
The GP-45 Protein, a Highly Variable Antigen from Babesia bigemina, Contains Conserved B-Cell Epitopes in Geographically Distant Isolates


 , , ,
, , , 


Abstract
:1. Introduction
2. Results
2.1. Accession Number
2.2. In Silico Analysis of GP-45 Protein Sequences
2.3. Selection of Predicted Peptides
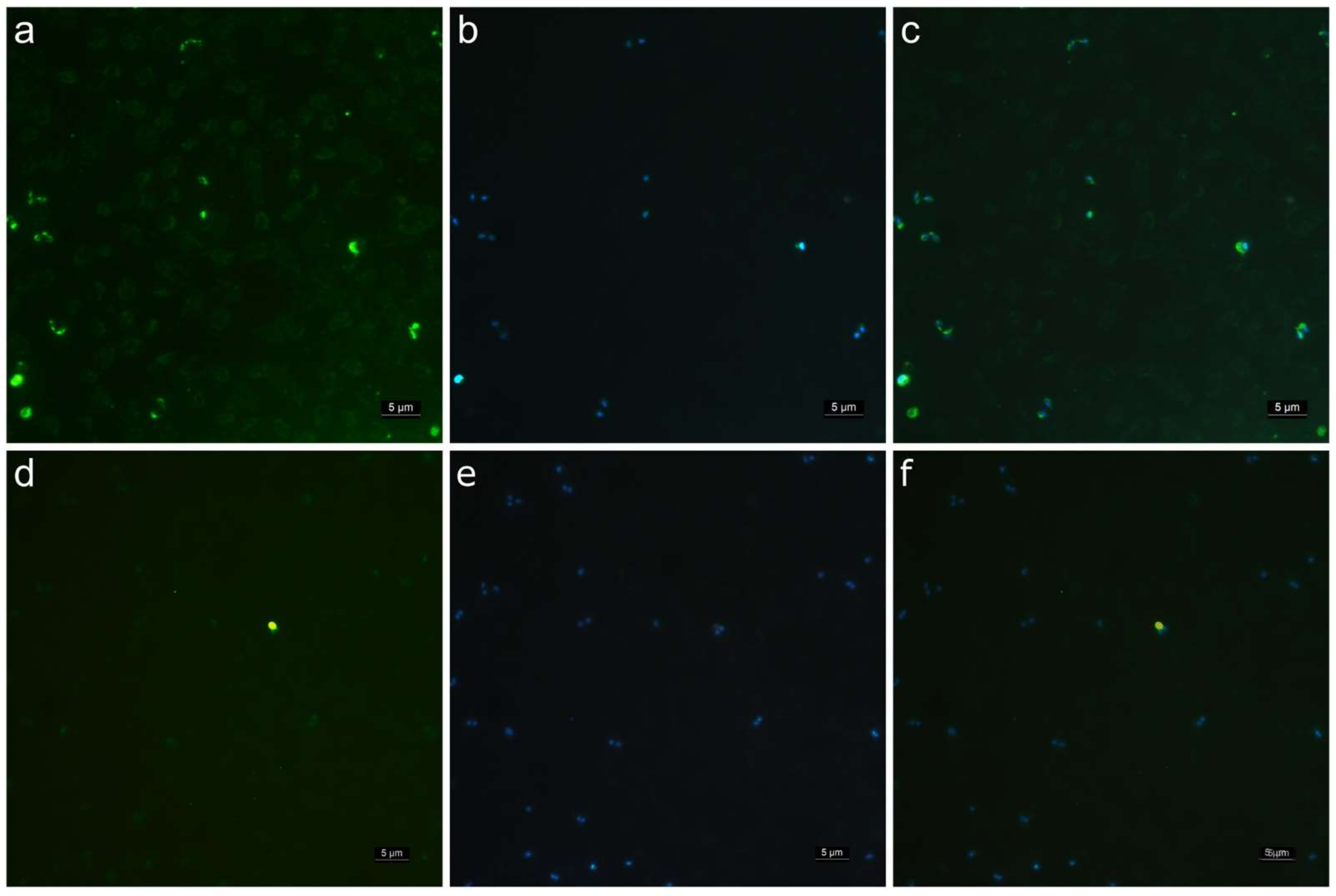
2.4. Evaluation of the Specificity of Anti-GP-45 Polyclonal Antibodies by Immunofluorescence
2.5. Evaluation of the Specificity of Anti-GP-45 Polyclonal Antibodies by Western Blot
2.6. Evaluation of the Presence of Anti-Babesia Antibodies in Naturally Infected Cattle
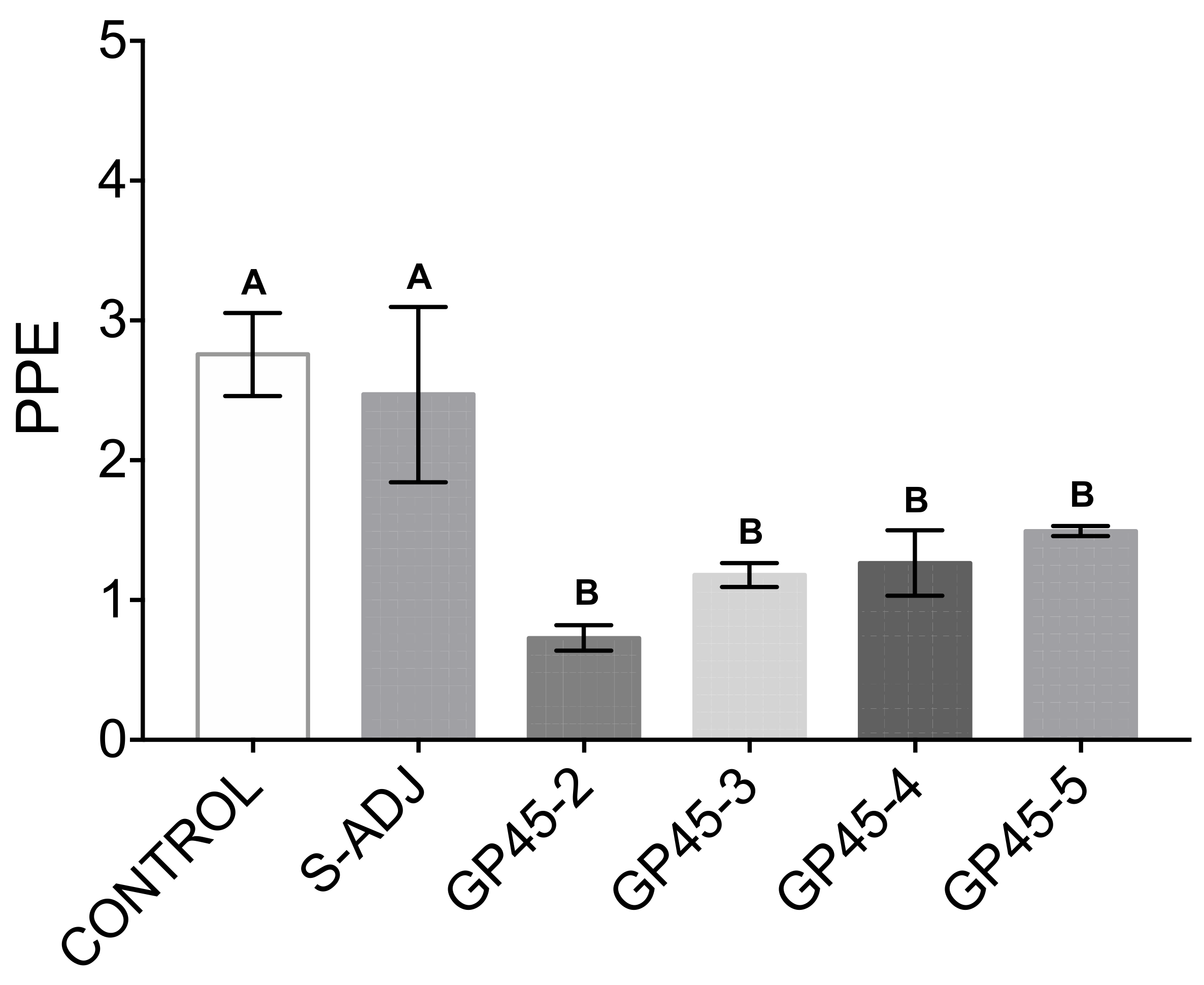
2.7. Evaluation of the Neutralizing Ability of Anti-Babesia Antibodies In Vitro Culture of Erythrocytes Infected with B. bigemina
3. Discussion
4. Materials and Methods
4.1. Evaluation of the Presence of Anti-Babesia Antibodies in Naturally Infected Cattle
4.2. In Silico Analysis of Predicted GP-45 Protein Sequences
4.3. Generation of Hyperimmune Sera against GP-45 Peptides in Rabbits and Mice
4.4. Evaluation of the Specificity of the Antibodies Present in Mouse Polyclonal Sera
4.5. Specificity of Anti-GP-45 Mouse Sera Used for Expression Analysis
4.6. Recognition of GP-45 Peptides by Antibodies Present in Cattle Naturally Infected with B. bigemina
4.7. In Vitro Neutralization Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dos Santos, J.P.C.; Cardoso, L.C.; Monteiro, E.; Lemes, N. A Fractional-Order Epidemic Model for Bovine Babesiosis Disease and Tick Populations. Abstr. Appl. Anal. 2015, 2015, 729894. [Google Scholar] [CrossRef]
- Domingos, A.; Antunes, S.; Villar, M.; De La Fuente, J. Functional Genomics of Tick Vectors Challenged with the Cattle Parasite Babesia bigemina. Methods Mol. Biol. 2014, 1247, 475–489. [Google Scholar] [CrossRef] [Green Version]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Ueti, M.W.; Olafson, P.U.; Freeman, J.M.; Johnson, W.C.; Scoles, G.A. A Virulent Babesia bovis Strain Failed to Infect White-Tailed Deer (Odocoileus virginianus). PLoS ONE 2015, 10, e0131018. [Google Scholar] [CrossRef] [Green Version]
- Bock, R.; Jackson, L.; De Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129, S247–S269. [Google Scholar] [CrossRef]
- Ghosh, S.; Azhahianambi, P.; Yadav, M.P. Upcoming and future strategies of tick control: A review. J. Vector Borne Dis. 2007, 44, 79–89. [Google Scholar]
- Suarez, C.E.; Palmer, G.H.; Hotzel, I.; McElwain, T.F. Structure, sequence, and transcriptional analysis of the Babesia bovis rap-1 multigene locus. Mol. Biochem. Parasitol. 1998, 93, 215–224. [Google Scholar] [CrossRef]
- Fisher, T.G.; McElwain, T.F.; Palmer, G.H. Molecular Basis for Variable Expression of Merozoite Surface Antigen gp45 among American Isolates of Babesia bigemina. Infect. Immun. 2001, 69, 3782–3790. [Google Scholar] [CrossRef] [Green Version]
- McElwain, T.F.; Perryman, L.E.; Musoke, A.J.; McGuire, T.C. Molecular characterization and immunogenicity of neutralization-sensitive Babesia bigemina merozoite surface proteins. Mol. Biochem. Parasitol. 1991, 47, 213–222. [Google Scholar] [CrossRef]
- Mtshali, P.S.; Tsotetsi, A.M.; Thekisoe, M.M.O.; Mtshali, M.S. Nested PCR Detection and Phylogenetic Analysis of Babesia bovis and Babesia bigemina in Cattle from Peri-Urban Localities in Gauteng Province, South Africa. J. Vet. Med Sci. 2014, 76, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Mosqueda, J.; Mejía-López, A.S.; Mercado-Uriostegui, M.A. Molecular Mechanisms of Babesia Invasion: Potential Targets for Vaccine Development. In Anti-infective Research and Development: Updates on Infection Mechanisms and Treatments; Guerrero Manriquez, G.G., Ed.; Bentham Science: Sharjah, United Arab Emirates, 2020; pp. 161–181. [Google Scholar]
- Mosqueda, J.; Figueroa, J.V.; Alvarez, A.; Bautista, R.; Falcon, A.; Ramos, A.; Canto, G.; Vega, C.A. Advances in the development of molecular tools for the control of bovine babesiosis in Mexico. Parassitologia 2007, 49, 19–22. [Google Scholar] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Springer: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Mosqueda, J.; Hidalgo-Ruiz, M.; Calvo-Olvera, D.A.; Hernandez-Silva, D.J.; Ueti, M.W.; Mercado-Uriostegui, M.A.; Rodriguez, A.; Ramos-Aragon, J.A.; Hernandez-Ortiz, R.; Kawazu, S.-I.; et al. RON2, a novel gene in Babesia bigemina, contains conserved, immunodominant B-cell epitopes that induce antibodies that block merozoite invasion. Parasitology 2019, 146, 1646–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carcy, B.; Bonnefoy, S.; Guillotte, M.; Le Scanf, C.; Grellier, P.; Schrevel, J.; Fandeur, T.; Mercereau-Puijalon, O. A large multi gene family expressed during the erythrocytic schizogony of Plasmodium falciparum. Mol. Biochem. Parasitol. 1994, 68, 221–233. [Google Scholar] [CrossRef]
- Carcy, B.; Precigout, E.; Valentin, A.; Gorenflot, A.; Schrevel, J. A 37-kilodalton glycoprotein of Babesia divergens is a major component of a protective fraction containing low-molecular-mass culture-derived exoantigens. Infect. Immun. 1995, 63, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachebat, J.; Kadekoppala, M.; Grainger, M.; Dluzewski, A.R.; Gunaratne, R.S.; Scott-Finnigan, T.J.; Ogun, S.A.; Ling, I.T.; Bannister, L.H.; Taylor, H.M.; et al. Extensive proteolytic processing of the malaria parasite merozoite surface protein 7 during biosynthesis and parasite release from erythrocytes. Mol. Biochem. Parasitol. 2007, 151, 59–69. [Google Scholar] [CrossRef] [PubMed]
- McElwain, T.F.; Perryman, L.E.; Davis, W.C.; McGuire, T.C. Antibodies define multiple proteins with epitopes exposed on the surface of live Babesia bigemina merozoites. J. Immunol. 1987, 138, 2298–2304. [Google Scholar]
- Al-Adhami, B.H.; Gajadhar, A.A. A new multi-host species indirect ELISA using protein A/G conjugate for detection of anti-Toxoplasma gondii IgG antibodies with comparison to ELISA-IgG, agglutination assay and Western blot. Veter- Parasitol. 2014, 200, 66–73. [Google Scholar] [CrossRef]
- Chung, C.J.; Suarez, C.E.; Bandaranayaka-Mudiyanselage, C.L.; Bandaranayaka-Mudiyanselage, C.-B.; Rzepka, J.; Heiniger, T.J.; Chung, G.; Lee, S.S.; Adams, E.; Yun, G.; et al. A novel modified-indirect ELISA based on spherical body protein 4 for detecting antibody during acute and long-term infections with diverse Babesia bovis strains. Parasites Vectors 2017, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Pérez, I.M.; Lira-Amaya, J.J.; Castañeda-Arriola, R.O.; Cantú-Covarrubias, A.; Mejía-Estrada, F.; Polanco-Martínez, D.J.; Figueroa-Millán, J.V. Comparación de pruebas serológicas para el diagnóstico epidemiológico de babesiosis bovina transmitida por garrapatas. Entomol. Mex. 2017, 4, 611–616. [Google Scholar]
- Kim, C.; Alhassan, A.; Verdida, R.A.; Yokoyama, N.; Xuan, X.; Fujisaki, K.; Kawazu, S.-I.; Igarashi, I. Development of two immunochromatographic tests for the serodiagnosis of bovine babesiosis. Vet. Parasitol. 2007, 148, 137–143. [Google Scholar] [CrossRef]
- International Office of Epizootics (OIE). Bovine babesiosis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; 2021. Available online: https://www.oie.int/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 15 October 2021).
- Hines, S.A.; Palmer, G.H.; Jasmer, D.P.; McGuire, T.C.; McElwain, T.F. Neutralization-sensitive merozoite surface antigens of Babesia bovis encoded by members of a polymorphic gene family. Mol. Biochem. Parasitol. 1992, 55, 85–94. [Google Scholar] [CrossRef]
- Dominguez, M.; Echaide, I.; de Echaide, S.T.; Mosqueda, J.; Cetrá, B.; Suarez, C.; Florin-Christensen, M. In silico predicted conserved B-cell epitopes in the merozoite surface antigen-2 family of B. bovis are neutralization sensitive. Vet. Parasitol. 2010, 167, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda, J.; Olvera-Ramirez, A.; Aguilar-Tipacamu, G.; Canto, G.J. Current Advances in Detection and Treatment of Babesiosis. Curr. Med. Chem. 2012, 19, 1504–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, D.L. Database resources of the National Center for Biotechnology. Nucleic Acids Res. 2003, 31, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K. TMbase-A database of membrane spanning proteins segments. Biol. Chem. 1993, 374, 166. [Google Scholar]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Sigrist, C.J.A.; de Castro, E.; Cerutti, L.; Cuche, B.A.; Hulo, N.; Bridge, A.; Bougueleret, L.; Xenarios, I. New and continuing developments at PROSITE. Nucleic Acids Res. 2012, 41, D344–D347. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2013, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Copley, R.R.; Doerks, T.; Ponting, C.P.; Bork, P. SMART: A web-based tool for the study of genetically mobile domains. Nucleic Acids Res. 2000, 28, 231–234. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P.S. BcePred: Prediction of Continuous B-Cell Epitopes in Antigenic Sequences Using Physico-chemical Properties. In Artificial Immune Systems, ICARIS; Nicosia, G., Cutello, V., Bentley, P.J., Timmis, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 3239. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.M.R.; Guo, D.; Hodges, R.S. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: Correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites. Biochemistry 1986, 25, 5425–5432. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Raghava, G.P.S. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins Struct. Funct. Bioinform. 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Larsen, J.E.P.; Lund, O.; Nielsen, M. Improved method for predicting linear B-cell epitopes. Immunome Res. 2006, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hernández-Silva, D.J.; Valdez-Espinoza, U.M.; Mercado-Uriostegui, M.A.; Aguilar-Tipacamú, G.; Ramos-Aragón, J.A.; Hernández-Ortiz, R.; Ueti, M.; Mosqueda, J. Immunomolecular Characterization of MIC-1, a Novel Antigen in Babesia bigemina, Which Contains Conserved and Immunodominant B-Cell Epitopes that Induce Neutralizing Antibodies. Vet. Sci. 2018, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, C.A.; Buening, G.M.; Rodriguez, S.D.; Carson, C.A.; McLaughlin, K. Cryopreservation of Babesia bigemina for in vitro cultivation. Am. J. Vet. Res. 1985, 46. [Google Scholar]
- Rodríguez-Hernández, E.; Mosqueda, J.; Alvarez-Sánchez, M.E.; Neri, A.F.; Mendoza-Hernández, G.; Camacho-Nuez, M. The identification of a VDAC-like protein involved in the interaction of Babesia bigemina sexual stages with Rhipicephalus microplus midgut cells. Vet. Parasitol. 2012, 187, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Ruiz, M.; Suarez, C.E.; Mercado-Uriostegui, M.A.; Hernandez-Ortiz, R.; Ramos, J.A.; Galindo-Velasco, E.; León-Ávila, G.; Hernández, J.M.; Mosqueda, J. Babesia bovis RON2 contains conserved B-cell epitopes that induce an invasion-blocking humoral immune response in immunized cattle. Parasites Vectors 2018, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- González, C.G.; Castán, J.P. Capítulo 8: Pruebas diagnósticas. Concordancia. Semer.-Med. De Fam. 2007, 33, 509–519. [Google Scholar] [CrossRef]
- Hötzel, I.; Suarez, C.E.; McElwain, T.F.; Palmer, G.H. Genetic variation in the dimorphic regions of rap-1 genes and rap-1 loci of Babesia bigemina. Mol. Biochem. Parasitol. 1997, 90, 479–489. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Isolate/Strain | Seed | ||
|---|---|---|---|
| GenBank ID | Coverage | Identity | |
| Veracruz | AEJ89907 | 100% | 99.70% |
| Jalisco | AEJ89910 | 100% | 99.40% |
| Mexico | AEJ89911 | 100% | 99.40% |
| Guerrero | AEJ89909 | 100% | 99.10% |
| Nayarit | AEJ89906 | 100% | 99.10% |
| Mexico JG-29 | AAG28757 | 100% | 98.30% |
| Argentina Corrientes | AEJ89908 | 100% | 98.00% |
| Chiapas | OM488277 | 100% | 94.90% |
| Australia Bond | XP_012767625.1 | 88.60% | 95.50% |
| South Africa Khutsong A14 | AGU67945 | 78.30% | 98.50% |
| South Africa Devon A04 | AGU67946 | 78.30% | 98.50% |
| South Africa Eikenhof A13 | AGU67947 | 78.30% | 97.10% |
| South Africa Sharpe A17 | AGU67944 | 77.50% | 58.90% |
| Prediction Order | Position (aa) | Length (aa) | Amino Acid Sequence |
|---|---|---|---|
| 1 | 282–301 | 19 | ERAVSGATTHGGDARGVNP |
| 2 | 52–70 | 18 | MHIVSNLLDVEPIVGMYG |
| 3 | 52–70 | 18 | MHIVSKLLDVQPIVGMYG |
| 4 | 142–159 | 17 | GFLSTATDVPESDLAKK |
| 5 | 178–195 | 17 | NLQMFLKVFYNKNSPLF |
| n = 126 | Estate | Ranch/Farm | Total | Peptide 1 | Peptide 2 | Peptide 3 | Peptide 4 | Peptide 5 | Total | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| “+” | “−” | “+” | “−” | “+” | “−” | “+” | “−” | “+” | “−” | |||||
| IFAT Positive n = 81 | Aguascalientes | Villa Guadalupe | 35 | 7 | 0 | 7 | 0 | 5 | 2 | 6 | 1 | 7 | 0 | 7 |
| Las Palomas | 23 | 1 | 20 | 4 | 17 | 7 | 14 | 10 | 19 | 5 | 24 | |||
| Granja María I | 4 | 0 | 3 | 1 | 2 | 2 | 3 | 1 | 3 | 1 | 4 | |||
| Querétaro | Granja Areceli | 2 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | |
| Sinaloa | El Torito | 20 | 6 | 2 | 7 | 1 | 6 | 2 | 7 | 1 | 7 | 1 | 8 | |
| La Herradura | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | |||
| El Barón | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | |||
| El Moral | 8 | 0 | 8 | 0 | 8 | 0 | 7 | 1 | 8 | 0 | 8 | |||
| Veracruz | El Arbolito | 24 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | |
| Playa Vicente | 2 | 0 | 2 | 0 | 1 | 1 | 2 | 0 | 2 | 0 | 2 | |||
| La Esperanza | 2 | 1 | 2 | 1 | 3 | 0 | 2 | 1 | 2 | 1 | 3 | |||
| Las Torres | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | |||
| San Faudila | 3 | 0 | 2 | 1 | 3 | 0 | 3 | 0 | 3 | 0 | 3 | |||
| El orijuelo | 11 | 0 | 9 | 2 | 10 | 1 | 11 | 0 | 11 | 0 | 11 | |||
| Buenos Aires | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | 0 | 2 | |||
| Total Positive/Negative | 81 | 77 | 4 | 71 | 10 | 66 | 15 | 66 | 15 | 73 | 8 | 81 | ||
| IFAT Negative n = 45 | Querétaro | Amazcala | 16 | 0 | 6 | 0 | 6 | 0 | 6 | 0 | 6 | 0 | 6 | 6 |
| Los Moreno | 0 | 10 | 0 | 10 | 0 | 10 | 0 | 10 | 0 | 10 | 10 | |||
| Durango | Arroyo Seco | 29 | 0 | 29 | 0 | 29 | 3 | 26 | 4 | 25 | 15 | 14 | 29 | |
| Total Positive/Negative | 45 | 0 | 45 | 0 | 45 | 3 | 42 | 4 | 41 | 15 | 30 | 45 | ||
| Parameter | Formula | Peptide 1 | Peptide 2 | Peptide 3 | Peptide 4 | Peptide 5 |
|---|---|---|---|---|---|---|
| TP | a | 77 | 71 | 66 | 66 | 73 |
| FP | b | 0 | 0 | 3 | 4 | 15 |
| FN | c | 4 | 10 | 15 | 15 | 8 |
| TN | d | 45 | 45 | 42 | 41 | 30 |
| % Sensitivity | a/(a + c) * 100 | 95.06 | 87.65 | 81.48 | 81.48 | 90.12 |
| % Specificity | d/(b + d) * 100 | 100.00 | 100.00 | 93.33 | 91.11 | 66.67 |
| % Concordance | (a + d)/(a + b + c + d) * 100 | 96.83 | 92.06 | 85.71 | 84.92 | 81.75 |
| PPV | a/(a + b) * 100 | 100.00 | 100.00 | 95.65 | 94.29 | 82.95 |
| NPV | d/(c + d) * 100 | 91.84 | 81.82 | 73.68 | 73.21 | 78.95 |
| Total = n | a + b + c + d | 126 | 126 | 126 | 126 | 126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mercado-Uriostegui, M.A.; Castro-Sánchez, L.A.; Batiha, G.E.-S.; Valdez-Espinoza, U.M.; Falcón-Neri, A.; Ramos-Aragon, J.A.; Hernández-Ortiz, R.; Kawazu, S.-I.; Igarashi, I.; Mosqueda, J. The GP-45 Protein, a Highly Variable Antigen from Babesia bigemina, Contains Conserved B-Cell Epitopes in Geographically Distant Isolates. Pathogens 2022, 11, 591. https://doi.org/10.3390/pathogens11050591
Mercado-Uriostegui MA, Castro-Sánchez LA, Batiha GE-S, Valdez-Espinoza UM, Falcón-Neri A, Ramos-Aragon JA, Hernández-Ortiz R, Kawazu S-I, Igarashi I, Mosqueda J. The GP-45 Protein, a Highly Variable Antigen from Babesia bigemina, Contains Conserved B-Cell Epitopes in Geographically Distant Isolates. Pathogens. 2022; 11(5):591. https://doi.org/10.3390/pathogens11050591
Chicago/Turabian StyleMercado-Uriostegui, Miguel Angel, Luis Alberto Castro-Sánchez, Gaber El-Saber Batiha, Uriel Mauricio Valdez-Espinoza, Alfonso Falcón-Neri, Juan Alberto Ramos-Aragon, Ruben Hernández-Ortiz, Shin-Ichiro Kawazu, Ikuo Igarashi, and Juan Mosqueda. 2022. "The GP-45 Protein, a Highly Variable Antigen from Babesia bigemina, Contains Conserved B-Cell Epitopes in Geographically Distant Isolates" Pathogens 11, no. 5: 591. https://doi.org/10.3390/pathogens11050591
APA StyleMercado-Uriostegui, M. A., Castro-Sánchez, L. A., Batiha, G. E. -S., Valdez-Espinoza, U. M., Falcón-Neri, A., Ramos-Aragon, J. A., Hernández-Ortiz, R., Kawazu, S. -I., Igarashi, I., & Mosqueda, J. (2022). The GP-45 Protein, a Highly Variable Antigen from Babesia bigemina, Contains Conserved B-Cell Epitopes in Geographically Distant Isolates. Pathogens, 11(5), 591. https://doi.org/10.3390/pathogens11050591