
Point-of-Care Tests for HIV Drug Resistance Monitoring: Advances and Potentials


Abstract
:1. Introduction
2. Technologies Attempted for HIVDR POCT
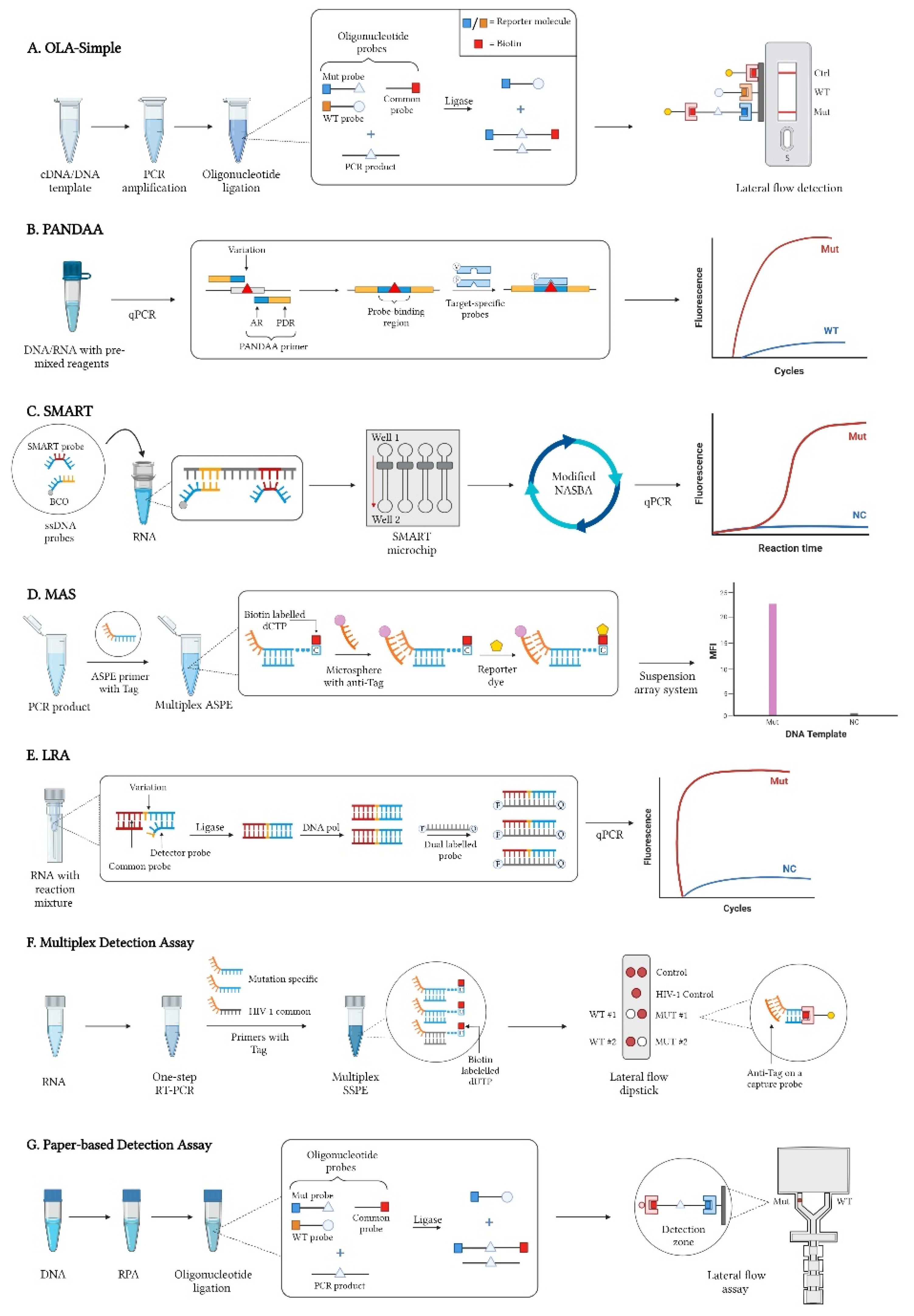
2.1. Oligonucleotide Ligation Assays (OLA)
2.2. Pan-Degenerate Amplification and Adaptation (PANDAA)
2.3. SMART (Simple Method for Amplifying RNA Targets)
2.4. Multiplex Allele-Specific (MAS) Assay
2.5. Ligation on RNA Amplification (LRA)
2.6. Multiplex Detection Assay
2.7. Paper-Based Detection Assay
3. Other Potential POCT Technologies
3.1. Multiplex Solid-Phase Melt Curve Analysis
3.2. µBAR Platform
3.3. Oxford Nanopore MinION (ONT) Sequencing
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- UNAIDS. Global HIV & AIDS Statistics—Fact Sheet; UNAIDS: Geneva, Switzerland, 2022. [Google Scholar]
- UNAIDS. Understanding Fast-Track Acceleratiing Action to End the AIDS Epidemic by 2030; UNAIDS: Geneva, Switzerland, 2015. [Google Scholar]
- UNAIDS. Prevailing against Pandemics by Putting People at the Center _World AIDS Day Report; UNAIDS: Geneva, Switzerland, 2020. [Google Scholar]
- World Health Organization. HIV Drug Resistance Report 2021; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- World Health Organization. Global Action Plan on HIV Drug Resistance 2017–2021; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Inzaule, S.C.; Hamers, R.L.; Paredes, R.; Yang, C.; Schuurman, R.; Rinke de Wit, T.F. The Evolving Landscape of HIV Drug Resistance Diagnostics for Expanding Testing in Resource-Limited Settings. AIDS Rev. 2017, 19, 219–230. [Google Scholar] [PubMed]
- Land, K.J.; Boeras, D.I.; Chen, X.S.; Ramsay, A.R.; Peeling, R.W. REASSURED diagnostics to inform disease control strategies, strengthen health systems and improve patient outcomes. Nat. Microbiol. 2019, 4, 46–54. [Google Scholar] [CrossRef]
- Duarte, H.A.; Panpradist, N.; Beck, I.A.; Lutz, B.; Lai, J.; Kanthula, R.M.; Kantor, R.; Tripathi, A.; Saravanan, S.; MacLeod, I.J.; et al. Current Status of Point-of-Care Testing for Human Immunodeficiency Virus Drug Resistance. J. Infect. Dis. 2017, 216, S824–S828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inzaule, S.C.; Ondoa, P.; Peter, T.; Mugyenyi, P.N.; Stevens, W.S.; de Wit, T.F.R.; Hamers, R.L. Affordable HIV drug-resistance testing for monitoring of antiretroviral therapy in sub-Saharan Africa. Lancet Infect. Dis. 2016, 16, e267–e275. [Google Scholar] [CrossRef]
- Noguera-Julian, M. HIV drug resistance testing—The quest for Point-of-Care. EBioMedicine 2019, 50, 11–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landegren, U.; Kaiser, R.; Sanders, J.; Hood, L. A ligase-mediated gene detection technique. Science 1988, 241, 1077–1080. [Google Scholar] [CrossRef]
- Panpradist, N.; Beck, I.A.; Chung, M.H.; Kiarie, J.N.; Frenkel, L.M.; Lutz, B.R. Simplified Paper Format for Detecting HIV Drug Resistance in Clinical Specimens by Oligonucleotide Ligation. PLoS ONE 2016, 11, e0145962. [Google Scholar] [CrossRef] [Green Version]
- Beck, I.A.; Crowell, C.; Kittoe, R.; Bredell, H.; Machaba, M.; Willamson, C.; Janssens, W.; Jallow, S.; van der Groen, G.; Shao, Y.; et al. Optimization of the oligonucleotide ligation assay, a rapid and inexpensive test for detection of HIV-1 drug resistance mutations, for non-North American variants. J. Acquir. Immune Defic. Syndr. 2008, 48, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Beck, I.A.; Mahalanabis, M.; Pepper, G.; Wright, A.; Hamilton, S.; Langston, E.; Frenkel, L.M. Rapid and sensitive oligonucleotide ligation assay for detection of mutations in human immunodeficiency virus type 1 associated with high-level resistance to protease inhibitors. J. Clin. Microbiol. 2002, 40, 1413–1419. [Google Scholar] [CrossRef] [Green Version]
- Edelstein, R.E.; Nickerson, D.A.; Tobe, V.O.; Manns-Arcuino, L.A.; Frenkel, L.M. Oligonucleotide ligation assay for detecting mutations in the human immunodeficiency virus type 1 pol gene that are associated with resistance to zidovudine, didanosine, and lamivudine. J. Clin. Microbiol. 1998, 36, 569–572. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, L.M.; Wagner, L.E.; Atwood, S.M.; Cummins, T.J.; Dewhurst, S. Specific, sensitive, and rapid assay for human immunodeficiency virus type 1 pol mutations associated with resistance to zidovudine and didanosine. J. Clin. Microbiol. 1995, 33, 342–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.H.; Beck, I.A.; Dross, S.; Tapia, K.; Kiarie, J.N.; Richardson, B.A.; Overbaugh, J.; Sakr, S.R.; John-Stewart, G.C.; Frenkel, L.M. Oligonucleotide ligation assay detects HIV drug resistance associated with virologic failure among antiretroviral-naive adults in Kenya. J. Acquir. Immune Defic. Syndr. 2014, 67, 246–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jourdain, G.; Wagner, T.A.; Ngo-Giang-Huong, N.; Sirirungsi, W.; Klinbuayaem, V.; Fregonese, F.; Nantasen, I.; Techapornroong, M.; Halue, G.; Nilmanat, A.; et al. Association between detection of HIV-1 DNA resistance mutations by a sensitive assay at initiation of antiretroviral therapy and virologic failure. Clin. Infect. Dis. 2010, 50, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Mutsvangwa, J.; Beck, I.A.; Gwanzura, L.; Manhanzva, M.T.; Stranix-Chibanda, L.; Chipato, T.; Frenkel, L.M. Optimization of the oligonucleotide ligation assay for the detection of nevirapine resistance mutations in Zimbabwean Human Immunodeficiency Virus type-1 subtype C. J. Virol. Methods 2014, 210, 36–39. [Google Scholar] [CrossRef] [Green Version]
- Van Dyke, R.B.; Ngo-Giang-Huong, N.; Shapiro, D.E.; Frenkel, L.; Britto, P.; Roongpisuthipong, A.; Beck, I.A.; Yuthavisuthi, P.; Prommas, S.; Puthanakit, T.; et al. A comparison of 3 regimens to prevent nevirapine resistance mutations in HIV-infected pregnant women receiving a single intrapartum dose of nevirapine. Clin. Infect. Dis. 2012, 54, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Panpradist, N.; Beck, I.A.; Vrana, J.; Higa, N.; McIntyre, D.; Ruth, P.S.; So, I.; Kline, E.C.; Kanthula, R.; Wong-On-Wing Lim, J.; et al. OLA-Simple: A software-guided HIV-1 drug resistance test for low-resource laboratories. EBioMedicine 2019, 50, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Panpradist, N.; Beck, I.A.; Ruth, P.S.; Avila-Rios, S.; Garcia-Morales, C.; Soto-Nava, M.; Tapia-Trejo, D.; Matias-Florentino, M.; Paz-Juarez, H.E.; Del Arenal-Sanchez, S.; et al. Near point-of-care, point-mutation test to detect drug resistance in HIV-1: A validation study in a Mexican cohort. AIDS 2020, 34, 1331–1338. [Google Scholar] [CrossRef]
- MacLeod, I.J.; Rowley, C.F.; Essex, M. PANDAA intentionally violates conventional qPCR design to enable durable, mismatch-agnostic detection of highly polymorphic pathogens. Commun. Biol. 2021, 4, 227. [Google Scholar] [CrossRef]
- Maruapula, D.; MacLeod, I.J.; Moyo, S.; Musonda, R.; Seatla, K.; Molebatsi, K.; Leteane, M.; Essex, M.; Gaseitsiwe, S.; Rowley, C.F. Use of a mutation-specific genotyping method to assess for HIV-1 drug resistance in antiretroviral-naive HIV-1 Subtype C-infected patients in Botswana. AAS Open Res. 2020, 3, 50. [Google Scholar] [CrossRef]
- Kouamou, V.; Manasa, J.; Katzenstein, D.; McGregor, A.M.; Ndhlovu, C.E.; Makadzange, T. Diagnostic Accuracy of Pan-Degenerate Amplification and Adaptation Assay for HIV-1 Drug Resistance Mutation Analysis in Low- and Middle-Income Countries. J. Clin. Microbiol. 2020, 58, e01045-20. [Google Scholar] [CrossRef]
- Chung, M.H.; McGrath, C.J.; Beck, I.A.; Levine, M.; Milne, R.S.; So, I.; Andersen, N.; Dross, S.; Coombs, R.W.; Chohan, B.; et al. Evaluation of the management of pretreatment HIV drug resistance by oligonucleotide ligation assay: A randomised controlled trial. Lancet HIV 2020, 7, e104–e112. [Google Scholar] [CrossRef]
- McCalla, S.E.; Ong, C.; Sarma, A.; Opal, S.M.; Artenstein, A.W.; Tripathi, A. A simple method for amplifying RNA targets (SMART). J. Mol. Diagn. 2012, 14, 328–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morabito, K.; Kantor, R.; Tai, W.; Schreier, L.; Tripathi, A. Detection of HIV-1 minority variants containing the K103N drug-resistance mutation using a simple method to amplify RNA targets (SMART). J. Mol. Diagn. 2013, 15, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tai, W.; Angione, S.L.; John, A.R.; Opal, S.M.; Artenstein, A.W.; Tripathi, A. Subtyping clinical specimens of influenza A virus by use of a simple method to amplify RNA targets. J. Clin. Microbiol. 2013, 51, 3324–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunbar, S.A. Applications of Luminex xMAP technology for rapid, high-throughput multiplexed nucleic acid detection. Clin. Chim. Acta 2006, 363, 71–82. [Google Scholar] [CrossRef]
- Zhang, G.; Cai, F.; Zhou, Z.; DeVos, J.; Wagar, N.; Diallo, K.; Zulu, I.; Wadonda-Kabondo, N.; Stringer, J.S.; Weidle, P.J.; et al. Simultaneous detection of major drug resistance mutations in the protease and reverse transcriptase genes for HIV-1 subtype C by use of a multiplex allele-specific assay. J. Clin. Microbiol. 2013, 51, 3666–3674. [Google Scholar] [CrossRef] [Green Version]
- Houser, B. Bio-Rad’s Bio-Plex(R) suspension array system, xMAP technology overview. Arch. Physiol. Biochem. 2012, 118, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Cai, F.; de Rivera, I.L.; Zhou, Z.; Zhang, J.; Nkengasong, J.; Gao, F.; Yang, C. Simultaneous Detection of Major Drug Resistance Mutations of HIV-1 Subtype B Viruses from Dried Blood Spot Specimens by Multiplex Allele-Specific Assay. J. Clin. Microbiol. 2016, 54, 220–222. [Google Scholar] [CrossRef] [Green Version]
- Barany, F. Genetic disease detection and DNA amplification using cloned thermostable ligase. Proc. Natl. Acad. Sci. USA 1991, 88, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, M.; Antson, D.O.; Barbany, G.; Landegren, U. RNA-templated DNA ligation for transcript analysis. Nucleic Acids Res. 2001, 29, 578–581. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Li, Z.; Liu, C.; Cheng, Y. Simple and sensitive detection of microRNAs with ligase chain reaction. Chem. Commun. 2010, 46, 2432–2434. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, X.; Li, Z.; Jiao, X.; Wang, Y.; Zhang, Y. Highly sensitive determination of microRNA using target-primed and branched rolling-circle amplification. Angew. Chem. Int. Ed. Engl. 2009, 48, 3268–3272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, J.; Coetzer, M.; Angione, S.; Kantor, R.; Tripathi, A. One-Step Ligation on RNA Amplification for the Detection of Point Mutations. J. Mol. Diagn. 2015, 17, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Martinez, J.; Silvy, M.; Chiaroni, J.; Fournier-Wirth, C.; Roubinet, F.; Bailly, P.; Bres, J.C. Multiplex Lateral Flow Assay for Rapid Visual Blood Group Genotyping. Anal. Chem. 2018, 90, 7502–7509. [Google Scholar] [CrossRef]
- Fountoglou, N.; Petropoulou, M.; Iliadi, A.; Christopoulos, T.K.; Ioannou, P.C. Tauwo-panel molecular testing for genetic predisposition for thrombosis using multi-allele visual biosensors. Anal. Bioanal. Chem. 2016, 408, 1943–1952. [Google Scholar] [CrossRef]
- Petropoulou, M.; Poula, A.; Traeger-Synodinos, J.; Kanavakis, E.; Christopoulos, T.K.; Ioannou, P.C. Multi-allele DNA biosensor for the rapid genotyping of ‘nondeletion’ alpha thalassaemia mutations in HBA1 and HBA2 genes by means of multiplex primer extension reaction. Clin. Chim. Acta 2015, 446, 241–247. [Google Scholar] [CrossRef]
- Papanikos, F.; Iliadi, A.; Petropoulou, M.; Ioannou, P.C.; Christopoulos, T.K.; Kanavakis, E.; Traeger-Synodinos, J. Lateral flow dipstick test for genotyping of 15 beta-globin gene (HBB) mutations with naked-eye detection. Anal. Chim. Acta 2012, 727, 61–66. [Google Scholar] [CrossRef]
- Gomez-Martinez, J.; Foulongne, V.; Laureillard, D.; Nagot, N.; Montes, B.; Cantaloube, J.F.; Van de Perre, P.; Fournier-Wirth, C.; Moles, J.P.; Bres, J.C. Near-point-of-care assay with a visual readout for detection of HIV-1 drug resistance mutations: A proof-of-concept study. Talanta 2021, 231, 122378. [Google Scholar] [CrossRef]
- Bui, M.H.; Stone, G.G.; Nilius, A.M.; Almer, L.; Flamm, R.K. PCR-oligonucleotide ligation assay for detection of point mutations associated with quinolone resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2003, 47, 1456–1459. [Google Scholar] [CrossRef] [Green Version]
- Natoli, M.E.; Rohrman, B.A.; De, S.C.; van Zyl, G.U.; Richards-Kortum, R.R. Paper-based detection of HIV-1 drug resistance using isothermal amplification and an oligonucleotide ligation assay. Anal. Biochem. 2018, 544, 64–71. [Google Scholar] [CrossRef]
- Clutter, D.S.; Mazarei, G.; Sinha, R.; Manasa, J.; Nouhin, J.; LaPrade, E.; Bolouki, S.; Tzou, P.L.; Hannita-Hui, J.; Sahoo, M.K.; et al. Multiplex Solid-Phase Melt Curve Analysis for the Point-of-Care Detection of HIV-1 Drug Resistance. J. Mol. Diagn. 2019, 21, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Myers, F.B.; Henrikson, R.H.; Xu, L.; Lee, L.P. A point-of-care instrument for rapid multiplexed pathogen genotyping. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2011, 2011, 3668–3671. [Google Scholar] [PubMed]
- Myers, F.B.; Henrikson, R.H.; Bone, J.M.; Lee, L.P. A handheld point-of-care genomic diagnostic system. PLoS ONE 2013, 8, e70266. [Google Scholar] [CrossRef]
- Gonzalez, C.; Gondola, J.; Ortiz, A.Y.; Castillo, J.; Pascale, J.M.; Martinez, A.A. Barcoding analysis of HIV drug resistance mutations using Oxford Nanopore MinION (ONT) sequencing. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| POCTs | Starting Material | Subtype Specificity | Major Equipment Required | Validated against Sanger | Refs. |
|---|---|---|---|---|---|
| OLA-Simple | DNA or RNA | HIV-1 (A, B, C, D, AE) | Thermocycler, Office scanner | ✓ | [16] |
| PANDAA | DNA or RNA | Subtype independent | qPCR machine | ✓ | [23] |
| SMART | RNA | HIV-1 (any subtype with a K103N region) | Microfluidic device, qPCR machine | 〤 | [27] |
| MAS | DNA | HIV-1 (B, C) | Compact suspension array system | ✓ | [31,33] |
| LRA | RNA | HIV-1 (any subtype with a K103N region) | qPCR machine | 〤 | [38] |
| Multiplex Detection Assay | RNA | HIV-1 (B) | Thermocycler, | ✓ | [43] |
| Dry oven | |||||
| Paper-based Detection Assay | DNA | HIV-1 (B) | Heat block, Thermocycler, Tabletop centrifuge | 〤 | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chua, R.J.; Capiña, R.; Ji, H. Point-of-Care Tests for HIV Drug Resistance Monitoring: Advances and Potentials. Pathogens 2022, 11, 724. https://doi.org/10.3390/pathogens11070724
Chua RJ, Capiña R, Ji H. Point-of-Care Tests for HIV Drug Resistance Monitoring: Advances and Potentials. Pathogens. 2022; 11(7):724. https://doi.org/10.3390/pathogens11070724
Chicago/Turabian StyleChua, Rayeil J., Rupert Capiña, and Hezhao Ji. 2022. "Point-of-Care Tests for HIV Drug Resistance Monitoring: Advances and Potentials" Pathogens 11, no. 7: 724. https://doi.org/10.3390/pathogens11070724
APA StyleChua, R. J., Capiña, R., & Ji, H. (2022). Point-of-Care Tests for HIV Drug Resistance Monitoring: Advances and Potentials. Pathogens, 11(7), 724. https://doi.org/10.3390/pathogens11070724