54A | Major | 0.8 | 3.5 (3.2–3.7) |
| V82S | Major | 0.8 | 3 (3–3) |
| G48M | Major | 0.4 | 2 (2–2) |
| I54M | Major | 0.4 | 5 (5–5) |
| I54S | Major | 0.4 | 2 (2–2) |
| I54T | Major | 0.4 | 2 (2–2) |
| V82C | Major | 0.4 | 4 (4–4) |
| N88T | Major | 0.4 | 3 (3–3) |
| L33F | Accessory | 20.8 | 4 (2–5) |
| Q58E | Accessory | 15.9 | 3 (1–5) |
| K20T | Accessory | 14.4 | 2 (1–4) |
| G73S | Accessory | 11.7 | 3 (1–4) |
| L10F | Accessory | 9.8 | 4 (2–5) |
| F53L | Accessory | 9.8 | 3.5 (2–5) |
| K43T | Accessory | 8.7 | 4 (2–5) |
| L24I | Accessory | 6.1 | 4 (2–4.2) |
| L23I | Accessory | 4.2 | 3 (1.5–4.5) |
| T74P | Accessory | 3 | 3 (2–4) |
| G73T | Accessory | 1.5 | 3.5 (2.7–4.5) |
| L89V | Accessory | 1.5 | 3.5 (2–5.2) |
| N83D | Accessory | 1.1 | 3 (3–4) |
| N88D | Accessory | 1.1 | 3 (2.5–6.5) |
| G73C | Accessory | 0.8 | 3.5 (2.2–4.7) |
| L24F | Accessory | 0.4 | 0 (0–0) |
| M46V | Accessory | 0.4 | 2 (2–2) |
| G73A | Accessory | 0.4 | 6 (6–6) |
| G73V | Accessory | 0.4 | 8 (8–8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhee, S.-Y.; Boehm, M.; Tarasova, O.; Di Teodoro, G.; Abecasis, A.B.; Sönnerborg, A.; Bailey, A.J.; Kireev, D.; Zazzi, M.; the EuResist Network Study Group; et al. Spectrum of Atazanavir-Selected Protease Inhibitor-Resistance Mutations. Pathogens 2022, 11, 546. https://doi.org/10.3390/pathogens11050546
Rhee S-Y, Boehm M, Tarasova O, Di Teodoro G, Abecasis AB, Sönnerborg A, Bailey AJ, Kireev D, Zazzi M, the EuResist Network Study Group, et al. Spectrum of Atazanavir-Selected Protease Inhibitor-Resistance Mutations. Pathogens. 2022; 11(5):546. https://doi.org/10.3390/pathogens11050546
Chicago/Turabian StyleRhee, Soo-Yon, Michael Boehm, Olga Tarasova, Giulia Di Teodoro, Ana B. Abecasis, Anders Sönnerborg, Alexander J. Bailey, Dmitry Kireev, Maurizio Zazzi, the EuResist Network Study Group, and et al. 2022. "Spectrum of Atazanavir-Selected Protease Inhibitor-Resistance Mutations" Pathogens 11, no. 5: 546. https://doi.org/10.3390/pathogens11050546
APA StyleRhee, S. -Y., Boehm, M., Tarasova, O., Di Teodoro, G., Abecasis, A. B., Sönnerborg, A., Bailey, A. J., Kireev, D., Zazzi, M., the EuResist Network Study Group, & Shafer, R. W. (2022). Spectrum of Atazanavir-Selected Protease Inhibitor-Resistance Mutations. Pathogens, 11(5), 546. https://doi.org/10.3390/pathogens11050546