

Effects of a Novel, Intelligent, pH-Responsive Resin Adhesive on Cariogenic Biofilms In Vitro


 ,
,
Abstract
:1. Introduction
2. Results
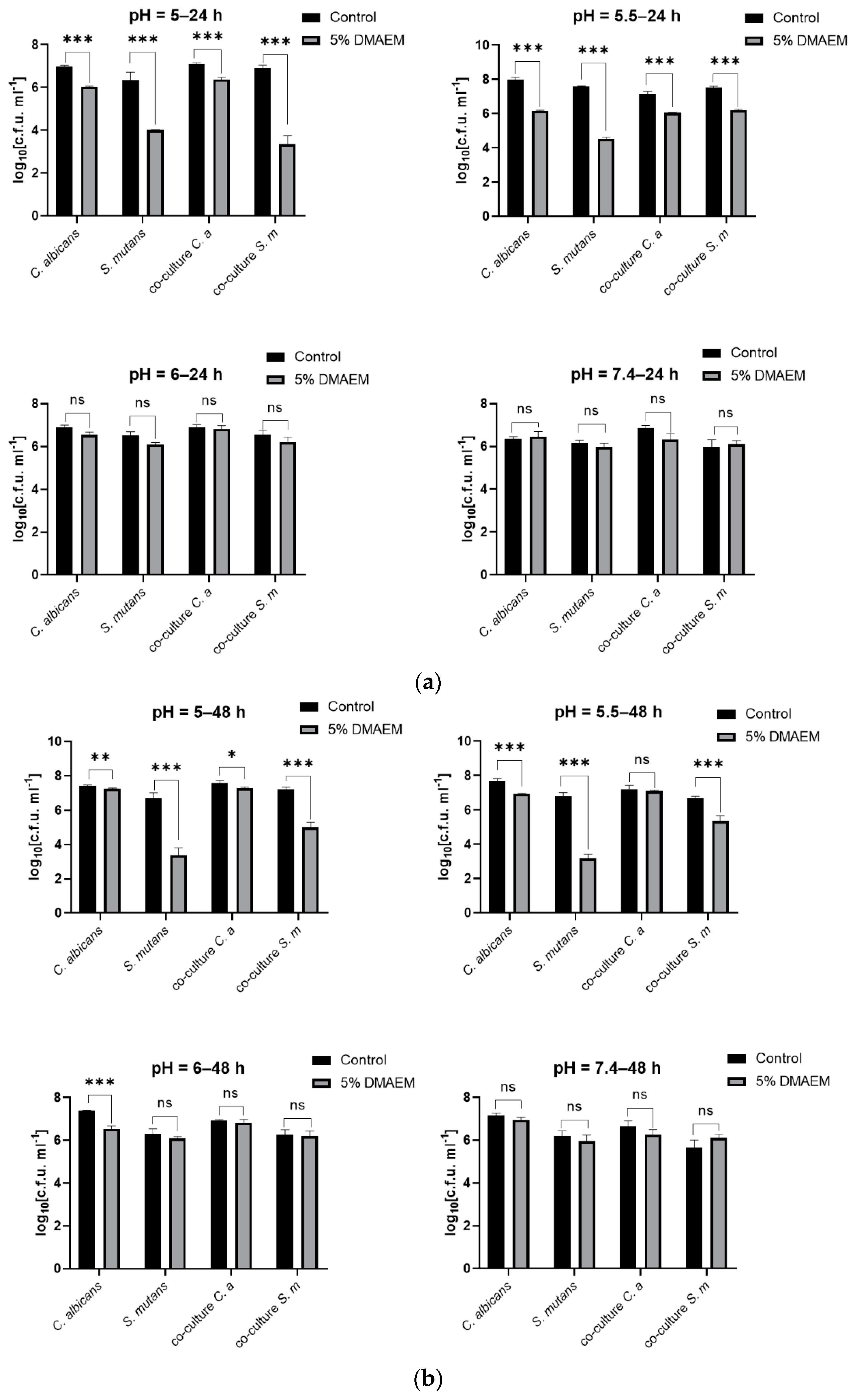
2.1. DMAEM@RA Inhibited the Growth of S. mutans and C. albicans Dual-Species Biofilms in a pH-Dependent Manner
2.2. DMAEM@RA Played pH-Dependent Antibiofilm Roles on S. mutans and C. albicans Dual-Species Biofilms In Vitro
2.3. DMAEM@RA Decreased the Production of EPS of the S. mutans and Dual-Species Biofilms at pH 5 and 5.5
2.4. DMAEM@RA pH-Responsively Reduced the Production of Lactic Acid in S. mutans and C. albicans Dual-Species Biofilms
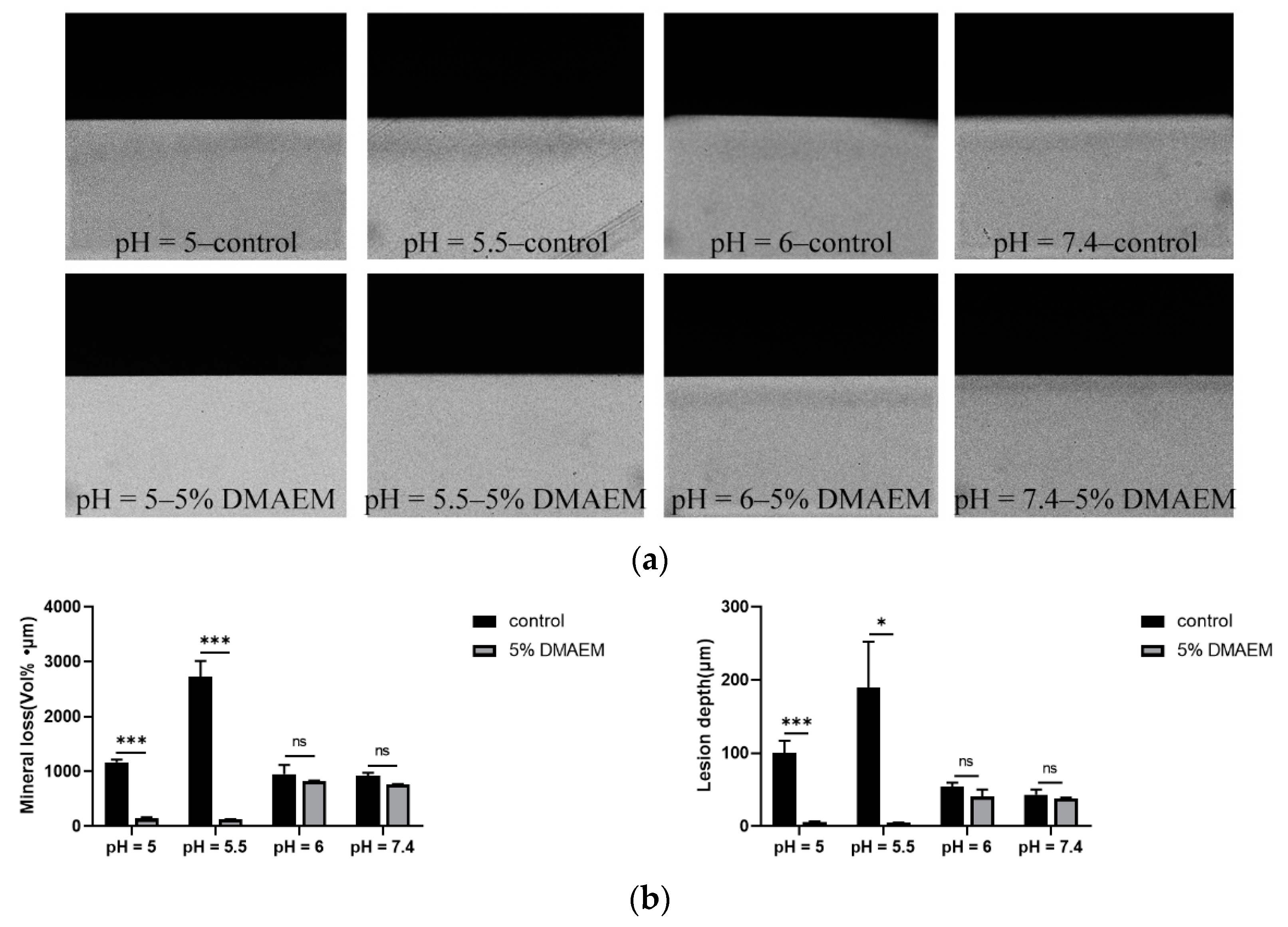
2.5. DMAEM@RA Inhibited Tooth Demineralization Caused by S. mutans and C. albicans Dual-Species Biofilms in a pH-Responsive Way In Vitro
2.6. DMAEM@RA Changed the Expression of the Cariogenic Genes, Virulence-Associated Genes, and pH-Regulated Genes of the Dual-Species Biofilms at pH 5 and 5.5
3. Discussion
4. Materials and Methods
4.1. Preparation of DMAEM@RA
4.2. Bacterial Strains and Growth Conditions
4.3. Colony-Forming Units (CFU) Counts
4.4. Scanning Electron Microscope (SEM) Examination
4.5. Exopolysaccharide (EPS) Staining
4.6. Lactic Acid Measurement
4.7. Transverse Microradiography (TMR) Analysis
4.8. RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental caries. Nat. Rev. Dis. Primers 2017, 3, 17030. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhang, L.; Yue, L.; Ling, J.; Fan, M.; Yang, D.; Huang, Z.; Niu, Y.; Liu, J.; Zhao, J.; et al. Expert consensus on dental caries management. Int. J. Oral. Sci. 2022, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Disease, G.B.D.; Injury, I.; Prevalence, C. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef]
- Heaton, B.; Cherng, S.T.; Sohn, W.; Garcia, R.I.; Galea, S. Complex Systems Model of Dynamic Mechanisms of Early Childhood Caries Development. J. Dent. Res. 2020, 99, 537–543. [Google Scholar] [CrossRef]
- Zou, J.; Meng, M.; Law, C.S.; Rao, Y.; Zhou, X. Common dental diseases in children and malocclusion. Int. J. Oral. Sci. 2018, 10, 7. [Google Scholar] [CrossRef]
- Tinanoff, N.; Baez, R.J.; Diaz Guillory, C.; Donly, K.J.; Feldens, C.A.; McGrath, C.; Phantumvanit, P.; Pitts, N.B.; Seow, W.K.; Sharkov, N.; et al. Early childhood caries epidemiology, aetiology, risk assessment, societal burden, management, education, and policy: Global perspective. Int. J. Paediatr. Dent. 2019, 29, 238–248. [Google Scholar] [CrossRef]
- Qu, X.; Houser, S.H.; Tian, M.; Zhang, Q.; Pan, J.; Zhang, W. Effects of early preventive dental visits and its associations with dental caries experience: A cross-sectional study. BMC Oral. Health 2022, 22, 150. [Google Scholar] [CrossRef]
- Lotto, M.; Strieder, A.P.; Ayala Aguirre, P.E.; Andrade Moreira Machado, M.A.; Rios, D.; Cruvinel, A.; Cruvinel, T. Parental perspectives on early childhood caries: A qualitative study. Int. J. Paediatr. Dent. 2020, 30, 451–458. [Google Scholar] [CrossRef]
- Ferracane, J.L. Resin composite—State of the art. Dent. Mater. 2011, 27, 29–38. [Google Scholar] [CrossRef]
- Opdam, N.J.; van de Sande, F.H.; Bronkhorst, E.; Cenci, M.S.; Bottenberg, P.; Pallesen, U.; Gaengler, P.; Lindberg, A.; Huysmans, M.C.; van Dijken, J.W. Longevity of posterior composite restorations: A systematic review and meta-analysis. J. Dent. Res. 2014, 93, 943–949. [Google Scholar] [CrossRef] [Green Version]
- Metz, I.; Rothmaier, K.; Pitchika, V.; Crispin, A.; Hickel, R.; Garcia-Godoy, F.; Bucher, K.; Kuhnisch, J. Risk factors for secondary caries in direct composite restorations in primary teeth. Int. J. Paediatr. Dent. 2015, 25, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Bucher, K.; Tautz, A.; Hickel, R.; Kuhnisch, J. Longevity of composite restorations in patients with early childhood caries (ECC). Clin. Oral. Investig. 2014, 18, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Jokstad, A. Secondary caries and microleakage. Dent. Mater. 2016, 32, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Metwalli, K.H.; Khan, S.A.; Krom, B.P.; Jabra-Rizk, M.A. Streptococcus mutans, Candida albicans, and the human mouth: A sticky situation. PLoS Pathog. 2013, 9, e1003616. [Google Scholar] [CrossRef]
- Xiao, J.; Huang, X.; Alkhers, N.; Alzamil, H.; Alzoubi, S.; Wu, T.T.; Castillo, D.A.; Campbell, F.; Davis, J.; Herzog, K.; et al. Candida albicans and Early Childhood Caries: A Systematic Review and Meta-Analysis. Caries Res. 2018, 52, 102–112. [Google Scholar] [CrossRef]
- He, Z.; Huang, Z.; Jiang, W.; Zhou, W. Antimicrobial Activity of Cinnamaldehyde on Streptococcus mutans Biofilms. Front. Microbiol. 2019, 10, 2241. [Google Scholar] [CrossRef]
- Pereira, D.; Seneviratne, C.J.; Koga-Ito, C.Y.; Samaranayake, L.P. Is the oral fungal pathogen Candida albicans a cariogen? Oral. Dis. 2018, 24, 518–526. [Google Scholar] [CrossRef]
- Kim, H.E.; Liu, Y.; Dhall, A.; Bawazir, M.; Koo, H.; Hwang, G. Synergism of Streptococcus mutans and Candida albicans Reinforces Biofilm Maturation and Acidogenicity in Saliva: An In Vitro Study. Front. Cell Infect. Microbiol. 2020, 10, 623980. [Google Scholar] [CrossRef]
- Koo, H.; Bowen, W.H. Candida albicans and Streptococcus mutans: A potential synergistic alliance to cause virulent tooth decay in children. Future Microbiol. 2014, 9, 1295–1297. [Google Scholar] [CrossRef]
- Xiao, J.; Grier, A.; Faustoferri, R.C.; Alzoubi, S.; Gill, A.L.; Feng, C.; Liu, Y.; Quivey, R.G.; Kopycka-Kedzierawski, D.T.; Koo, H.; et al. Association between Oral Candida and Bacteriome in Children with Severe ECC. J. Dent. Res. 2018, 97, 1468–1476. [Google Scholar] [CrossRef]
- Hwang, G.; Liu, Y.; Kim, D.; Li, Y.; Krysan, D.J.; Koo, H. Candida albicans mannans mediate Streptococcus mutans exoenzyme GtfB binding to modulate cross-kingdom biofilm development in vivo. PLoS Pathog. 2017, 13, e1006407. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, E.; Parsaei, Y.; Klein, M.I.; Koo, H. Advances in the microbial etiology and pathogenesis of early childhood caries. Mol. Oral. Microbiol. 2017, 32, 24–34. [Google Scholar] [CrossRef]
- Bowen, W.H. The Stephan Curve revisited. Odontology 2013, 101, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhou, Y.; Zhou, X.; Liao, B.; Xu, H.H.K.; Chu, C.H.; Cheng, L.; Ren, B. Dimethylaminododecyl methacrylate inhibits Candida albicans and oropharyngeal candidiasis in a pH-dependent manner. Appl. Microbiol. Biotechnol. 2020, 104, 3585–3595. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ding, C.; Wang, Y.; Tan, H.; Li, J. pH-Responsive polymeric nanocarriers for efficient killing of cariogenic bacteria in biofilms. Biomater Sci. 2019, 7, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Horev, B.; Klein, M.I.; Hwang, G.; Li, Y.; Kim, D.; Koo, H.; Benoit, D.S. pH-activated nanoparticles for controlled topical delivery of farnesol to disrupt oral biofilm virulence. ACS Nano. 2015, 9, 2390–2404. [Google Scholar] [CrossRef]
- Zhou, Z.; Hu, F.; Hu, S.; Kong, M.; Feng, C.; Liu, Y.; Cheng, X.; Ji, Q.; Chen, X. pH-Activated nanoparticles with tar-geting for the treatment of oral plaque biofilm. J. Mater. Chem. B 2018, 6, 586–592. [Google Scholar] [CrossRef]
- Liang, J.; Liu, F.; Zou, J.; Xu, H.H.K.; Han, Q.; Wang, Z.; Li, B.; Yang, B.; Ren, B.; Li, M.; et al. pH-Responsive Antibacterial Resin Adhesives for Secondary Caries Inhibition. J. Dent. Res. 2020, 99, 1368–1376. [Google Scholar] [CrossRef]
- Popolo, L.; Degani, G.; Camilloni, C.; Fonzi, W.A. The PHR Family: The Role of Extracellular Transglycosylases in Shaping Candida albicans Cells. J. Fungi 2017, 3, 59. [Google Scholar] [CrossRef]
- Degani, G.; Popolo, L. The Glucan-Remodeling Enzyme Phr1p and the Chitin Synthase Chs1p Cooperate to Maintain Proper Nuclear Segregation and Cell Integrity in Candida albicans. Front. Cell Infect. Microbiol. 2019, 9, 400. [Google Scholar] [CrossRef] [Green Version]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef]
- Fernandes, R.A.; Monteiro, D.R.; Arias, L.S.; Fernandes, G.L.; Delbem, A.C.; Barbosa, D.B. Biofilm formation by Candida albicans and Streptococcus mutans in the presence of farnesol: A quantitative evaluation. Biofouling 2016, 32, 329–338. [Google Scholar] [CrossRef]
- Garcia, B.A.; Acosta, N.C.; Tomar, S.L.; Roesch, L.F.W.; Lemos, J.A.; Mugayar, L.R.F.; Abranches, J. Association of Candida albicans and Cbp(+) Streptococcus mutans with early childhood caries recurrence. Sci. Rep. 2021, 11, 10802. [Google Scholar] [CrossRef]
- Hwang, G.; Marsh, G.; Gao, L.; Waugh, R.; Koo, H. Binding Force Dynamics of Streptococcus mu-tans-glucosyltransferase B to Candida albicans. J. Dent. Res. 2015, 94, 1310–1317. [Google Scholar] [CrossRef] [PubMed]
- Ellepola, K.; Liu, Y.; Cao, T.; Koo, H.; Seneviratne, C.J. Bacterial GtfB Augments Candida albicans Accumulation in Cross-Kingdom Biofilms. J. Dent. Res. 2017, 96, 1129–1135. [Google Scholar] [CrossRef]
- Koo, H.; Andes, D.R.; Krysan, D.J. Candida-streptococcal interactions in biofilm-associated oral diseases. PLoS Pathog. 2018, 14, e1007342. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.I.V.; Rinaldi, T.B.; Christiano, C.M.S.; De Sales Leite, L.; Barbugli, P.A.; Klein, M.I. Dual-species biofilms of Streptococcus mutans and Candida albicans exhibit more biomass and are mutually beneficial compared with sin-gle-species biofilms. J. Oral. Microbiol. 2019, 11, 1581520. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Scoffield, J.; Wu, R.; Deivanayagam, C.; Zou, J.; Wu, H. Antigen I/II mediates interactions between Strepto-coccus mutans and Candida albicans. Mol. Oral. Microbiol. 2018, 33, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.R.; Yang, W.D.; Niu, K.X.; Wang, B.; Wang, W.M. An Update on the Evolution of Glucosyltransferase (Gtf) Genes in Streptococcus. Front. Microbiol. 2018, 9, 2979. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Fox, E.P.; Nett, J.E.; Sorrells, T.R.; Mitrovich, Q.M.; Hernday, A.D.; Tuch, B.B.; Andes, D.R.; Johnson, A.D. A recently evolved transcriptional network controls biofilm development in Candida albicans. Cell 2012, 148, 126–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazier, V.E.; Murante, T.; Murante, D.; Koselny, K.; Liu, Y.; Kim, D.; Koo, H.; Krysan, D.J. Genetic analysis of the Candida albicans biofilm transcription factor network using simple and complex haploinsufficiency. PLoS Genet. 2017, 13, e1006948. [Google Scholar] [CrossRef]
- Glazier, V.E. EFG1, Everyone’s Favorite Gene in Candida albicans: A Comprehensive Literature Review. Front. Cell Infect. Microbiol. 2022, 12, 855229. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.S.; Hancock, T.J.; Lumsdaine, S.W.; Kauffman, S.J.; Mangrum, M.M.; Phillips, E.K.; Sparer, T.E.; Reynolds, T.B. Activation of Cph1 causes β(1,3)-glucan unmasking in Candida albicans and attenuates virulence in mice in a neutrophil-dependent manner. PLoS Pathog. 2021, 17, e1009839. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Ren, B.; He, J.; Peng, X.; Guo, Q.; Zheng, L.; Li, J.; Dai, H.; Chen, V.; Zhang, L.; et al. Candida albicans promotes tooth decay by inducing oral microbial dysbiosis. ISME J. 2021, 15, 894–908. [Google Scholar] [CrossRef]
- Kovacova, K.; Degani, G.; Stratilova, E.; Farkas, V.; Popolo, L. Catalytic properties of Phr family members of cell wall glucan remodeling enzymes: Implications for the adaptation of Candida albicans to ambient pH. FEMS Yeast Res. 2015, 15, fou011. [Google Scholar] [CrossRef]
- Liang, J.; Li, M.; Ren, B.; Wu, T.; Xu, H.H.K.; Liu, Y.; Peng, X.; Yang, G.; Weir, M.D.; Zhang, S.; et al. The anti-caries effects of dental adhesive resin influenced by the position of functional groups in quaternary ammonium monomers. Dent. Mater. 2018, 34, 400–411. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, K.; Zhou, X.; Xu, N.; Xu, H.H.; Weir, M.D.; Ge, Y.; Wang, S.; Li, M.; Li, Y.; et al. Anti-bacterial effect of dental adhesive containing dimethylaminododecyl methacrylate on the development of Streptococcus mutans biofilm. Int. J. Mol. Sci. 2014, 15, 12791–12806. [Google Scholar] [CrossRef] [PubMed]
- Sztajer, H.; Szafranski, S.P.; Tomasch, J.; Reck, M.; Nimtz, M.; Rohde, M.; Wagner-Dobler, I. Cross-feeding and in-terkingdom communication in dual-species biofilms of Streptococcus mutans and Candida albicans. ISME J. 2014, 8, 2256–2271. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Weir, M.D.; Xu, H.H.; Antonucci, J.M.; Kraigsley, A.M.; Lin, N.J.; Lin-Gibson, S.; Zhou, X. Antibacterial amorphous calcium phosphate nanocomposites with a quaternary ammonium dimethacrylate and silver nanoparticles. Dent. Mater. 2012, 28, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, W.; Zhang, M.; Zhang, L.; Shen, Y.; Huang, S.; Li, M.; Qiu, W.; Pan, Y.; Zhou, L.; et al. The Inhibitory Effects of Ficin on Streptococcus mutans Biofilm Formation. Biomed. Res. Int. 2021, 2021, 6692328. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Cheng, L.; Liao, B.; Shi, Y.; Niu, Y.; Zhu, C.; Ye, X.; Zhou, X.; Ren, B. Candida albicans CHK1 gene from two-component system is essential for its pathogenicity in oral candidiasis. Appl. Microbiol. Biotechnol. 2021, 105, 2485–2496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wang, S.; Zhou, X.; Xu, H.H.; Weir, M.D.; Ge, Y.; Li, M.; Wang, S.; Li, Y.; Xu, X.; et al. Effect of antibacterial dental adhesive on multispecies biofilms formation. J. Dent. Res. 2015, 94, 622–629. [Google Scholar] [CrossRef]
- Huang, Y.; Li, H.; Zhu, C.G.; Zhou, X.; Wang, H.; Han, Q.; Ren, B.; Cheng, L. Anti-bacterial and anti-microbial aging effects of resin-based sealant modified by quaternary ammonium monomers. J. Dent. 2021, 112, 103767. [Google Scholar] [CrossRef]
- He, L.; Deng, D.; Zhou, X.; Cheng, L.; ten Cate, J.M.; Li, J.; Li, X.; Crielaard, W. Novel tea polyphenol-modified calcium phosphate nanoparticle and its remineralization potential. J. Biomed. Mater Res. B Appl. Biomater 2015, 103, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Li, B.; Zhou, X.; Ge, Y.; Wang, S.; Li, M.; Ren, B.; Wang, H.; Zhang, K.; Xu, H.H.K.; et al. Anti-Caries Effects of Dental Adhesives Containing Quaternary Ammonium Methacrylates with Different Chain Lengths. Materials 2017, 10, 643. [Google Scholar] [CrossRef]
- Deng, Y.; Yang, Y.; Zhang, B.; Chen, H.; Lu, Y.; Ren, S.; Lei, L.; Hu, T. The vicK gene of Streptococcus mutans mediates its cariogenicity via exopolysaccharides metabolism. Int. J. Oral. Sci. 2021, 13, 45. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, Y.; Han, Q.; Ye, X.; Chen, Y.; Sun, Y.; Liu, Y.; Zou, J.; Qi, G.; Zhou, X.; et al. Synonymous point mutation of gtfB gene caused by therapeutic X-rays exposure reduced the biofilm formation and cariogenic abilities of Streptococcus mutans. Cell Biosci. 2021, 11, 91. [Google Scholar] [CrossRef]
- Yang, W.; Zhou, Y.; Wu, C.; Tang, J. Enterohemorrhagic Escherichia coli promotes the invasion and tissue damage of enterocytes infected with Candida albicans in vitro. Sci. Rep. 2016, 6, 37485. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′→3′) | Template Strand |
|---|---|---|
| 16S rRNA | F R | AGCGTTGTCCGGATTTATTG CTACGCATTTCACCGCTACA |
| gtfB | F R | CACTATCGGCGGTTACGAAT CAATTTGGAGCAAGTCAGCA |
| gtfC | F R | GATGCTGCAAACTTCGAACA TATTGACGCTGCGTTTCTTG |
| gtfD | F | TTGACGGTGTTCGTGTTGAT |
| R | AAAGCGATAGGCGCAGTTTA | |
| 18S rRNA | F | TCTTTCTTGATTTTGTGGGTGG |
| R | TCGATAGTCCCTCTAAGAAGTG | |
| EFG1 | F | ACGTGGTAGAAGAGATGGGA |
| R | TGCATTAGGAGTTACTCCGG | |
| CPH1 | F | GGTGGCGGCAGTGATAGTG |
| R | GTGTACTCCGGTGACGATTTTTC | |
| PHR1 | F | GGTTTGGTTCTGGTTGATGG |
| R | AGCAGCAGTTCCTGGACATT | |
| PHR2 | F | CTCCTCCATTTCCAGAACCA |
| R | CGTCTGAATCAACCTTGTCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Liang, J.; Zhou, X.; Ren, B.; Wang, H.; Han, Q.; Li, H.; Cheng, L. Effects of a Novel, Intelligent, pH-Responsive Resin Adhesive on Cariogenic Biofilms In Vitro. Pathogens 2022, 11, 1014. https://doi.org/10.3390/pathogens11091014
Shi Y, Liang J, Zhou X, Ren B, Wang H, Han Q, Li H, Cheng L. Effects of a Novel, Intelligent, pH-Responsive Resin Adhesive on Cariogenic Biofilms In Vitro. Pathogens. 2022; 11(9):1014. https://doi.org/10.3390/pathogens11091014
Chicago/Turabian StyleShi, Yangyang, Jingou Liang, Xuedong Zhou, Biao Ren, Haohao Wang, Qi Han, Hao Li, and Lei Cheng. 2022. "Effects of a Novel, Intelligent, pH-Responsive Resin Adhesive on Cariogenic Biofilms In Vitro" Pathogens 11, no. 9: 1014. https://doi.org/10.3390/pathogens11091014
APA StyleShi, Y., Liang, J., Zhou, X., Ren, B., Wang, H., Han, Q., Li, H., & Cheng, L. (2022). Effects of a Novel, Intelligent, pH-Responsive Resin Adhesive on Cariogenic Biofilms In Vitro. Pathogens, 11(9), 1014. https://doi.org/10.3390/pathogens11091014