
The Phytopathogen Fusarium verticillioides Modifies the Intestinal Morphology of the Sugarcane Borer



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Culture
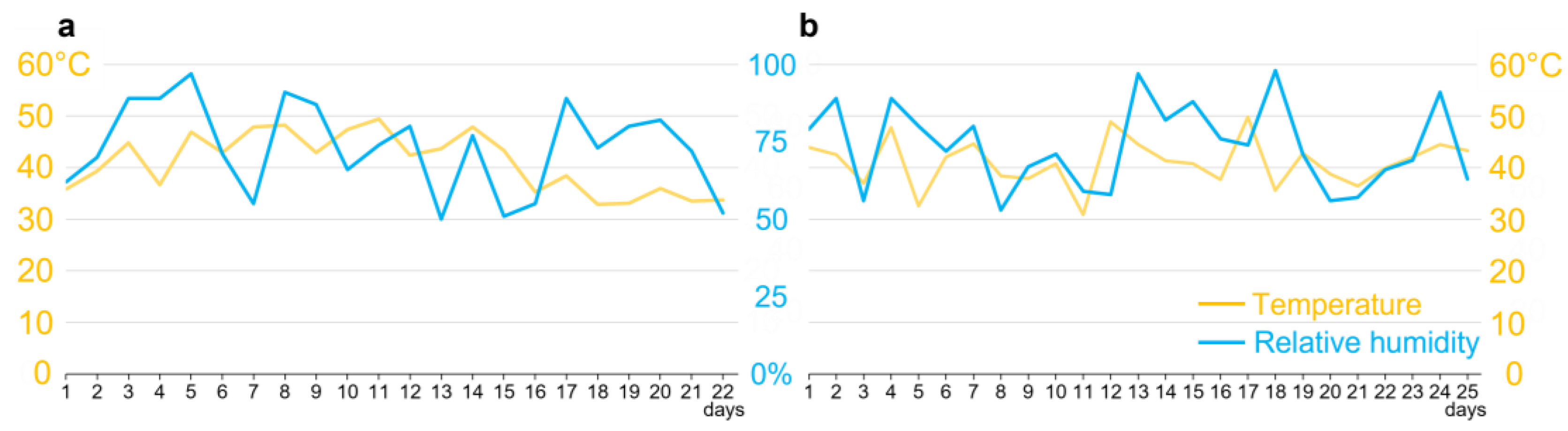
2.2. Insect Rearing
2.3. Sugarcane Cultivation
2.4. Artificial Diet Contaminated with F. verticillioides or C. falcatum
2.5. Plants Infected with F. verticillioides or C. falcatum
2.6. Contamination of Insects by Fungi
2.7. Assay with D. saccharalis Insects by Light Microscopy (LM)
2.8. Assay with Intestines of D. saccharalis by Scanning Electron Microscopy (SEM)
2.9. Experimental Design for Quantification of Intestinal Thickness of D. saccharalis
2.10. Experimental Design for Analysis of Intestinal Microvilli of D. saccharalis
3. Results
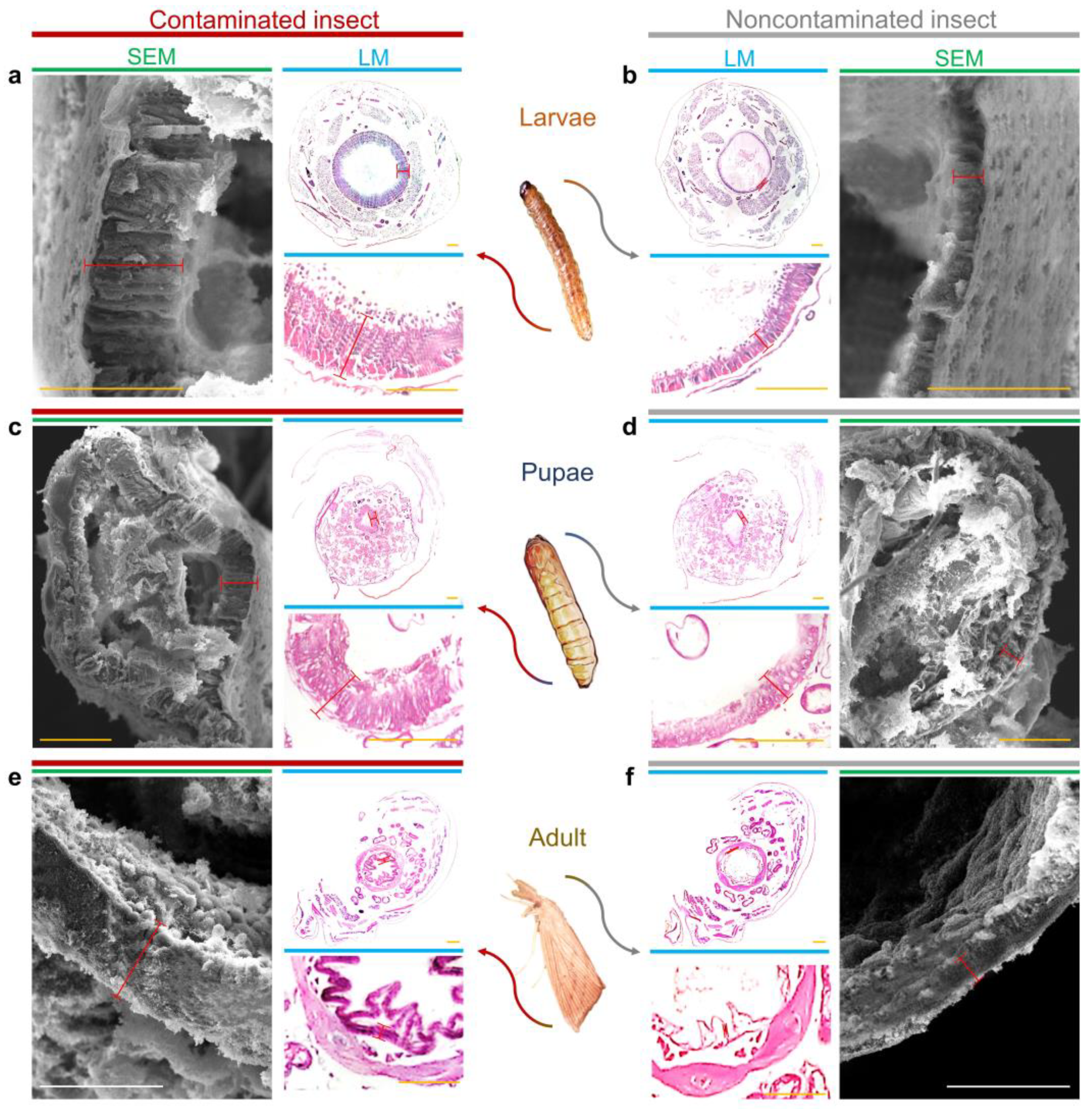
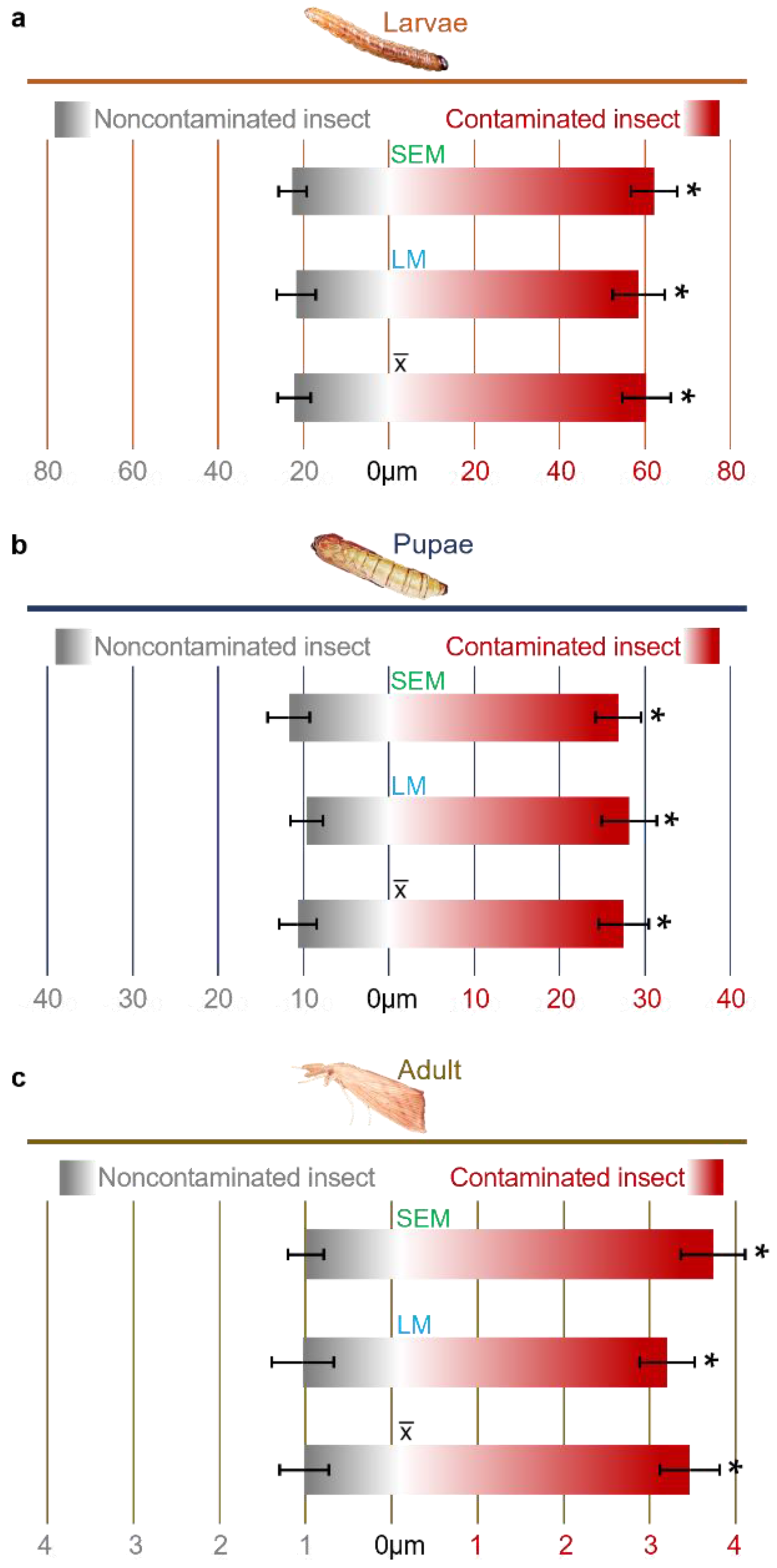
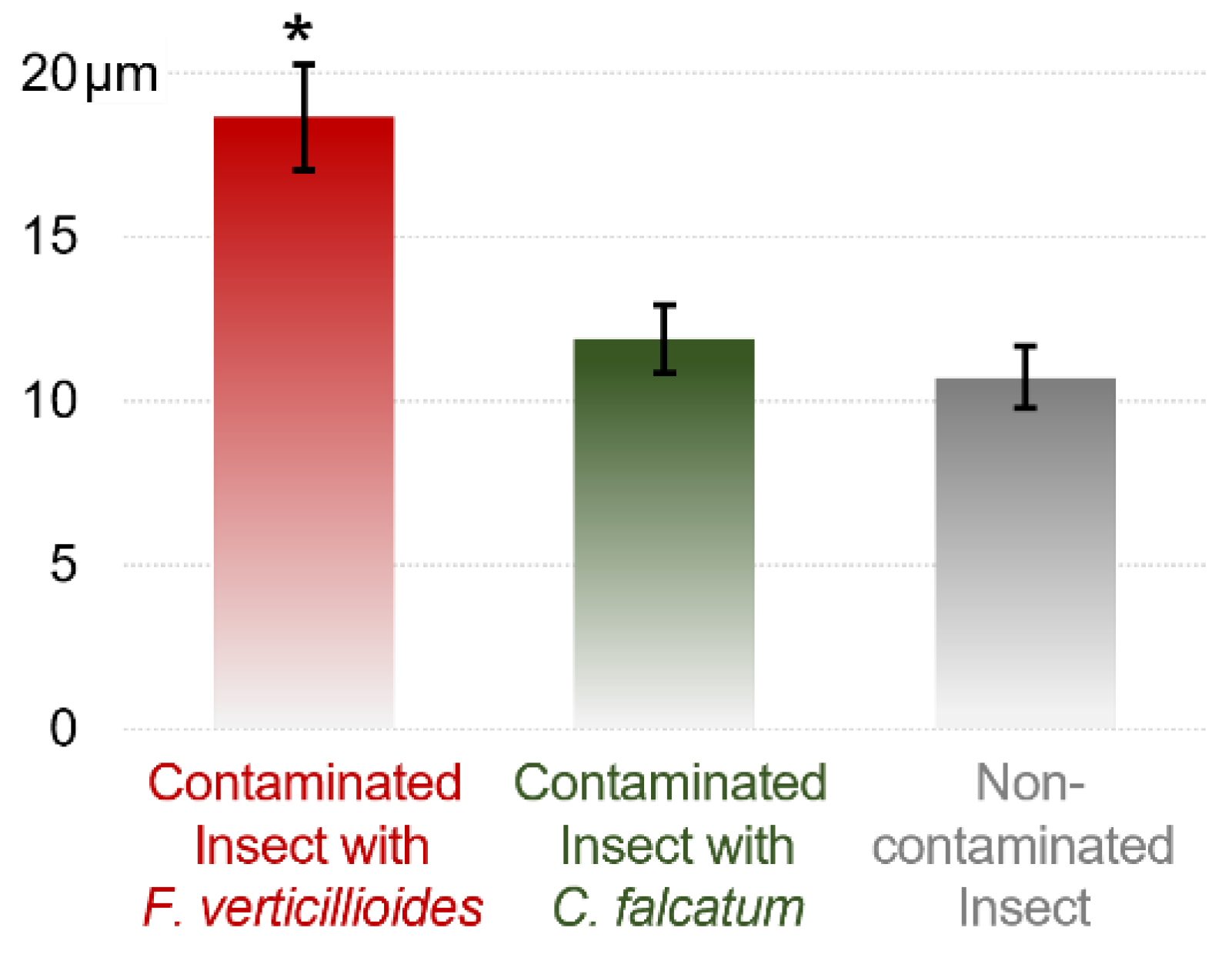
3.1. Effects of F. verticillioides on the Intestines of D. saccharalis
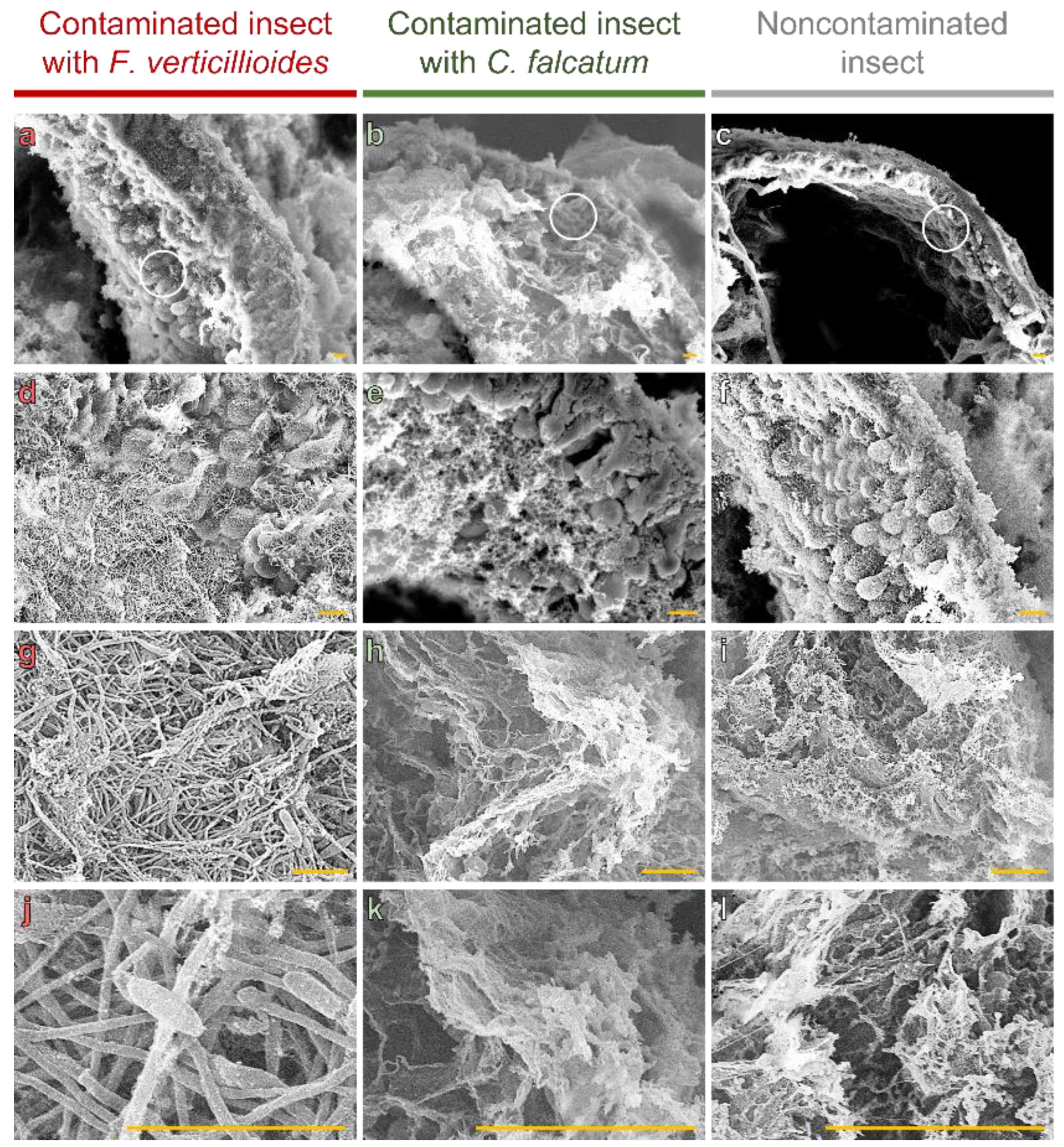
3.2. Intestinal Colonization of D. saccharalis by F. verticillioides
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Franco, F.P.; Túler, A.C.; Gallan, D.Z.; Gonçalves, F.G.; Favaris, A.P.; Peñaflor, M.F.G.; Leal, W.S.; Moura, D.S.; Bento, J.M.S.; Silva-Filho, M.C. Colletotrichum falcatum modulates the olfactory behavior of the sugarcane borer, favoring pathogen infection. FEMS Microbiol. Ecol. 2022, 98, fiac035. [Google Scholar] [CrossRef]
- Urbaneja-Bernat, P.; Waller, T.; Rodriguez-Saona, C. Repellent, oviposition-deterrent, and insecticidal activity of the fungal pathogen Colletotrichum fioriniae on Drosophila suzukii (Diptera: Drosophilidae) in highbush blueberries. Sci. Rep. 2020, 10, 14467. [Google Scholar] [CrossRef] [PubMed]
- Ratnadass, A.; Fernandes, P.; Avelino, J.; Habib, R. Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: A review. Agron. Sustain. Dev. 2012, 32, 273–303. [Google Scholar] [CrossRef] [Green Version]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. In Invertebrate Biodiversity as Bioindicators of Sustainable Landscapes; Elsevier: Amsterdam, The Netherlands, 1999; pp. 19–31. [Google Scholar]
- Gallan, D.Z.; Penteriche, A.B.; Henrique, M.O.; Silva-Filho, M.C. Sugarcane multitrophic interactions: Integrating belowground and aboveground organisms. Genet. Mol. Biol. 2022, 46, e20220163. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, S. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Douglas, A. Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Ramos, J.A.; Rojas, M.G.; Sittertz-Bhatkar, H.; Saldaña, G. Symbiotic relationship between Hypothenemus hampei (coleoptera: Scolytidae) and Fusarium solani (moniliales: Tuberculariaceae). Ann. Entomol. Soc. Am. 2000, 93, 541–547. [Google Scholar] [CrossRef]
- Cardoza, Y.J.; Teal, P.E.; Tumlinson, J.H. Effect of peanut plant fungal infection on oviposition preference by Spodoptera exigua and on host-searching behavior by Cotesia marginiventris. Environ. Entomol. 2003, 32, 970–976. [Google Scholar] [CrossRef]
- Franco, F.P.; Túler, A.C.; Gallan, D.Z.; Gonçalves, F.G.; Favaris, A.P.; Peñaflor, M.F.G.V.; Leal, W.S.; Moura, D.S.; Bento, J.M.S.; Silva-Filho, M.C. Fungal phytopathogen modulates plant and insect responses to promote its dissemination. ISME J. 2021, 15, 3522–3533. [Google Scholar] [CrossRef] [PubMed]
- Heil, M. Host manipulation by parasites: Cases, patterns, and remaining doubts. Front. Ecol. Evol. 2016, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Bosque-Perez, N.A.; Eigenbrode, S.D. The influence of virus-induced changes in plants on aphid vectors: Insights from luteovirus pathosystems. Virus Res. 2011, 159, 201–205. [Google Scholar] [CrossRef]
- Mauck, K.E.; De Moraes, C.M.; Mescher, M.C. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc. Natl. Acad. Sci. USA 2010, 107, 3600–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoletti, R.; Becchimanzi, A. Ecological and molecular interactions between insects and fungi. Microorganisms 2022, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Naundrup, A.; Bohman, B.; Kwadha, C.A.; Jensen, A.B.; Becher, P.G.; De Fine Licht, H.H. Pathogenic fungus uses volatiles to entice male flies into fatal matings with infected female cadavers. ISME J. 2022, 16, 2388–2397. [Google Scholar] [CrossRef]
- Soldano, A.; Alpizar, Y.A.; Boonen, B.; Franco, L.; Lopez-Requena, A.; Liu, G.; Mora, N.; Yaksi, E.; Voets, T.; Vennekens, R.; et al. Gustatory-mediated avoidance of bacterial lipopolysaccharides via TRPA1 activation in Drosophila. Elife 2016, 5, e13133. [Google Scholar] [CrossRef]
- Evans, H.C.; Elliot, S.L.; Hughes, D.P. Hidden diversity behind the zombie-ant fungus Ophiocordyceps unilateralis: Four new species described from carpenter ants in Minas Gerais, Brazil. PLoS ONE 2011, 6, e17024. [Google Scholar] [CrossRef] [PubMed]
- Steinert, M.; Hentschel, U.; Hacker, J. Symbiosis and pathogenesis: Evolution of the microbe-host interaction. Die Nat. 2000, 87, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; Zilber-Rosenberg, I. Symbiosis and development: The hologenome concept. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 56–66. [Google Scholar] [CrossRef]
- Hooper, L.V. Do symbiotic bacteria subvert host immunity? Nat. Rev. Microbiol. 2009, 7, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Grell, K.G.; Grell, K.G. Parasitism and Symbiosis. Protozoology 1973, 350–364. [Google Scholar] [CrossRef]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajek, A.; St. Leger, R. Interactions between fungal pathogens and insect hosts. Annu. Rev. Entomol. 1994, 39, 293–322. [Google Scholar] [CrossRef]
- Butt, T.; Coates, C.; Dubovskiy, I.; Ratcliffe, N. Entomopathogenic fungi: New insights into host–pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef] [PubMed]
- Cory, J.S.; Hoover, K. Plant-mediated effects in insect–pathogen interactions. Trends Ecol. Evol. 2006, 21, 278–286. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. Insect-microbe mutualism without vertical transmission: A stinkbug acquires a beneficial gut symbiont from the environment every generation. Appl. Environ. Microbiol. 2007, 73, 4308–4316. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Hosokawa, T.; Fukatsu, T. Diversity of bacterial symbiosis in stinkbugs. In Microbial Ecology Research Trends; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2008; pp. 39–63. [Google Scholar]
- Feldhaar, H.; Gross, R. Insects as hosts for mutualistic bacteria. Int. J. Med. Microbiol. 2009, 299, 1–8. [Google Scholar] [CrossRef]
- Rajagopal, R. Beneficial interactions between insects and gut bacteria. Indian J. Microbiol. 2009, 49, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Rani, A.; Sharma, A.; Rajagopal, R.; Adak, T.; Bhatnagar, R.K. Bacterial diversity analysis of larvae and adult midgut microflora using culture-dependent and culture-independent methods in lab-reared and field-collected Anopheles stephensi-an Asian malarial vector. BMC Microbiol. 2009, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Gu, F.; Ai, S.; Chen, Y.; Jin, S.; Xie, X.; Zhang, T.; Zhong, G.; Yi, X. Mutualism promotes insect fitness by fungal nutrient compensation and facilitates fungus propagation by mediating insect oviposition preference. ISME J. 2022, 16, 1831–1842. [Google Scholar] [CrossRef]
- Ogunwolu, E.; Reagan, T.; Flynn, J.; Hensley, S. Effects of Diatraea saccharalis (F.)(Lepidoptera: Pyralidae) damage and stalk rot fungi on sugarcane yield in Louisiana. Crop Prot. 1991, 10, 57–61. [Google Scholar] [CrossRef]
- Franco, F.P.; Moura, D.S.; Vivanco, J.M.; Silva-Filho, M.C. Plant–insect–pathogen interactions: A naturally complex ménage à trois. Curr. Opin. Microbiol. 2017, 37, 54–60. [Google Scholar] [CrossRef]
- King, E.; Hartley, G. Diatraea saccharalis. Handb. Insect Rearing 1985, 2, 265–270. [Google Scholar]
- Karnovsky, M. A formaldehyde-glutaraldehyde fixative of high osmolarity for use in electron microscopy. J. Cell Biol. 1965, 27, 137A–138A. [Google Scholar]
- Ruiz, L.M.; Segura, C.; Trujillo, J.; Orduz, S. In vivo binding of the Cry11Bb toxin of Bacillus thuringiensis subsp. medellin to the midgut of mosquito larvae (Diptera: Culicidae). Memórias Do Inst. Oswaldo Cruz 2004, 99, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Horridge, G.A.; Tamm, S. Critical point drying for scanning electron microscopic study of ciliary motion. Science 1969, 163, 817–818. [Google Scholar] [CrossRef]
- Ma, E.; Zhu, Y.; Liu, Z.; Wei, T.; Wang, P.; Cheng, G. Interaction of viruses with the insect intestine. Annu. Rev. Virol. 2021, 8, 115–131. [Google Scholar] [CrossRef]
- Medeiros, A.H.; Franco, F.P.; Matos, J.L.; de Castro, P.A.; Santos-Silva, L.K.; Henrique-Silva, F.; Goldman, G.H.; Moura, D.S.; Silva-Filho, M.C. Sugarwin: A sugarcane insect-induced gene with antipathogenic activity. Mol. Plant-Microbe Interact. 2012, 25, 613–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, F.P.; Santiago, A.C.; Henrique-Silva, F.; de Castro, P.A.; Goldman, G.H.; Moura, D.S.; Silva-Filho, M.C. The sugarcane defense protein SUGARWIN2 causes cell death in Colletotrichum falcatum but not in non-pathogenic fungi. PLoS ONE 2014, 9, e91159. [Google Scholar] [CrossRef]
- Franco, F.P.; Dias, R.O.; Toyama, D.; Henrique-Silva, F.; Moura, D.S.; Silva-Filho, M.C. Structural and Functional Characterization of PR-4 SUGARWINs From Sugarcaneand Their Role in Plant Defense. Front. Plant Sci. 2019, 9, 1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [Green Version]
- Peñaflor, M.F.G.; Bento, J.M.S. Red-rot infection in sugarcane attenuates the attractiveness of sugarcane borer-induced plant volatiles to parasitoid. Arthropod-Plant Interact. 2019, 13, 117–125. [Google Scholar] [CrossRef]
- Shapira, M. Gut microbiotas and host evolution: Scaling up symbiosis. Trends Ecol. Evol. 2016, 31, 539–549. [Google Scholar] [CrossRef]
- Bution, M.L.; Caetano, F.H. The midgut of Cephalotes ants (Formicidae: Myrmicinae): Ultrastructure of the epithelium and symbiotic bacteria. Micron 2010, 41, 448–454. [Google Scholar] [CrossRef]
- Bosch, I. Symbiosis between bacteria and oceanic clonal sea star larvae in the western North Atlantic Ocean. Mar. Biol. 1992, 114, 495–502. [Google Scholar] [CrossRef]
- Duperron, S.; Gros, O. Colwellia and sulfur-oxidizing bacteria: An unusual dual symbiosis in a Terua mussel (Mytilidae: Bathymodiolinae) from whale falls in the Antilles arc. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 115, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Aubé, J.; Cambon-Bonavita, M.-A.; Velo-Suárez, L.; Cueff-Gauchard, V.; Lesongeur, F.; Guéganton, M.; Durand, L.; Reveillaud, J. A novel and dual digestive symbiosis scales up the nutrition and immune system of the holobiont Rimicaris exoculata. Microbiome 2022, 10, 189. [Google Scholar] [CrossRef]
- Caetano, F.; Torres, A., Jr.; Camargo-Mathias, M.; Tomotake, M. Apocrine secretion in the ant, Pachycondyla striata, ventriculus (Formicidae: Ponerinae). Cytobios 1994, 80, 235–242. [Google Scholar]
- Caetano, F.; Jaffe, K.; Crewe, R. The digestive tract of Cataulacus ants: Presence of microorganisms in the ileum. In Les Insectes Sociaux; Lenoir, A., Arnold, G., Lepage, M., Eds.; Éditions Quæ: Versailles, France, 1994; p. 391. [Google Scholar]
- Terra, W.R.; Barroso, I.G.; Dias, R.O.; Ferreira, C. Molecular physiology of insect midgut. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 56, pp. 117–163. [Google Scholar]
- Nardon, P.; Charles, H. Morphological aspects of symbiosis. Symbiosis: Mech. Model Syst. 2002, 4, 13–44. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T.; Matsumoto, S.; Simon, J.-C.; Fukatsu, T. Symbiotic bacterium modifies aphid body color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef]
- Vigneron, A.; Masson, F.; Vallier, A.; Balmand, S.; Rey, M.; Vincent-Monégat, C.; Aksoy, E.; Aubailly-Giraud, E.; Zaidman-Rémy, A.; Heddi, A. Insects recycle endosymbionts when the benefit is over. Curr. Biol. 2014, 24, 2267–2273. [Google Scholar] [CrossRef] [Green Version]
- Frolov, A.O.; Kostygov, A.Y.; Yurchenko, V. Development of monoxenous trypanosomatids and phytomonads in insects. Trends Parasitol. 2021, 37, 538–551. [Google Scholar] [CrossRef]
- Żuwała, K.; Jakubowski, M. Structural diversification of the gustatory organs during metamorphosis in the alpine newt Triturus alpestris. J. Anat. 2007, 211, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, O.; Miguel-Aliaga, I.; Trajkovski, M. Intestinal plasticity and metabolism as regulators of organismal energy homeostasis. Nat. Metab. 2022, 4, 1444–1458. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uncontamnated Insect | Contaminated Insect with C. falcatum | Contaminated Insect with F. verticillioides | p Value | |
|---|---|---|---|---|
| Larvae | ||||
| Artificial Diet | 22.57 ± 3.23 a | 23.97 ± 3.02 a | 62.23 ± 5.39 b | >0.01 |
| Sugarcane Plants | 25.07 ± 4.75 a | 20.03 ± 5.17 a | 56.53 ± 6.63 b | >0.01 |
| p value | 0.2927 | 0.1160 | 0.1085 | |
| Pupae | ||||
| Artificial Diet | 11.73 ± 2.45 a | 14.47 ± 2.33 a | 26.90 ± 2.68 b | >0.01 |
| Sugarcane Plants | 15.17 ± 3.37 a | 13.90 ± 2.68 a | 23.47 ± 5.87 b | >0.01 |
| p value | 0.0641 | 0.5813 | 0.1736 | |
| Adult | ||||
| Artificial Diet | 1.10 ± 0.24 a | 1.47 ± 0.24 a | 3.63 ± 0.37 b | >0.01 |
| Sugarcane Plants | 1.57 ± 0.41 a | 1.33 ± 0.33 a | 3.07 ± 0.75 b | >0.01 |
| p value | 0.0605 | 0.4518 | 0.3301 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallan, D.Z.; Henrique, M.O.; Silva-Filho, M.C. The Phytopathogen Fusarium verticillioides Modifies the Intestinal Morphology of the Sugarcane Borer. Pathogens 2023, 12, 443. https://doi.org/10.3390/pathogens12030443
Gallan DZ, Henrique MO, Silva-Filho MC. The Phytopathogen Fusarium verticillioides Modifies the Intestinal Morphology of the Sugarcane Borer. Pathogens. 2023; 12(3):443. https://doi.org/10.3390/pathogens12030443
Chicago/Turabian StyleGallan, Diego Z., Maressa O. Henrique, and Marcio C. Silva-Filho. 2023. "The Phytopathogen Fusarium verticillioides Modifies the Intestinal Morphology of the Sugarcane Borer" Pathogens 12, no. 3: 443. https://doi.org/10.3390/pathogens12030443
APA StyleGallan, D. Z., Henrique, M. O., & Silva-Filho, M. C. (2023). The Phytopathogen Fusarium verticillioides Modifies the Intestinal Morphology of the Sugarcane Borer. Pathogens, 12(3), 443. https://doi.org/10.3390/pathogens12030443