
Molecular Epidemiology and Genetic Diversity of Human Respiratory Syncytial Virus in Sicily during Pre- and Post-COVID-19 Surveillance Seasons

 , ,
, ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection
2.2. Laboratory Methods
2.3. Phylogenetic Analysis
2.4. Analysis of Deduced Amino Acid Sequences and Mutations
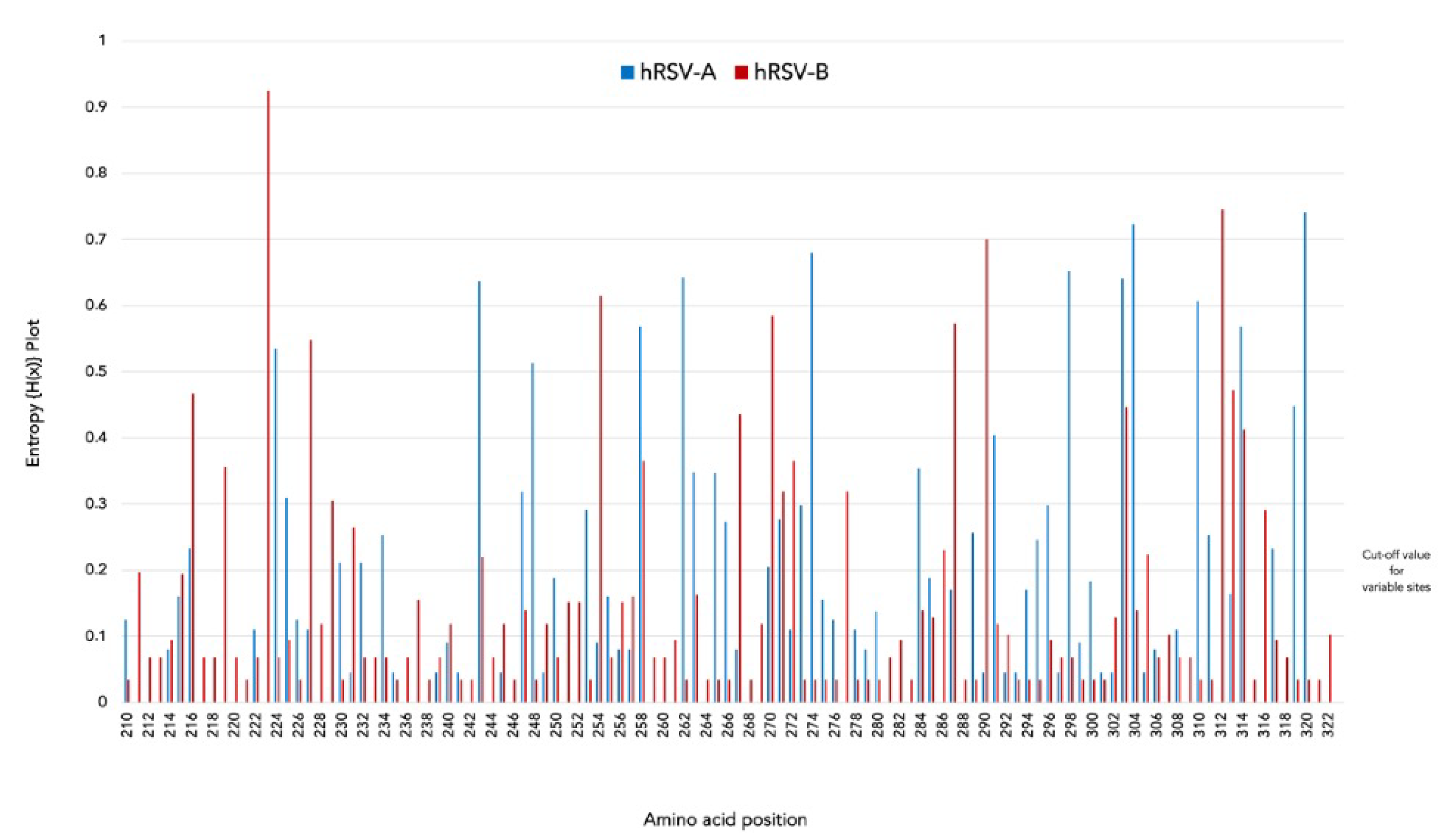
2.5. Entropy Analysis and N-Linked Glycosylation Sites
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Wang, X.; Blau, D.M.; Caballero, M.T.; Feikin, D.R.; Gill, C.J.; Madhi, S.A.; Omer, S.B.; Simões, E.A.F.; Campbell, H.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in children younger than 5 years in 2019: A systematic analysis. Lancet 2022, 399, 2047–2064. [Google Scholar] [CrossRef]
- Savic, M.; Penders, Y.; Shi, T.; Branche, A.; Pirçon, J.Y. Respiratory syncytial virus disease burden in adults aged 60 years and older in high-income countries: A systematic literature review and meta-analysis. Influenza Other Respir. Viruses 2023, 17, e13031. [Google Scholar] [CrossRef]
- Shi, T.; Vennard, S.; Jasiewicz, F.; Brogden, R.; Nair, H.; RESCEU Investigators. Disease Burden Estimates of Respiratory Syncytial Virus related Acute Respiratory Infections in Adults With Comorbidity: A Systematic Review and Meta-Analysis. J. Infect. Dis. 2022, 226, S17–S21. [Google Scholar] [CrossRef]
- Battles, M.B.; McLellan, J.S. Respiratory syncytial virus entry and how to block it. Nat. Rev. Microbiol. 2019, 17, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Pierangeli, A.; Nenna, R.; Fracella, M.; Scagnolari, C.; Oliveto, G.; Sorrentino, L.; Frasca, F.; Conti, M.G.; Petrarca, L.; Papoff, P.; et al. Genetic diversity and its impact on disease severity in respiratory syncytial virus subtype-A and -B bronchiolitis before and after pandemic restrictions in Rome. J. Infect. 2023, S0163-4453(23)00381-X. [Google Scholar] [CrossRef]
- Vila, J.; Lera, E.; Peremiquel-Trillas, P.; Andrés, C.; Martínez, L.; Barceló, I.; Carsi, A.; Balcells, J.; Ángel Rodrigo-Pendás, J.; Soler-Palacín, P.; et al. Increased RSV-A Bronchiolitis Severity in RSV-Infected Children Admitted to a Reference Center in Catalonia (Spain) between 2014 and 2018. J. Pediatr. Infect. Dis Soc. 2023, 12, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Rios Guzman, E.; Hultquist, J.F. Clinical and biological consequences of respiratory syncytial virus genetic diversity. Ther. Adv. Infect. Dis. 2022, 9, 20499361221128091. [Google Scholar] [CrossRef]
- Muñoz-Escalante, J.C.; Comas-García, A.; Bernal-Silva, S.; Noyola, D.E. Respiratory syncytial virus B sequence analysis reveals a novel early genotype. Sci. Rep. 2021, 11, 3452. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qiu, X.; Avadhanula, V.; Shepard, S.S.; Kim, D.K.; Hixson, J.; Piedra, P.A.; Bahl, J. Novel and extendable genotyping system for human respiratory syncytial virus based on whole-genome sequence analysis. Influenza Other Respir. Viruses 2022, 16, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Goya, S.; Galiano, M.; Nauwelaers, I.; Trento, A.; Openshaw, P.J.; Mistchenko, A.S.; Zambon, M.; Viegas, M. Toward unified molecular surveillance of RSV: A proposal for genotype definition. Influenza Other Respir. Viruses 2020, 14, 274–285. [Google Scholar] [CrossRef]
- Muñoz-Escalante, J.C.; Comas-García, A.; Bernal-Silva, S.; Robles-Espinoza, C.D.; Gómez-Leal, G.; Noyola, D.E. Respiratory syncytial virus A genotype classification based on systematic intergenotypic and intragenotypic sequence analysis. Sci. Rep. 2019, 9, 20097. [Google Scholar] [CrossRef]
- Ramaekers, K.; Rector, A.; Cuypers, L.; Lemey, P.; Keyaerts, E.; Van Ranst, M. Towards a unified classification for human respiratory syncytial virus genotypes. Virus. Evol. 2020, 6, veaa052. [Google Scholar] [CrossRef]
- Obando-Pacheco, P.; Justicia-Grande, A.J.; Rivero-Calle, I.; Rodríguez-Tenreiro, C.; Sly, P.; Ramilo, O.; Mejías, A.; Baraldi, E.; Papadopoulos, N.G.; Nair, H.; et al. Respiratory Syncytial Virus Seasonality: A Global Overview. J. Infect. Dis. 2018, 217, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Tramuto, F.; Maida, C.M.; Di Naro, D.; Randazzo, G.; Vitale, F.; Restivo, V.; Costantino, C.; Amodio, E.; Casuccio, A.; Graziano, G.; et al. Respiratory Syncytial Virus: New Challenges for Molecular Epidemiology Surveillance and Vaccination Strategy in Patients with ILI/SARI. Vaccines 2021, 9, 1334. [Google Scholar] [CrossRef] [PubMed]
- Ciofi Degli Atti, M.; Rizzo, C.; D’Amore, C.; Ravà, L.; Reale, A.; Barbieri, M.A.; Bernaschi, P.; Russo, C.; Villani, A.; Perno, C.F.; et al. Acute respiratory infection emergency access in a tertiary care children hospital in Italy, prior and after the SARS-CoV-2 emergence. Influenza Other Respir. Viruses 2023, 17, e13102. [Google Scholar] [CrossRef] [PubMed]
- Nenna, R.; Matera, L.; Pierangeli, A.; Oliveto, G.; Viscido, A.; Petrarca, L.; La Regina, D.P.; Mancino, E.; Di Mattia, G.; Villani, A.; et al. First COVID-19 lockdown resulted in most respiratory viruses disappearing among hospitalised children, with the exception of rhinoviruses. Acta Paediatr. 2022, 111, 1399–1403. [Google Scholar] [CrossRef]
- Garg, I.; Shekhar, R.; Sheikh, A.B.; Pal, S. Impact of COVID-19 on the Changing Patterns of Respiratory Syncytial Virus Infections. Infect. Dis. Rep. 2022, 14, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Redlberger-Fritz, M.; Kundi, M.; Aberle, S.W.; Puchhammer-Stöckl, E. Significant impact of nationwide SARS-CoV-2 lockdown measures on the circulation of other respiratory virus infections in Austria. J. Clin. Virol. 2021, 137, 104795. [Google Scholar] [CrossRef] [PubMed]
- Varela, F.H.; Scotta, M.C.; Polese-Bonatto, M.; Sartor, I.T.S.; Ferreira, C.F.; Fernandes, I.R.; Zavaglia, G.O.; de Almeida, W.A.F.; Arakaki-Sanchez, D.; Pinto, L.A.; et al. Absence of detection of RSV and influenza during the COVID-19 pandemic in a Brazilian cohort: Likely role of lower transmission in the community. J. Glob. Health 2021, 11, 05007. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Xiao, X.; Gu, X.; Liang, D.; Cao, T.; Mou, J.; Huang, C.; Chen, L.; Liu, J. Surveillance of common respiratory infections during the COVID-19 pandemic demonstrates the preventive efficacy of non-pharmaceutical interventions. Int. J. Infect. Dis. 2021, 105, 442–447. [Google Scholar] [CrossRef]
- Den Hartog, G.; van Kasteren, P.B.; Schepp, R.M.; Teirlinck, A.C.; van der Klis, F.R.M.; van Binnendijk, R.S. Decline of RSV-specific antibodies during the COVID-19 pandemic. Lancet Infect. Dis. 2023, 23, 23–25. [Google Scholar] [CrossRef]
- Reicherz, F.; Xu, R.Y.; Abu-Raya, B.; Majdoubi, A.; Michalski, C.; Golding, L.; Stojic, A.; Vineta, M.; Granoski, M.; Cieslak, Z.; et al. Waning immunity against respiratory syncytial virus during the coronavirus disease 2019 pandemic. J. Infect. Dis. 2022, 226, 2064–2068. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Intensified Circulation of Respiratory Syncytial Virus (RSV) and Associated Hospital Burden in the EU/EEA—12 December 2022. ECDC: Stockholm. 2022. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/RRA-20221128-473.pdf (accessed on 27 August 2023).
- Falsey, A.R.; Cameron, A.; Branche, A.R.; Walsh, E.E. Perturbations in Respiratory Syncytial Virus Activity During the SARS-CoV-2 Pandemic. J. Infect. Dis. 2022, 227, 83–86. [Google Scholar] [CrossRef]
- Goya, S.; Sereewit, J.; Pfalmer, D.; Nguyen, T.V.; Bakhash, S.A.K.M.; Sobolik, E.B.; Greninger, A.L. Genomic Characterization of Respiratory Syncytial Virus during 2022-23 Outbreak, Washington, USA. Emerg. Infect. Dis. 2023, 29, 865–868. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Influenza-like Illness definition. In Commission Implementing Decision (EU) 2018/945 of 22 June 2018. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018D0945&from=EN#page=24 (accessed on 27 August 2023).
- Ministero della Salute. Direzione Generale della Prevenzione Sanitaria “Monitoraggio Dell’andamento Delle Forme Gravi e Complicate Di Malattia Da Virus Influenzali e/o da SARS-CoV-2 Confermate in Laboratorio, Stagione 2020–2021”-0042940- 30/12/2020-DGPRE-DGPRE-P. Available online: https://www.trovanorme.salute.gov.it/norme/renderNormsanPdf?anno=2020&codLeg=78081&parte=1%20&serie=null (accessed on 27 August 2023).
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar]
- Agca, H.; Akalin, H.; Saglik, I.; Hacimustafaoglu, M.; Celebi, S.; Ener, B. Changing epidemiology of influenza and other respiratory viruses in the first year of COVID-19 pandemic. J. Infect. Public Health 2021, 14, 1186–1190. [Google Scholar] [CrossRef]
- Hu, W.; Fries, A.C.; DeMarcus, L.S.; Thervil, J.W.; Kwaah, B.; Brown, K.N.; Sjoberg, P.A.; Robbins, A.S. Circulating Trends of Influenza and Other Seasonal Respiratory Viruses among the US Department of Defense Personnel in the United States: Impact of the COVID-19 Pandemic. Int. J. Environ. Res. Public Health 2022, 19, 5942. [Google Scholar] [CrossRef]
- Groves, H.E.; Piché-Renaud, P.P.; Peci, A.; Farrar, D.S.; Buckrell, S.; Bancej, C.; Sevenhuysen, C.; Campigotto, A.; Gubbay, J.B.; Morris, S.K. The impact of the COVID-19 pandemic on influenza, respiratory syncytial virus, and other seasonal respiratory virus circulation in Canada: A population-based study. Lancet Reg. Health Am. 2021, 1, 100015. [Google Scholar] [CrossRef]
- Yeoh, D.K.; Foley, D.A.; Minney-Smith, C.A.; Martin, A.C.; Mace, A.O.; Sikazwe, C.T.; Le, H.; Levy, A.; Blyth, C.C.; Moore, H.C. Impact of Coronavirus Disease 2019 Public Health Measures on Detections of Influenza and Respiratory Syncytial Virus in Children During the 2020 Australian Winter. Clin. Infect. Dis. 2021, 72, 2199–2202. [Google Scholar] [CrossRef]
- Hönemann, M.; Thiem, S.; Bergs, S.; Berthold, T.; Propach, C.; Siekmeyer, M.; Frille, A.; Wallborn, T.; Maier, M.; Pietsch, C. In-Depth Analysis of the Re-Emergence of Respiratory Syncytial Virus at a Tertiary Care Hospital in Germany in the Summer of 2021 after the Alleviation of Non-Pharmaceutical Interventions Due to the SARS-CoV-2 Pandemic. Viruses 2023, 15, 877. [Google Scholar] [CrossRef]
- Esposito, S.; Piralla, A.; Zampiero, A.; Bianchini, S.; Di Pietro, G.; Scala, A.; Pinzani, R.; Fossali, E.; Baldanti, F.; Principi, N. Characteristics and Their Clinical Relevance of Respiratory Syncytial Virus Types and Genotypes Circulating in Northern Italy in Five Consecutive Winter Seasons. PLoS ONE 2015, 10, e0129369. [Google Scholar] [CrossRef] [PubMed]
- Gaymard, A.; Bouscambert-Duchamp, M.; Pichon, M.; Frobert, E.; Vallee, J.; Lina, B.; Casalegno, J.S.; Morfin, F. Genetic characterization of respiratory syncytial virus highlights a new BA genotype and emergence of the ON1 genotype in Lyon, France, between 2010 and 2014. J. Clin. Virol. 2018, 102, 12–18. [Google Scholar] [CrossRef]
- Krivitskaya, V.; Komissarova, K.; Pisareva, M.; Sverlova, M.; Fadeev, A.; Petrova, E.; Timonina, V.; Sominina, A.; Danilenko, D. Respiratory Syncytial Virus G Protein Sequence Variability among Isolates from St. Petersburg, Russia, during the 2013–2014 Epidemic Season. Viruses 2021, 13, 119. [Google Scholar] [CrossRef] [PubMed]
- Sáez-López, E.; Cristóvão, P.; Costa, I.; Pechirra, P.; Conde, P.; Guiomar, R.; Peres, M.J.; Viseu, R.; Lopes, P.; Soares, V.; et al. Epidemiology and genetic variability of respiratory syncytial virus in Portugal, 2014–2018. J. Clin. Virol. 2019, 121, 104200. [Google Scholar] [CrossRef]
- Tabatabai, J.; Ihling, C.M.; Rehbein, R.M.; Schnee, S.V.; Hoos, J.; Pfeil, J.; Grulich-Henn, J.; Schnitzler, P. Molecular epidemiology of respiratory syncytial virus in hospitalised children in Heidelberg, Southern Germany, 2014–2017. Infect. Genet. Evol. 2022, 98, 105209. [Google Scholar] [CrossRef] [PubMed]
- Agoti, C.N.; Otieno, J.R.; Gitahi, C.W.; Cane, P.A.; Nokes, D.J. Rapid spread and diversification of respiratory syncytial virus genotype ON1, Kenya. Emerg. Infect. Dis. 2014, 20, 950–959. [Google Scholar] [CrossRef]
- Razanajatovo Rahombanjanahary, N.H.; Rybkina, K.; Randriambolamanantsoa, T.H.; Razafimanjato, H.; Heraud, J.M. Genetic diversity and molecular epidemiology of respiratory syncytial virus circulated in Antananarivo, Madagascar, from 2011 to 2017: Predominance of ON1 and BA9 genotypes. J. Clin. Virol. 2020, 129, 104506. [Google Scholar] [CrossRef]
- Al-Hassinah, S.; Parveen, S.; Somily, A.M.; AlSaadi, M.M.; Alamery, S.F.; Haq, S.H.; Alsenaidy, H.A.; Ahmed, A. Evolutionary analysis of the ON1 genotype of subtype a respiratory syncytial virus in Riyadh during 2008-16. Infect. Genet. Evol. 2020, 79, 104153. [Google Scholar] [CrossRef]
- Kim, H.N.; Hwang, J.; Yoon, S.Y.; Lim, C.S.; Cho, Y.; Lee, C.K.; Nam, M.H. Molecular characterization of human respiratory syncytial virus in Seoul, South Korea, during 10 consecutive years, 2010–2019. PLoS ONE 2023, 18, e0283873. [Google Scholar] [CrossRef]
- Lee, C.Y.; Fang, Y.P.; Wang, L.C.; Chou, T.Y.; Liu, H.F. Genetic Diversity and Molecular Epidemiology of Circulating Respiratory Syncytial Virus in Central Taiwan, 2008–2017. Viruses 2021, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.P.; Lei, S.Y.; Wang, Y.B.; Wang, Y.Z.; Qiang, H.S.; Yin, Y.F.; Jiang, Z.M.; Zhu, M.; Chen, X.L.; Ye, H.M.; et al. Molecular Evolution of Attachment Glycoprotein (G) and Fusion Protein (F) Genes of Respiratory Syncytial Virus ON1 and BA9 Strains in Xiamen, China. Microbiol. Spectr. 2022, 10, e0208321. [Google Scholar] [CrossRef]
- Yun, K.W.; Choi, E.H.; Lee, H.J. Molecular epidemiology of respiratory syncytial virus for 28 consecutive seasons (1990–2018) and genetic variability of the duplication region in the G gene of genotypes ON1 and BA in South Korea. Arch. Virol. 2020, 165, 1069–1077. [Google Scholar] [CrossRef]
- Guo, Y.J.; Wang, B.H.; Li, L.; Li, Y.L.; Chu, X.L.; Li, W. Epidemiological and genetic characteristics of respiratory syncytial virus infection in children from Hangzhou after the peak of COVID-19. J. Clin. Virol. 2023, 158, 105354. [Google Scholar] [CrossRef]
- Haider, M.S.H.; Khan, W.H.; Deeba, F.; Ali, S.; Ahmed, A.; Naqvi, I.H.; Dohare, R.; Alsenaidy, H.A.; Alsenaidy, A.M.; Broor, S.; et al. BA9 lineage of respiratory syncytial virus from across the globe and its evolutionary dynamics. PLoS ONE 2018, 13, e0193525. [Google Scholar] [CrossRef]
- Kamau, E.; Otieno, J.R.; Lewa, C.S.; Mwema, A.; Murunga, N.; Nokes, D.J.; Agoti, C.N. Evolution of respiratory syncytial virus genotype BA in Kilifi, Kenya, 15 years on. Sci. Rep. 2020, 10, 21176. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, S.; Venter, M. Replacement of previously circulating respiratory syncytial virus subtype B strains with the BA genotype in South Africa. J. Virol. 2011, 85, 8789–8797. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Haider, S.H.; Parveen, S.; Arshad, M.; Alsenaidy, H.A.; Baaboud, A.O.; Mobaireek, K.F.; AlSaadi, M.M.; Alsenaidy, A.M.; Sullender, W. Co-Circulation of 72bp Duplication Group A and 60bp Duplication Group B Respiratory Syncytial Virus (RSV) Strains in Riyadh, Saudi Arabia during 2014. PLoS ONE 2016, 11, e0166145. [Google Scholar] [CrossRef] [PubMed]
- Malasao, R.; Okamoto, M.; Chaimongkol, N.; Imamura, T.; Tohma, K.; Dapat, I.; Dapat, C.; Suzuki, A.; Saito, M.; Saito, M.; et al. Molecular Characterization of Human Respiratory Syncytial Virus in the Philippines, 2012–2013. PLoS ONE 2015, 10, e0142192. [Google Scholar] [CrossRef]
- Hindupur, A.; Menon, T.; Dhandapani, P. Genetic diversity of human respiratory syncytial virus in children with acute respiratory infections in Chennai, South India. Indian J. Med. Microbiol. 2019, 37, 248–254. [Google Scholar] [CrossRef]
- Feng, T.; Zhang, J.; Chen, Z.; Pan, W.; Chen, Z.; Yan, Y.; Dai, J. Glycosylation of viral proteins: Implication in virus-host interaction and virulence. Virulence 2022, 13, 670–683. [Google Scholar] [CrossRef] [PubMed]
- Cane, P.A.; Matthews, D.A.; Pringle, C.R. Identification of variable domains of the attachment (G) protein of subgroup A respiratory syncytial viruses. J. Gen. Virol. 1991, 72, 2091–2096. [Google Scholar] [CrossRef] [PubMed]
- Brakel, K.A.; Ma, Y.; Binjawadagi, R.; Harder, O.; Watts, M.; Li, J.; Binjawadagi, B.; Niewiesk, S. Codon-optimization of the respiratory syncytial virus (RSV) G protein expressed in a vesicular stomatitis virus (VSV) vector improves immune responses in a cotton rat model. Virology 2022, 575, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Otieno, J.R.; Kamau, E.M.; Agoti, C.N.; Lewa, C.; Otieno, G.; Bett, A.; Ngama, M.; Cane, P.A.; Nokes, D.J. Spread and Evolution of Respiratory Syncytial Virus A Genotype ON1, Coastal Kenya, 2010–2015. Emerg. Infect. Dis. 2017, 23, 264–271. [Google Scholar] [CrossRef]
- Tabatabai, J.; Prifert, C.; Pfeil, J.; Grulich-Henn, J.; Schnitzler, P. Novel respiratory syncytial virus (RSV) genotype ON1 predominates in Germany during winter season 2012–2013. PLoS ONE 2014, 9, e109191. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Hassan, F.; Harrison, C.J.; Dien Bard, J.; Dunn, J.; Kehl, S.; Selvarangan, R. A multi-center study to determine genetic variations in the fusion gene of respiratory syncytial virus (RSV) from children <2 years of age in the U.S. J. Clin. Virol. 2022, 154, 105223. [Google Scholar] [CrossRef] [PubMed]
- Maida, C.M.; Tramuto, F.; Di Naro, D.; Randazzo, G.; Stefanelli, P.; Marotta, C.; Reale, S.; Cernigliaro, A.; Barone, T.; Cesari, C.; et al. First detection of SARS-CoV-2 A.23.1 sub-lineage in migrants arriving to Italy via the Mediterranean Sea and public health implications. Travel. Med. Infect. Dis. 2021, 43, 102142. [Google Scholar] [CrossRef] [PubMed]
- Marotta, C.; Stefanelli, P.; Tramuto, F.; Angeloni, U.; Maida, C.M.; Cernigliaro, A.; Barone, T.; Vitale, F.; Rezza, G.; Mazzucco, W.; et al. The Dual/Global Value of SARS-CoV-2 Genome Surveillance on Migrants Arriving to Europe via the Mediterranean Routes. Ann. Glob. Health 2021, 87, 71. [Google Scholar] [CrossRef]
- Jia, R.; Lu, L.; Su, L.; Lin, Z.; Gao, D.; Lv, H.; Xu, M.; Liu, P.; Cao, L.; Xu, J. Resurgence of Respiratory Syncytial Virus Infection During COVID-19 Pandemic Among Children in Shanghai, China. Front Microbiol. 2022, 13, 938372. [Google Scholar] [CrossRef]
- Rodriguez-Fernandez, R.; Tapia, L.I.; Yang, C.F.; Torres, J.P.; Chavez-Bueno, S.; Garcia, C.; Jaramillo, L.M.; Moore-Clingenpeel, M.; Jafri, H.S.; Peeples, M.E.; et al. Respiratory Syncytial Virus Genotypes, Host Immune Profiles, and Disease Severity in Young Children Hospitalized With Bronchiolitis. J. Infect. Dis. 2017, 217, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Valley-Omar, Z.; Tempia, S.; Hellferscee, O.; Walaza, S.; Variava, E.; Dawood, H.; Kahn, K.; McMorrow, M.; Pretorius, M.; Mtshali, S.; et al. Human respiratory syncytial virus diversity and epidemiology among patients hospitalized with severe respiratory illness in South Africa, 2012–2015. Influenza Other Respir. Viruses 2022, 16, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Darbeheshti, F.; Mahdiannasser, M.; Uhal, B.D.; Ogino, S.; Gupta, S.; Rezaei, N. Interindividual immunogenic variants: Susceptibility to coronavirus, respiratory syncytial virus and influenza virus. Rev. Med. Virol. 2021, 31, e2234. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, V.A.; Hoet, B.; Hochrein, H.; De Moerlooze, L. The Quest for a Respiratory Syncytial Virus Vaccine for Older Adults: Thinking beyond the F Protein. Vaccines 2023, 11, 382. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
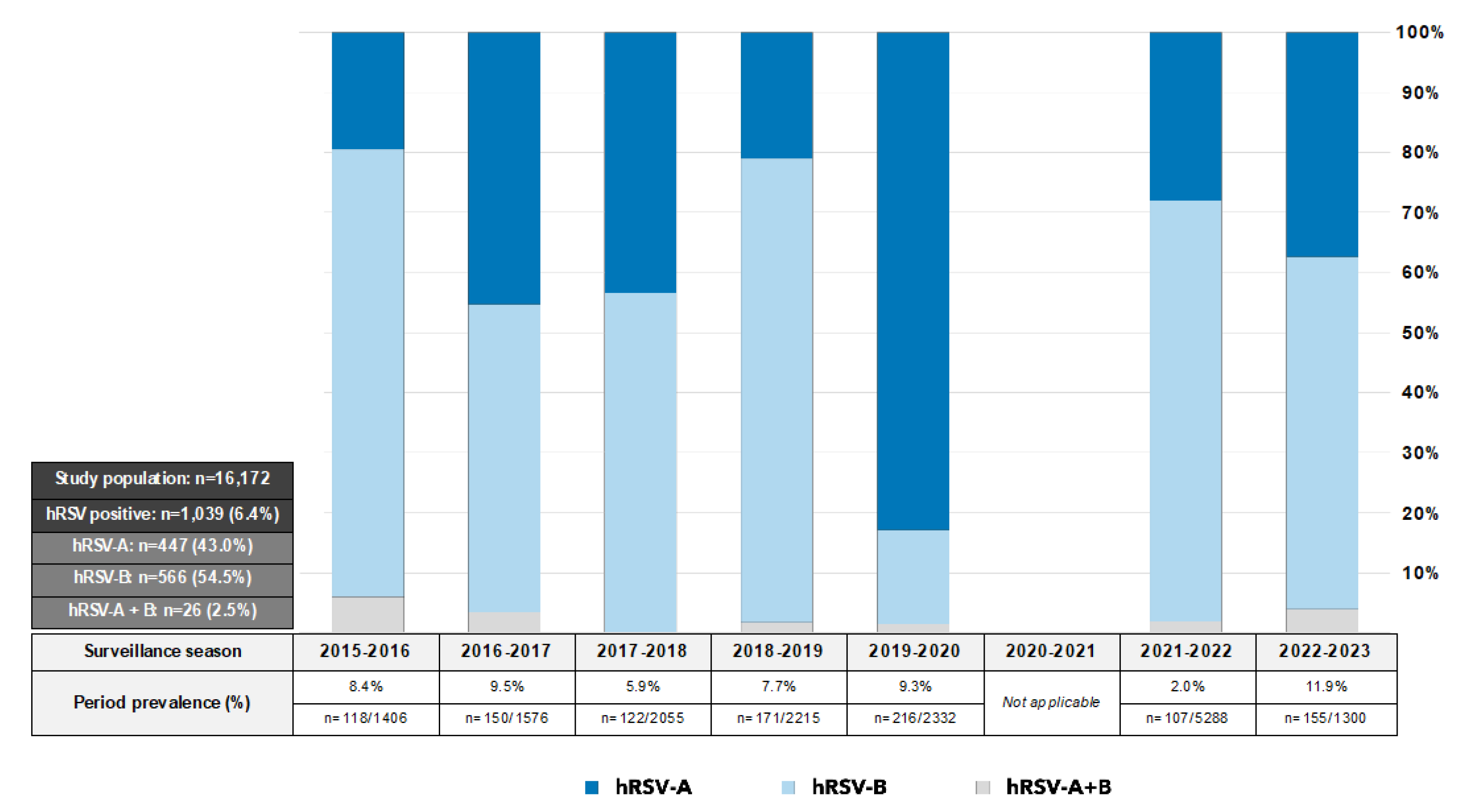
| Total Sequences | hRSV-A | hRSV-B | |
|---|---|---|---|
| Overall | 307 | 128 | 179 |
| 2015–2016 | 92 | 19 | 73 |
| 2016–2017 | 67 | 32 | 35 |
| 2017–2018 | 44 | 23 | 21 |
| 2018–2019 | 30 | 8 | 22 |
| 2019–2020 | 24 | 22 | 2 |
| 2020–2021 | NA | ||
| 2021–2022 | 23 | 11 | 12 |
| 2022–2023 | 27 | 13 | 14 |
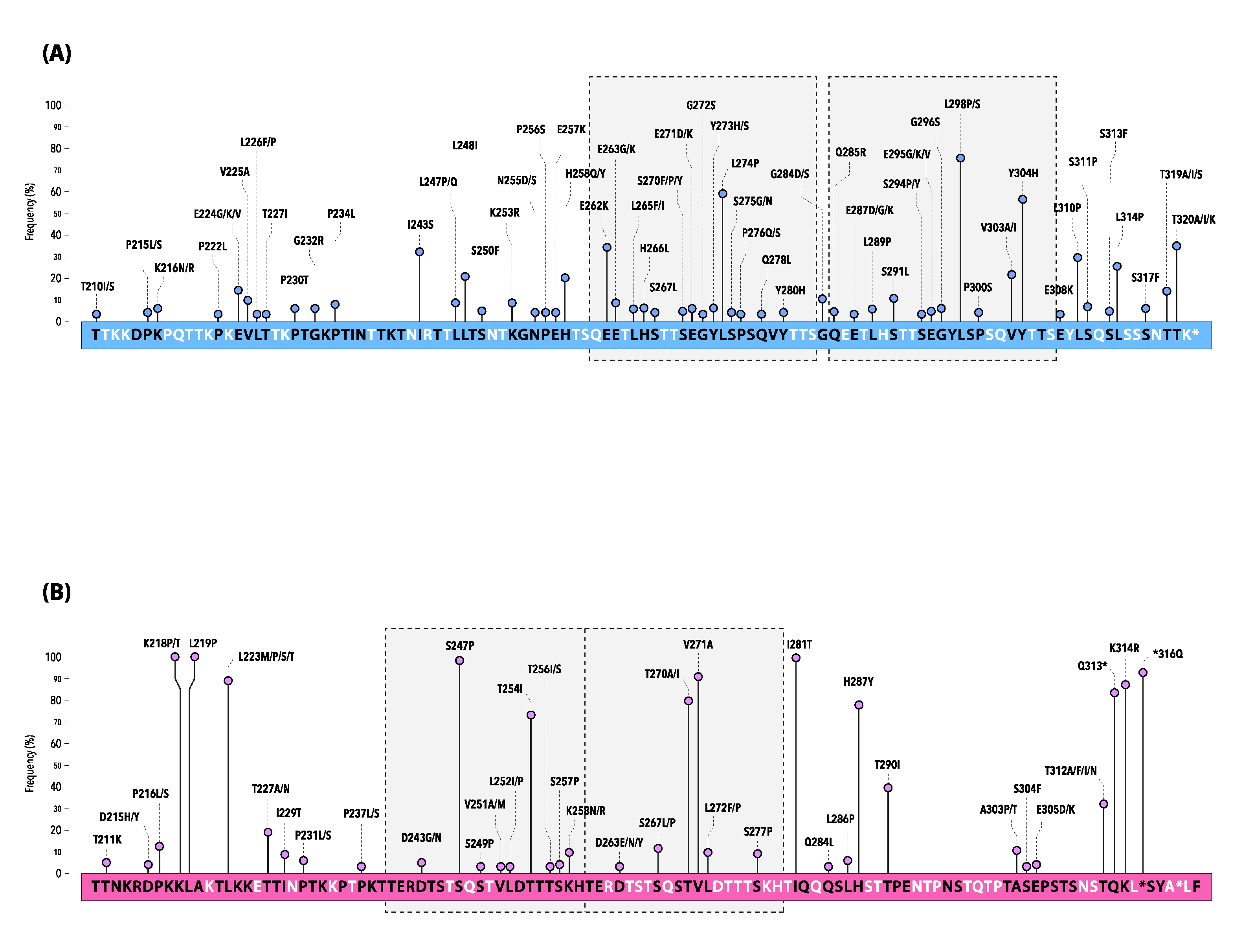
| AA Position | Frequency (%) | |
|---|---|---|
| hRSV-A | N85 | 100.0 |
| N103 | 99.2 | |
| N135 | 95.3 | |
| N237 | 99.2 | |
| N318 * | 66.4 | |
| hRSV-B | N81 | 100.0 |
| N86 | 100.0 | |
| N296 | 98.3 | |
| N310 ** | 67.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tramuto, F.; Maida, C.M.; Mazzucco, W.; Costantino, C.; Amodio, E.; Sferlazza, G.; Previti, A.; Immordino, P.; Vitale, F. Molecular Epidemiology and Genetic Diversity of Human Respiratory Syncytial Virus in Sicily during Pre- and Post-COVID-19 Surveillance Seasons. Pathogens 2023, 12, 1099. https://doi.org/10.3390/pathogens12091099
Tramuto F, Maida CM, Mazzucco W, Costantino C, Amodio E, Sferlazza G, Previti A, Immordino P, Vitale F. Molecular Epidemiology and Genetic Diversity of Human Respiratory Syncytial Virus in Sicily during Pre- and Post-COVID-19 Surveillance Seasons. Pathogens. 2023; 12(9):1099. https://doi.org/10.3390/pathogens12091099
Chicago/Turabian StyleTramuto, Fabio, Carmelo Massimo Maida, Walter Mazzucco, Claudio Costantino, Emanuele Amodio, Giuseppe Sferlazza, Adriana Previti, Palmira Immordino, and Francesco Vitale. 2023. "Molecular Epidemiology and Genetic Diversity of Human Respiratory Syncytial Virus in Sicily during Pre- and Post-COVID-19 Surveillance Seasons" Pathogens 12, no. 9: 1099. https://doi.org/10.3390/pathogens12091099
APA StyleTramuto, F., Maida, C. M., Mazzucco, W., Costantino, C., Amodio, E., Sferlazza, G., Previti, A., Immordino, P., & Vitale, F. (2023). Molecular Epidemiology and Genetic Diversity of Human Respiratory Syncytial Virus in Sicily during Pre- and Post-COVID-19 Surveillance Seasons. Pathogens, 12(9), 1099. https://doi.org/10.3390/pathogens12091099