
Identification of Potential Hazards Associated with South Korean Prawns and Monitoring Results Targeting Fishing Bait

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Preparation
2.1.1. Sampling
2.1.2. Preparation of Tissues for Analysis
2.2. Identification of Hazardous Factors of Imported Prawns
2.3. Molecular Biological Analysis
2.3.1. DNA Extraction
2.3.2. Conventional PCR for Pathogen Detection and Real-Time PCR for Viral Copies
2.3.3. Definitive Diagnosis
2.4. Histopathological Analysis
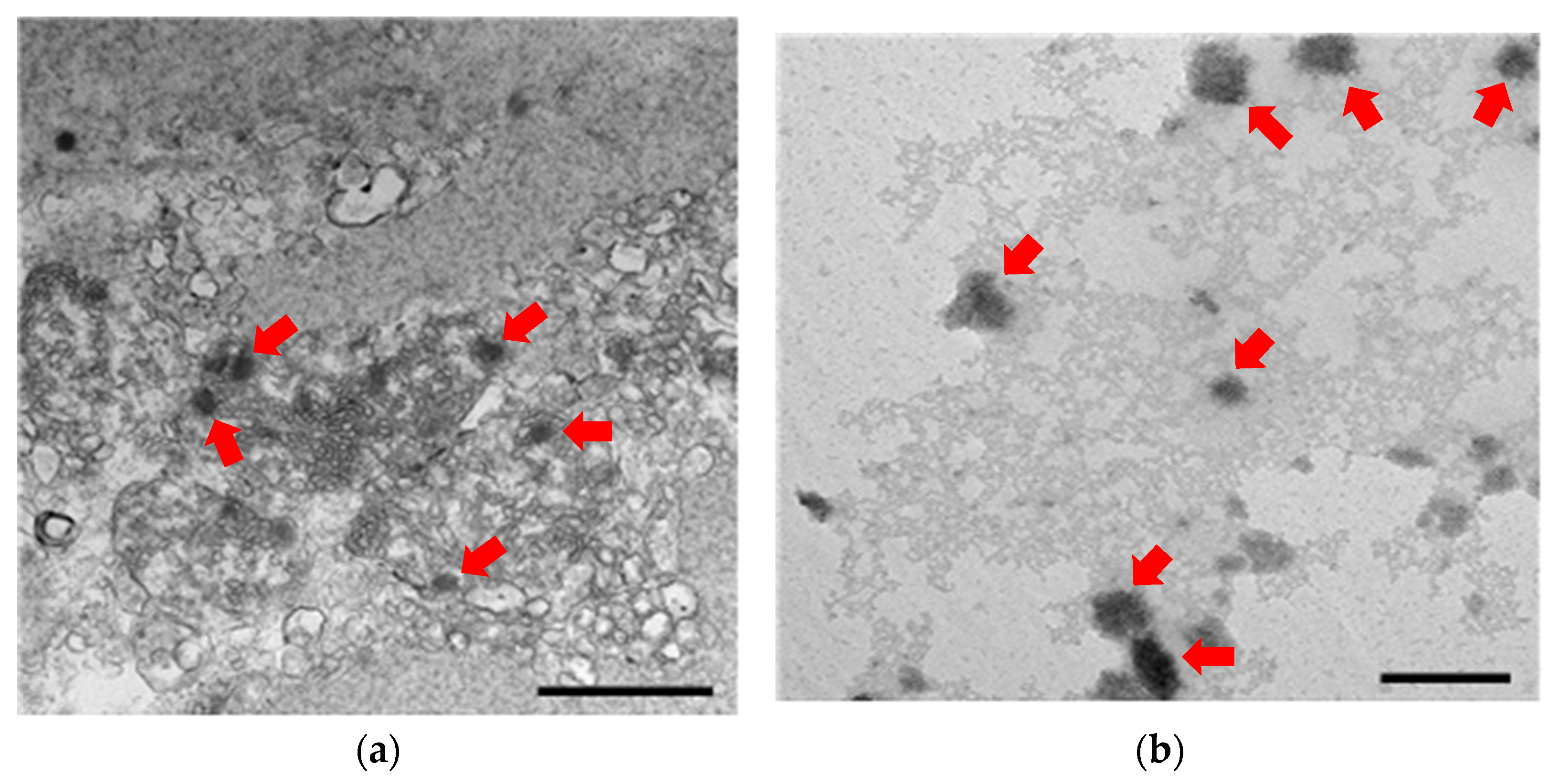
2.5. Transmission Electron Microscopy (TEM) Analysis
2.6. Infectiousness Assessment
3. Results
3.1. Sample Acquisition Status
3.2. Pathogen Detection
3.2.1. Identification of Hazardous Factors
3.2.2. Prevalence of VpAHPND and WSSV
3.2.3. Viral Copies of WSSV
3.3. Infectiousness Assessment of Detected Pathogens
3.3.1. Infectiousness Assessment of VpAHPND
3.3.2. Infectiousness Assessment of WSSV
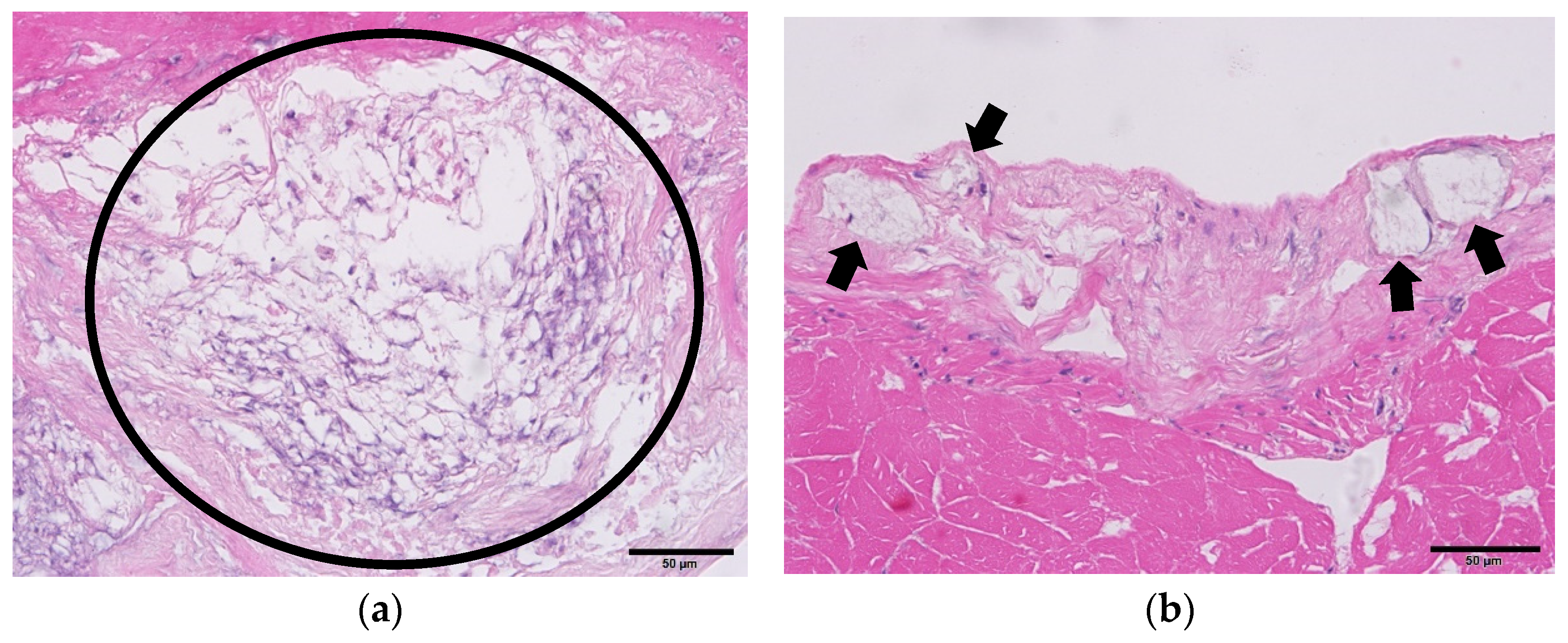
3.4. Histopathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WTO; SPS. G/SPS/N/KOR/692. Available online: https://docs.wto.org/dol2fe/Pages/FE_Search/FE_S_S009-DP.aspx?language=E&HasEnglishRecord=True&HasFrenchRecord=True&HasSpanishRecord=True&CatalogueIdList=265503,265504,265505,265110,265020,265023,264848,264849,264837,264776&CurrentCatalogueIdIndex=3&FullTextHash=371857150 (accessed on 1 March 2023).
- WTO; SPS. G/SPS/N/TPKM/531. Available online: https://docs.wto.org/dol2fe/Pages/FE_Search/FE_S_S009-DP.aspx?language=E&CatalogueIdList=264269,264195,264036,263563,263350,263356,263359,263112,263053,263055&CurrentCatalogueIdIndex=1&FullTextHash=1&HasEnglishRecord=True&HasFrenchRecord=True&HasSpanishRecord=True (accessed on 1 March 2023).
- Flegel, T.W. Review of disease transmission risks from prawn products exported for human consumption. Aquaculture 2009, 290, 179–189. [Google Scholar] [CrossRef]
- Kim, D.-H. Low-value fish used as feed is a source of disease in farmed fish. Fish. Aquat. Sci. 2015, 18, 203–209. [Google Scholar] [CrossRef]
- Qiu, L.; Dong, X.; Wan, X.Y.; Huang, J. Analysis of iridescent viral disease of shrimp (SHID) in 2017. In Analysis of Important Diseases of Aquatic Animals in China in 2017; Fishery and Fishery Administration Bureau under the Ministry of Agriculture and Rural Affairs, National Fishery Technical Extension Center, Ed.; China Agriculture Press: Beijing, China, 2018; pp. 187–204. (In Chinese) [Google Scholar]
- Ramos, P. Parasites in fishery products—Laboratorial and educational strategies to control. Exp. Parasitol. 2020, 211, 107865. [Google Scholar] [CrossRef]
- Department of Agriculture, Fisheries and Forestry. 2022. Available online: https://www.outbreak.gov.au/current-responses-to-outbreaks/white-spot-disease (accessed on 1 March 2023).
- Department of Agriculture, Fisheries and Forestry. Review of the Biosecurity Risks of Prawns Imported from All Countries for Human Consumption-Final Report; Canberra, Australia. 2023. Available online: agriculture.gov.au (accessed on 1 August 2023).
- The Fish Site. Vietnam on Alrert for Decapod Iridescent Virus. 2020. Available online: https://thefishsite.com/articles/vietnam-on-alert-for-decapod-iridescent-virus (accessed on 1 March 2023).
- Ko, J.-Y.; Seul, D.-E.; Kim, S.-H.; Magollah, T.M.; Kwak, M.-G.; Song, T.-W.; Lee, Y.-B. Manufacture optimization and quality evaluation of artificial fishing lures. In Proceedings of the Academic Conference of Aquaculture Division of Korean Society of Fisheries Science, Online, 4–6 November 2020; p. 274. Available online: https://www.dbpia.co.kr/pdf/pdfView.do?nodeId=NODE09380505 (accessed on 1 March 2023). (In Korean).
- KOSIS. Available online: https://kosis.kr/index/index.do (accessed on 1 July 2023).
- Sritunyalucksana, K.; Apisawetakan, S.; Boon-Nat, A.; Withyachumnarnkul, B.; Flegel, T.W. A new RNA virus found in black tiger shrimp Penaeus monodon from Thailand. Virus Res. 2006, 118, 31–38. [Google Scholar] [CrossRef]
- Prakasha, B.K.; Ramakrishna, R.P.; Karunasagar, I.; Karunasagar, I. Detection of Laem-Singh virus (LSNV) in cultured Penaeus monodon from India. Dis. Aquat. Org. 2007, 77, 83–86. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Q.; Liu, S.; Yang, H.; Liu, S.; Zhu, L.; Yang, B.; Jin, J.; Ding, L.; Wang, X.; et al. A new nodavirus is associated with covert mortality disease of shrimp. J. Gen. Virol. 2014, 95, 2700–2709. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, M.-M.; Wan, X.-Y.; Li, C.; Zhang, Q.-L.; Wang, R.-Y.; Cheng, D.-Y.; Dong, X.; Yang, B.; Wang, X.-H.; et al. Characterization of a new member of Iridoviridae, shrimp hemocyte iridescent virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci. Rep. 2017, 7, 11834. [Google Scholar] [CrossRef]
- Park, S.C.; Choi, S.-K.; Han, S.-H.; Park, S.; Jeon, H.J.; Lee, S.C.; Kim, K.Y.; Lee, Y.S.; Kim, J.H.; Han, J.E. Detection of infectious hypodermal and hematopoietic necrosis virus and white spot syndrome virus in whiteleg shrimp (Penaeus vannamei) imported from Vietnam to South Korea. J. Vet. Sci. 2020, 21, e31. [Google Scholar] [CrossRef]
- Sarri, C.; Stamatis, C.; Sarafidou, T.; Galara, I.; Godosopoulos, V.; Kolovos, M.; Liakou, C.; Tastsoglou, S.; Mamuris, Z. A new set of 16S rRNA universal primers for identification of animal species. Food Control 2014, 43, 35–41. [Google Scholar] [CrossRef]
- WOAH. Infection with White Spot Syndrome Virus (Version Adopted in 2018). In Manual of Diagnostic Tests for Aquatic Animals 2022; World Organisation for Animal Health: Paris, France, 2022; Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/aahm/current/2.2.08_WSSV.pdf (accessed on 15 February 2023).
- Trang, T.T.; Hung, N.H.; Ninh, N.H.; Nguyen, N.H. Selection for improved white spot syndrome virus resistance increased larval survival and growth rate of Pacific whiteleg shrimp, Liptopenaeus vannamei. J. Invertebr. Pathol. 2019, 166, 107219. [Google Scholar] [CrossRef]
- Cruz-Flores, R.; Mai, H.N.; Kanrar, S.; Aranguren-Caro, L.F.; Dhar, A.K. Genome reconstruction of white spot syndrome virus (WSSV) from archival Davidson’s-fixed paraffin embedded shrimp (Penaeus vannamei) tissue. Sci. Rep. 2020, 10, 13425. [Google Scholar] [CrossRef] [PubMed]
- Janewanthanakul, S.; Supungul, P.; Tang, S.; Tassanakajon, A. Heat shock protein 70 from Litopenaeus vannamei (LvHSP70) is involved in the innate immune response against white spot syndrome virus (WSSV) infection. Dev. Comp. Immunol. 2020, 102, 103476. [Google Scholar] [CrossRef] [PubMed]
- Hasson, K.W.; Fan, Y.; Reisinger, T.; Venuti, J.; Varner, P.W. White-spot syndrome virus (WSSV) introduction into the Gulf of Mexico and Texas freshwater systems through imported, frozen bait-shrimp. Dis. Aquat. Org. 2006, 71, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.; Nunan, L.; Redman, R.M.; Lightner, D.V.; Fitzsimmons, K. EMS/AHPNS: Infectious disease caused by bacteria. Glob. Aquac. Advocate 2013, 20, 18–20. [Google Scholar]
- Lightner, D.V.; Mohney, L.L.; Williams, R.R.; Redman, R.M. Glycerol tolerance of IHHN virus of penaeid shrimp. J. World Aquac. Soc. 1987, 18, 196–197. [Google Scholar] [CrossRef]
- Lightner, D.V.; Redman, R.M.; Arce, S.; Moss, S.M. Specific pathogen-free (SPF) shrimp stocks in shrimp farming facilities as a novel method for disease control in crustaceans. In Shellfish Safety and Quality; Shumway, S., Rodrick, G., Eds.; Woodhead Publishers: London, UK, 2009; pp. 384–424. [Google Scholar]
- Lightner, D.V. Epizootiology, distribution and the impact on international trade of two penaeid shrimp viruses in the Americas. Rev. Sci. Tech. L’office Int. Epizoot. 1996, 15, 579–601. [Google Scholar] [CrossRef]
- Lightner, D.V.; Redman, R.M.; Poulos, B.T.; Nunan, L.M.; Mari, J.L.; Hasson, K.W. Risk of spread of penaeid shrimp viruses in the Americas by the international movement of live and frozen shrimp. Rev. Sci. Tech. L’office Int. Epizoot. 1997, 16, 146–160. [Google Scholar] [CrossRef]
- Wang, C.S.; Tang, K.F.J.; Kou, G.H.; Chen, S.N. Light and electron microscopic evidence of white spot disease in the giant tiger shrimp, Penaeus monodon (Fabricius), and the kuruma shrimp, Penaeus japonicus (Bate), cultured in Taiwan. J. Fish Dis. 1997, 20, 323–331. [Google Scholar] [CrossRef]
- Wang, Q.; White, B.L.; Redman, R.M.; Lightner, D.V. Per os challenge of Litopenaeus vannamei postlarvae and Farfantepenaeus duorarum juveniles with six geographic isolates of white spot syndrome virus. Aquaculture 1999, 170, 179–194. [Google Scholar] [CrossRef]
- Nunan, L.M.; Poulos, B.T.; Lightner, D.V. The detection of white spot syndrome virus (WSSV) and yellow head virus (YHV) in imported commodity shrimp. Aquaculture 1998, 160, 19–30. [Google Scholar] [CrossRef]
- John, K.R.; George, M.R.; Iyappan, T.; Thangarani, A.J.; Jeyaseelan, M.J.P. Indian isolates of white spot syndrome virus exhibit variations in their pathogenicity and genomic tandem repeats. J. Fish Dis. 2010, 33, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.A.; Jeyasekaran, G.; Jeya, S.R. Incidence of white spot syndrome virus (WSSV) in Indian farmed frozen shrimp products and testing for viability through bio-inoculation studies. J. Aquac. Res. Dev. 2010, 1, 102. [Google Scholar]
- Aranguren, L.F.C.; Mai, H.N.; Nunan, L.; Lin, J.; Noble, B.; Dhar, A.K. Assessment of transmission risk in WSSV-infected shrimp Litopenaeus vannamei upon cooking. J. Fish Dis. 2020. epub ahead of print. [Google Scholar] [CrossRef]
- Otta, S.K.; Arulraj, R.; Ezhil Praveena, P.; Manivel, R.; Panigrahi, A.; Bhuvaneswari, T.; Ravichandran, P.; Jithendran, K.P.; Ponniah, A.G. Association of dual viral infection with mortality of Pacific white shrimp (Litopenaeus vannamei) in culture ponds in India. Virusdisease 2014, 25, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Kotob, M.H.; Menanteau-Ledouble, S.; Kumar, G.; Abdelzaher, M.; El-Matbouli, M. The impact of co-infections on fish: A review. Vet. Res. 2017, 47, 98. [Google Scholar] [CrossRef] [PubMed]
- Kotob, M.H.; Gorgoglione, B.; Kumar, G.; Abdelzaher, M.; Saleh, M.; El-Matbouli, M. The impact of Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections on pathology in rainbow trout. Parasites Vectors 2017, 10, 442. [Google Scholar] [CrossRef]
- Tang, K.F.; Durand, S.V.; White, B.L.; Redman, R.M.; Mohney, L.L.; Lightner, D.V. Induced resistance to white spot syndrome virus infection in Penaeus stylirostris through pre-infection with infectious hypodermal and hematopoietic necrosis virus—A preliminary study. Aquaculture 2003, 216, 19–29. [Google Scholar] [CrossRef]
- Kotob, M.H.; Kumar, G.; Saleh, M.; Gorgoglione, B.; Abdelzaher, M.; El-Matbouli, M. Differential modulation of host immune genes in the kidney and cranium of the rainbow trout (Oncorhynchus mykiss) in response to Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections. Parasites Vectors 2018, 11, 326. [Google Scholar] [CrossRef]
- Rai, P.; Pradeep, B.; Karunasagar, I.; Karunasagar, I. Detection of viruses in Penaeus monodon from India showing signs of slow growth syndrome. Aquaculture 2009, 289, 231–235. [Google Scholar] [CrossRef]
- Lotz, J.M. Special topic review: Viruses, biosecurity and specific pathogen-free stocks in shrimp aquaculture. World J. Microbiol. Biotechnol. 1997, 13, 405–413. [Google Scholar] [CrossRef]
- Jaroenlak, P.; Sanguanrut, P.; Williams, B.A.P.; Stentiford, G.D.; Flegel, T.W.; Sritunyalucksana, K.; Itsathitphaisarn, O. A nested PCR assay to avoid false positive detection of the microsporidian Enterocytozoon hepatopenaei (EHP) in environmental samples in shrimp farms. PLoS ONE 2016, 11, e0166320. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.F.; Redman, R.M.; Pantoja, C.R.; Groumellec, M.L.; Duraisamy, P.; Lightner, D.V. Identification of an iridovirus in Acetes erythraeus (Sergestidae) and the development of in situ hybridization and PCR method for its detection. J. Invertebr. Pathol. 2007, 96, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Poulos, B.T.; Lightner, D.V. Detection of infectious myonecrosis virus (IMNV) of penaeid shrimp by reverse-transcriptase polymerase chain reaction (RT-PCR). Dis. Aquat. Org. 2006, 73, 69–72. [Google Scholar] [CrossRef]
- Aranguren, L.F.; Tang, K.F.J.; Lightner, D.V. Quantification of the bacterial agent of necrotizing hepatopancreatitis (NHP-B) by real-time PCR and comparison of survival and NHP load of two shrimp populations. Aquaculture 2010, 307, 187–192. [Google Scholar] [CrossRef]
- Dangtip, S.; Sirikharin, R.; Sanguanrut, P.; Thitamadee, S.; Sritunyalucksana, K.; Taengchaiyaphum, S.; Mavichak, R.; Proespraiwong, P.; Flegel, T.W. AP4 method for two-tube nested PCR detection of AHPND isolates of Vibrio parahaemolyticus. Aquac. Rep. 2015, 2, 158–162. [Google Scholar] [CrossRef]
- Lo, C.F.; Ho, C.H.; Peng, S.E.; Chen, C.H.; Hsu, H.C.; Chiu, Y.L.; Chang, C.F.; Liu, K.F.; Su, M.S.; Wang, C.H.; et al. White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Dis. Aquat. Org. 1996, 27, 215–225. [Google Scholar] [CrossRef]
- Yoganandhan, K.; Sri Widada, J.; Bonami, J.R.; Sahul Hameed, A.S. Simultaneous detection of Macrobrachium rosenbergii nodavirus and extra small virus by a single tube, one-step multiplex RT-PCR assay. J. Fish Dis. 2005, 28, 65–69. [Google Scholar] [CrossRef]
- WOAH. Infection with yellow head virus genotype 1 (Version Adopted in 2019). In Manual of Diagnostic Tests for Aquatic Animals 2022; World Organisation for Animal Health: Paris, France, 2022; Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/aahm/current/2.2.09_YHD.pdf (accessed on 1 October 2022).
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef]
- Lightner, D.V.; Redman, R.M.; Pantoja, C.; Noble, B.L.; Tran, L. Early mortality syndrome affects shrimp in Asia. Glob. Aquac. Advocate 2012, 40. Available online: https://www.globalseafood.org/advocate/early-mortality-syndrome-affects-shrimp-in-asia/ (accessed on 1 March 2022).
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Org. 2013, 105, 45–55. [Google Scholar] [CrossRef]
- Kondo, H.; Van, P.T.; Dang, L.T.; Hirono, I. Draft genome sequence of non-Vibrio parahaemolyticus acute hepatopancreatic necrosis disease strain KC13.17.5, isolated from diseased shrimp in Vietnam. Genome Announc. 2015, 3, e00978-15. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.T.; Chen, I.T.; Yang, Y.T.; Ko, T.P.; Huang, Y.T.; Huang, J.Y.; Huang, M.F.; Lin, S.J.; Chen, C.Y.; Lin, S.S.; et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc. Natl. Acad. Sci. USA 2015, 112, 10798–10803. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wang, H.; Zou, P.; Chen, J.; Liu, Z.; Wang, X.; Huang, J. Complete genome sequence of Vibrio campbellii strain 20130629003S01 isolated from shrimp with acute hepatopancreatic necrosis disease. Gut Pathog. 2017, 9, 31. [Google Scholar] [CrossRef]
- Eshik, M.E.; Abedin, M.; Punom, N.; Begum, M.K.; Rahman, M.S. Molecular Identification of AHPND positive Vibrio parahaemolyticus causing an outbreak in South-West shrimp farming regions of Bangladesh. J. Bangladesh Acad. Sci. 2018, 41, 127–135. [Google Scholar] [CrossRef]
- Megahed, M.E. Quantitative genetics of disease resistance in different groups of Penaeus semisulcatus from preliminary controlled challenge test with Vibrio parahaemolyticus the aetiological agent of acute hepatopancreatic necrosis disease (AHPND). J. Appl. Aquac. 2018, 1–17, epub ahead of print. [Google Scholar]
- Nunan, L.M.; Pantoja, C.R.; Gomez-Jimenez, S.; Lightner, D.V. “Candidatus Hepatobacter penaei”, an intracellular pathogenic enteric bacterium in the hepatopancreas of the marine shrimp Penaeus vannamei (Crustacea: Decapoda). Appl. Environ. Microbiol. 2013, 79, 1407–1409. [Google Scholar] [CrossRef]
- Hudson, D.A.; Hudson, N.B.; Pyecroft, S.B. Mortalities of Penaeus japonicus prawns associated with microsporidean infection. Aust. Vet. J. 2001, 79, 504–505. [Google Scholar] [CrossRef]
- Tourtip, S.; Wongtripop, S.; Stentiford, G.D.; Bateman, K.S.; Sriurairatana, S.; Chavadej, J.; Sritunyalucksana, K.; Withyachumnarnkul, B. Enterocytozoon hepatopenaei sp. nov. (Microsporida: Enterocytozoonidae), a parasite of the black tiger shrimp Penaeus monodon (Decapoda: Penaeidae): Fine structure and phylogenetic relationships. J. Invertebr. Pathol. 2009, 102, 21–29. [Google Scholar] [CrossRef]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Aranguren, L.F.; Han, J.E.; Tang, K.F.J. Enterocytozoon hepatopenaei (EHP) is a risk factor for acute hepatopancreatic necrosis disease (AHPND) and septic hepatopancreatic necrosis (SHPN) in the Pacific white shrimp Penaeus vannamei. Aquaculture 2017, 471, 37–42. [Google Scholar] [CrossRef]
- Salachan, P.V.; Jaroenlak, P.; Thitamadee, S.; Itsathitphaisarn, O.; Sritunyalucksana, K. Laboratory cohabitation challenge model for shrimp hepatopancreatic microsporidiosis (HPM) caused by Enterocytozoon hepatopenaei (EHP). BMC Vet. Res. 2017, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.F.J.; Aranguren, L.F.; Piamsomboon, P.; Han, J.E.; Maskaykina, I.Y.; Schmidt, M.M. Detection of the microsporidian Enterocytozoon hepatopenaei (EHP) and Taura syndrome virus in Penaeus vannamei cultured in Venezuela. Aquaculture 2017, 480, 17–21. [Google Scholar] [CrossRef]
- Ma, B.; Yu, H.T.; Fang, J.B.; Sun, C.; Zhang, M. Employing DNA binding dye to improve detection of Enterocytozoon hepatopenaei in real-time LAMP. Sci. Rep. 2019, 9, 15860. [Google Scholar] [CrossRef]
- Kim, B.S.; Jang, G.I.; Kim, S.M.; Kim, Y.S.; Jeon, Y.G.; Oh, Y.K.; Hwang, J.Y.; Kwon, M.G. First report of Enterocytozoon hepatopenaei infection in Pacific whiteleg shrimp (Litopenaeus vannamei) cultured in Korea. Animals 2021, 11, 3150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xu, T.; Wan, X.; Liu, S.; Wang, X.; Li, X.; Dong, X.; Yang, B.; Huang, J. Prevalence and distribution of covert mortality nodavirus (CMNV) in cultured crustacean. Virus Res. 2017, 233, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Liu, S.; Li, J.; Xu, T.T.; Wang, X.H.; Fu, G.M.; Li, X.P.; Sang, S.W.; Bian, X.D.; Hao, J.W. Evidence for cross-species transmission of covert mortality nodavirus to new host of Mugilogobius abei. Front. Microbiol. 2018, 9, 1447. Available online: https://www.frontiersin.org/article/10.3389/fmicb.2018.01447 (accessed on 1 March 2022). [CrossRef] [PubMed]
- Wang, C.; Liu, S.; Li, X.; Hao, J.; Tang, K.F.J.; Zhang, Q. Infection of covert mortality nodavirus in Japanese flounder reveals host jump of the emerging alphanodavirus. J. Gen. Virol. 2018, 100, 166–175. [Google Scholar] [CrossRef]
- Xu, L.; Wang, T.; Li, F.; Yang, F. Isolation and preliminary characterization of a new pathogenic iridovirus from redclaw crayfish Cherax quadricarinatus. Dis. Aquat. Org. 2016, 120, 17–26. [Google Scholar] [CrossRef]
- Li, F.; Xu, L.; Yang, F. Genomic characterization of a novel iridovirus from redclaw crayfish Cherax quadricarinatus: Evidence for a new genus within the family Iridoviridae. J. Gen. Virol. 2017, 98, 2589–2595. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, M.-M.; Wan, X.-Y.; Zhang, Q.-L.; Li, C.; Dong, X.; Yang, B.; Huang, J. Detection and quantification of shrimp hemocyte iridescent virus by TaqMan probe based real-time PCR. J. Invertebr. Pathol. 2018, 154, 95–1001. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, M.M.; Wang, R.Y.; Wan, X.Y.; Li, C.; Zhang, Q.L.; Dong, X.; Yang, B.; Xiang, J.H.; Huang, J. Complete genome sequence of shrimp hemocyte iridescent virus (SHIV) isolated from white leg shrimp, Litopenaeus vannamei. Arch. Virol. 2018, 163, 781–785. [Google Scholar] [CrossRef]
- Qiu, L.; Chen, X.; Zhao, R.; Li, C.; Gao, W.; Zhang, Q.; Huang, J. First description of a natural infection with shrimp hemocyte iridescent virus in farmed giant freshwater prawn, Macrobrachium rosenbergii. Viruses 2019, 11, 354. [Google Scholar] [CrossRef]
- Ramsden, N.; Smith, J. Clarification: Shrimp Disease SHIV Detected in China, Thailand, but Not Vietnam. Undercurrent News, 2018. Available online: https://www.undercurrentnews.com/2018/10/01/clarification-shrimp-disease-shiv-detected-in-chinathailand-but-not-vietnam/(accessed on 1 February 2023).
- Chen, X.; Qiu, L.; Wang, H.; Zou, P.; Dong, X.; Li, F.; Huang, J. Susceptibility of Exopalaemon carinicauda to the infection with shrimp hemocyte iridescent virus. Viruses 2019, 11, 387. [Google Scholar] [CrossRef] [PubMed]
- Chung, J. Sixteen shrimp farms infected with DIV1: Agriculture council. In Taipei Times; Taipei Times: Taipei, Taiwan, 2020; Available online: https://www.taipeitimes.com/News/taiwan/archives/2020/06/19/2003738488 (accessed on 1 February 2023).
- Lightner, D.V.; Redman, R.M. Shrimp diseases and current diagnostic methods. Aquaculture 1998, 164, 201–220. [Google Scholar] [CrossRef]
- Rai, P.; Safeena, M.P.; Krabsetsve, K.; La Fauce, K.; Owens, L.; Karunasagar, I. Genomics, molecular epidemiology and diagnostics of infectious hypodermal and hematopoietic necrosis virus. Indian J. Virol. 2012, 23, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Bateman, K.S.; Stentiford, G.D. A taxonomic review of viruses infecting crustaceans with an emphasis on wild hosts. J. Invertebr. Pathol. 2017, 147, 86–110. [Google Scholar] [CrossRef] [PubMed]
- Srisala, J.; Thaiue, D.; Sanguanrut, P.; Aldama-Cano, D.J.; Flegel, T.W.; Sritunyalucksana, K. Potential universal PCR method to detect decapod hepanhamaparvovirus (DHPV) in crustaceans. Aquaculture 2021, 541, 736782. [Google Scholar] [CrossRef]
- Tang, K.F.J.; Pantoja, C.R.; Lightner, D.V. Infectious myonecrosis virus infection in Litopenaeus vannamei, Litopenaeus stylirostris, and Penaeus monodon. In Aquaculture America 2005: Shrimp Growout, Nutrition and Disease; World Aquaculture Society: New Orleans, LA, USA, 2005; pp. 1–2. [Google Scholar]
- Coelho, M.G.L.; Silva, A.C.G.; Vila Nova, C.M.V.; Neto, J.M.O.; Lima, A.C.N.; Feijó, R.G.; Apolinário, D.F.; Maggioni, R.; Gesteira, T.C.V. Susceptibility of the wild southern brown shrimp (Farfantepenaeus subtilis) to infectious hypodermal and hematopoietic necrosis (IHHN) and infectious myonecrosis (IMN). Aquaculture 2009, 294, 1–4. [Google Scholar] [CrossRef]
- Sahul Hameed, A.S.; Abdul Majeed, S.; Vimal, S.; Madan, N.; Rajkumar, T.; Santhoshkumar, S.; Sivakumar, S. Studies on the occurrence of infectious myonecrosis virus in pond-reared Litopenaeus vannamei (Boone, 1931) in India. J. Fish Dis. 2017, 40, 1823–1830. [Google Scholar] [CrossRef]
- Sittidilokratna, N.; Dangtip, S.; Sritunyalucksana, K.; Babu, R.; Pradeep, B.; Mohan, C.V.; Gudkovs, N.; Walker, P.J. Detection of Laem-Singh virus in cultured Penaeus monodon shrimp from several sites in the IndoPacific region. Dis. Aquat. Org. 2009, 84, 195–200. [Google Scholar] [CrossRef]
- Panphut, W.; Senapin, S.; Sriurairatana, S.; Withyachumnarnkul, B.; Flegel, T.W. A novel integrase-containing element may interact with laem-singh virus (LSNV) to cause slow growth in giant tiger shrimp. BMC Vet. Res. 2011, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Poornima, M.; Seetang-Nun, Y.; Alavandi, S.V.; Dayal, J.S. Laem-Singh virus: A probable etiological agent associated with monodon slow growth syndrome in farmed black tiger shrimp (Penaeus monodon). Indian J. Virol. 2012, 23, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, X.; Tian, J.; Chen, L.; Chen, X.; Li, C.; Qin, X.; Li, J.; Cao, J.; Eden, J.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539. [Google Scholar] [CrossRef] [PubMed]
- Taengchaiyaphum, S.; Srisala, J.; Sanguanrut, P.; Ongvarrasopone, C.; Flegel, T.W.; Sritunyalucksana, K. Full genome characterization of Laem Singh virus (LSNV) in shrimp Penaeus monodon. Aquaculture 2021, 538, 736533. [Google Scholar] [CrossRef]
- Brock, J.A. Taura syndrome, a disease important to shrimp farms in the Americas. World J. Microbiol. Biotechnol. 1997, 13, 415–418. [Google Scholar] [CrossRef]
- Jimenez, R.; Barniol, R.; de Barniol, L.; Machuca, M.; Romero, X. Viral-like particles associated with cuticular epithelium necrosis in cultured Litopenaeus vannamei (Decapoda: Crustacea) in Ecuador. Aquac. Res. 2000, 31, 519–528. [Google Scholar] [CrossRef]
- Inouye, K.; Miwa, S.; Oseko, N.; Nakano, H.; Kimura, T.; Momoyama, K.; Hiraoka, M. Mass mortalities of cultured kuruma shrimp Penaeus japonicus in Japan in 1993: Electron microscopic evidence of the causative virus. Fish Pathol. 1994, 29, 149–158. (In Japanese) [Google Scholar] [CrossRef]
- Takahashi, Y.; Itami, T.; Kondo, M.; Maeda, M.; Fuji, R.; Tomonaga, S.; Supamattaya, K.; Boonyaratpalin, S. Electron microscopic evidence of bacilliform virus infection in kuruma shrimp (Penaeus japonicus). Fish Pathol. 1994, 29, 121–125. [Google Scholar] [CrossRef]
- Wongteerasupaya, C.; Wongwisansri, S.; Boonsaeng, V.; Panyim, S.; Pratanpipat, P.; Nash, G.L.; Withyachumnarnkul, B.; Flegel, T.W. DNA fragment of Penaeus monodon baculovirus PmNOBII gives positive in situ hybridization with white-spot viral infections in six penaeid shrimp species. Aquaculture 1996, 143, 23–32. [Google Scholar] [CrossRef]
- Lightner, D.V.; Hasson, K.W.; White, B.L.; Redman, R.M. Experimental infection of Western hemisphere penaeid shrimp with Asian white spot syndrome virus and Asian yellow head virus. J. Aquat. Anim. Health 1998, 10, 271–281. [Google Scholar] [CrossRef]
- van Hulten, M.C.W.; Witteveldt, J.; Peters, S.; Kloosterboer, N.; Tarchini, R.; Fiers, M.; Sandbrink, H.; Lankhorst, R.; Vlak, J.M. The white spot syndrome virus DNA genome sequence. Virology 2001, 286, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; He, J.; Lin, X.; Li, Q.; Pan, D.; Zhang, X.; Xu, X. Complete genome sequence of the shrimp white spot bacilliform virus. J. Virol. 2001, 75, 11811–11820. [Google Scholar] [CrossRef]
- Sudhakaran, R.; Syed Musthaq, S.; Haribabu, P.; Mukherjee, S.C.; Gopal, C.; Sahul Hameed, A.S. Experimental transmission of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) in three species of marine shrimp (Penaeus indicus, Penaeus japonicus and Penaeus monodon). Aquaculture 2006, 257, 136–141. [Google Scholar] [CrossRef]
- Ravi, M.; Nazeer Basha, A.; Sarathi, M.; Rosa Idalia, H.H.; Sri Widada, J.; Bonami, J.R.; Sahul Hameed, A.S. Studies on the occurrence of white tail disease (WTD) caused by MrNV and XSV in hatchery-reared postlarvae of Penaeus indicus and P. monodon. Aquaculture 2009, 292, 117–120. [Google Scholar] [CrossRef]
- Pillai, D.; Bonami, J.R. A review on the diseases of freshwater prawns with special focus on white tail disease of Macrobrachium rosenbergii. Aquac. Res. 2012, 43, 1029–1037. [Google Scholar] [CrossRef]
- Senapin, S.; Jaengsanong, C.; Phiwsaiya, K.; Prasertsri, S.; Laisutisan, K.; Chuchird, N.; Limsuwan, C.; Flegel, T.W. Infections of MrNV (Macrobrachium rosenbergii nodavirus) in cultivated whiteleg shrimp Penaeus vannamei in Asia. Aquaculture 2012, 338–341, 41–46. [Google Scholar] [CrossRef]
- Kibenge, F.S.B.; Godoy, M.G. Aquaculture Virology, 1st ed.; Elsevier Inc.: London, UK, 2016. [Google Scholar]
- Murwantoko, M.; Bimantara, A.; Roosmanto, R.; Kawaichi, M. Macrobrachium rosenbergii nodavirus infection in a giant freshwater prawn hatchery in Indonesia. SpringerPlus 2016, 5, 1729. [Google Scholar] [CrossRef]
- Gangnonngiw, W.; Bunnontae, M.; Phiwsaiya, K.; Senapin, S.; Dhar, A.K. In experimental challenge with infectious clones of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV), MrNV alone can cause mortality in freshwater prawn (Macrobrachium rosenbergii). Virology 2020, 540, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Chayaburakul, K.; Nash, G.; Pratanpipat, P.; Sriurairatana, S.; Withyachumnarnkul, B. Multiple pathogens found in growth-retarded black tiger shrimp Penaeus monodon cultivated in Thailand. Dis. Aquat. Org. 2004, 60, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Flegel, T.W.; Nielsen, L.; Thamavit, V.; Kongtim, S.; Pasharawipas, T. Presence of multiple viruses in non-diseased, cultivated shrimp at harvest. Aquaculture 2004, 240, 55–68. [Google Scholar] [CrossRef]
- Megahed, M.E.; Cruz-Flores, R.; Dhar, A.K. Complete genome sequences of four major viruses infecting marine shrimp in Egypt. Microbiol. Resour. Announc. 2018, 7, e00809-18. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| No. | Samples | Sampling Locations | Origins |
|---|---|---|---|
| 1 | Metapenaeus joyneri, Portunus trituberculatus, Uroteuthis duvaucelii | Jeonnam | China |
| 2 | Eriocheir sinensis | Jeonnam | China |
| 3 | Perinereis aibuhitensis, Marphysa sanguinea, Metapenaeus joyneri, Euphausia superba | Jeonnam | China, Antarctic Ocean |
| 4 | Perinereis aibuhitensis, Loliolus beka, Todarodes pacificus, Amphioctopus fangsiao, Palaemon orientis, Euphausia superba, Metapenaeus joyneri, Penaeus vannamei | Chungnam | Vietnam, Antarctic Ocean |
| 5 | Loliolus beka | Jeonnam | China |
| 6 | Artemia franciscana | Internet Market | China |
| 7 | Perinereis aibuhitensis, Atrina pectinata | Jeonbuk | China |
| 8 | Artemia franciscana | Chungnam | United States |
| 9 | Todarodes pacificus | Gyeongnam | China |
| 10 | Penaeus vannamei, Euphausia superba, Perinereis aibuhitensis | Jeju | China, Antarctic Ocean |
| 11 | Marphysa sanguinea, Penaeus vannamei, Metapenaeus joyneri | Gyeonggi | China |
| 12 | Eriocheir sinensis | Jeonbuk | China |
| 13 | Perinereis aibuhitensis, Marphysa sanguinea | Jeonnam | China |
| 14 | Perinereis aibuhitensis, Marphysa sanguinea, Dosidicus gigas, Euphausia superba | Kangwon | China, Argentina, Antarctic Ocean |
| 15 | Metapenaeus joyneri, Perinereis aibuhitensis | Gyeongbuk | China |
| Phylum | Species and Quantity |
|---|---|
| Crustacean | 11 of Portunus trituberculatus |
| 159 of Metapenaeus joyneri | |
| 60 of Eriocheir sinensis | |
| 210 of Euphausia superba | |
| 30 of Palaemon orientis | |
| 104 of Penaeus vannamei | |
| 24 pooled samples * of Artemia franciscana | |
| Semi-total | 574 samples of 7 species + 24 pooled samples * |
| Mollusks | 11 of Uroteuthis duvaucelii |
| 20 of Loliolus beka | |
| 199 of Perinereis aibuhitensis | |
| 57 of Marphysa sanguinea | |
| 68 of Todarodes pacificus | |
| 10 of Amphioctopus fangsiao | |
| 1 of Atrina pectinata | |
| 18 of Dosidicus gigas | |
| Semi-total | 384 animals of 8 species |
| Total | 1556 samples of 37 species (+ 24 pooled samples *) |
| No. | Detailed Region | Species | Origin | Prevalence |
|---|---|---|---|---|
| 1 | Jeonnam Mokpo | M. joyneri | China | 20% (6/30) of WSSV |
| 2 | Jeonnam Jindo | P. vannamei | Vietnam | 100% (24/24) of WSSV |
| 3 | Jeju Hamduk | P. vannamei | Vietnam | 100% (25/25) of WSSV and 28% (7/25) of VpAHPND |
| 4 | Incheon Ganghwa | M. joyneri | Vietnam | 26.7% (8/30) of WSSV |
| 5 | Incheon Ganghwa | P. vannamei | Vietnam | 100% (25/25) of WSSV |
| 6 | Gyeongbuk Ulgin | M. joyneri | China | 30% (9/30) of WSSV |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, G.; Woo, W.-S.; Kim, K.-H.; Son, H.-J.; Sohn, M.-Y.; Kong, H.J.; Kim, Y.-O.; Kim, D.-G.; Kim, E.M.; Noh, E.S.; et al. Identification of Potential Hazards Associated with South Korean Prawns and Monitoring Results Targeting Fishing Bait. Pathogens 2023, 12, 1228. https://doi.org/10.3390/pathogens12101228
Kang G, Woo W-S, Kim K-H, Son H-J, Sohn M-Y, Kong HJ, Kim Y-O, Kim D-G, Kim EM, Noh ES, et al. Identification of Potential Hazards Associated with South Korean Prawns and Monitoring Results Targeting Fishing Bait. Pathogens. 2023; 12(10):1228. https://doi.org/10.3390/pathogens12101228
Chicago/Turabian StyleKang, Gyoungsik, Won-Sik Woo, Kyung-Ho Kim, Ha-Jeong Son, Min-Young Sohn, Hee Jeong Kong, Young-Ok Kim, Dong-Gyun Kim, Eun Mi Kim, Eun Soo Noh, and et al. 2023. "Identification of Potential Hazards Associated with South Korean Prawns and Monitoring Results Targeting Fishing Bait" Pathogens 12, no. 10: 1228. https://doi.org/10.3390/pathogens12101228
APA StyleKang, G., Woo, W. -S., Kim, K. -H., Son, H. -J., Sohn, M. -Y., Kong, H. J., Kim, Y. -O., Kim, D. -G., Kim, E. M., Noh, E. S., & Park, C. -I. (2023). Identification of Potential Hazards Associated with South Korean Prawns and Monitoring Results Targeting Fishing Bait. Pathogens, 12(10), 1228. https://doi.org/10.3390/pathogens12101228