1. Introduction
Streptococcus suis is an important zoonotic pathogen that primarily causes meningitis, sepsis, endocarditis, arthritis, and pneumonia in pigs and humans [
1]. Recently,
S. suis serotypes 20, 22, and 26 were reclassified as
Streptococcus parasuis [
2]. Although most
S. parasuis strains have been isolated from the saliva of clinically healthy pigs, the presence of
S. parasuis in diseased pigs and calves with pneumonia or systemic infection (meningitis, arthritis, endocarditis, or septicemia) indicates that this species may be pathogenic in pigs and calves [
3,
4,
5,
6]. In our previous study, two
S. parasuis clinical strains, BS26 and BS27, were isolated from a patient with peritonitis and a patient with pneumonia and arthritis, respectively [
7]. In the present study, a third
S. parasuis clinical strain, NN1, was isolated from a patient with pneumonia. This indicates that the zoonotic pathogen
S. parasuis is an emerging threat to public health and underscores the urgent need to further evaluate the pathogenesis of
S. parasuis clinical strains. In our previous study, the capacity of
S. parasuis clinical strains to cause systemic infections and histopathological lesions in lungs and livers of infected C57BL/6 mice were revealed [
7]. However, the mechanism of immunity response against the infection caused by
S. parasuis is still poorly understood. An important hallmark of the pathogenicity of
S. suis is its ability to cross the blood–brain barrier (BBB) and induce an inflammatory response in the brain, which leads to damage to the central nervous system (CNS) [
8]. Our previous study demonstrated the capacity of
S. parasuis clinical strains to enter the CNS of infected mice [
7]. In the present study, we investigated the characteristics and mechanism of cerebral inflammation against the infection caused by
S. parasuis clinical strains. Firstly, we evaluated the proportion and time of the neurological symptoms developed in mice infected with two S. parasuis clinical strains. In addition, the characteristics of histopathological changes and the cerebral immune response in mice with neurological symptoms were analyzed. Furthermore, we explored the roles of microglia and astrocytes in the
S. parasuis strain-induced cerebral inflammation.
3. Results
3.1. Evaluation of the Genomic Characteristics, Antimicrobial Profile, and Virulence Genes of S. parasuis Strain NN1
S. parasuis strain NN1 showed 100% 16S rRNA gene sequence similarity with strains BS26 and BS27. In addition, the phylogenetic tree based on the alignment of core genes of eight
S. parasuis genomes and one
S. suis genome indicated that NN1 had a phylogenetic affinity with the other two clinical strains BS26 and BS27 (
Figure S1). Strains 4253 and H35 were clustered into the same lineage with clinical strains. The phylogenetic analysis based on the gene presence and absence of 14
S. parasuis strains also clustered strains 4253 and H35 into the same clade with two clinical strains BS26 and BS27 [
18]. The
cps gene cluster of NN1 was clustered into type X and shared 99.2% sequence similarity with BS26 [
7].
NN1 was resistant to tetracycline, erythromycin, and azithromycin, with MIC values of 16 μg/mL, 32 μg/mL, and >256 μg/mL, respectively. The tetracycline resistance gene tet(M) was present in NN1, whereas genes encoding tetracycline resistance were absent in BS26 and BS27. Three S. parasuis clinical strains harbored msr(D) and mef(A) genes encoding macrolides resistance. The presence of two copies of msr(D) and mef(A) in NN1 may partially explain its higher resistance to erythromycin and azithromycin in comparison with BS26 and BS27.
NN1 was susceptible to penicillin G, cefaclor, clindamycin, streptomycin, kanamycin, spectinomycin, gentamicin, vancomycin, rifampicin, chloramphenicol, and trimethoprim–sulfamethoxazole.
The presence of 129 putative virulence genes of
S. suis were investigated in three
S. parasuis clinical strains. No significant difference in the distribution of putative virulence genes was observed among
S. parasuis clinical strains. Indeed, 47 of them were present in all three
S. parasuis clinical strains. In addition, the
oppa gene encoding the ABC transporter substrate-binding protein was only present in
S. parasuis clinical strains BS26 and BS27, whereases the
hhly3 gene encoding amino acid transporter protein was only present in
S. parasuis clinical strain NN1 (
Supplemental Table S1).
3.2. Survival Rates of Mice Infected with S. suis Strain P1/7, S. parasuis Strain BS26, and S. parasuis Strain NN1 Differed
The survival level of mice infected with NN1 was significantly higher than those of mice infected with BS26 and
S. suis strain P1/7. Mice infected 5 × 10
7 CFUs of NN1 had a 90% survival rate at 24 h post-infection, whereas the survival rates of mice infected with
S. suis strain P1/7 and
S. parasuis strain BS26 were 10% and 30% at the same time point, respectively. At 72 h post-infection, the survival rate of mice infected with NN1 (75%) was significantly higher than those of mice infected with
S. suis strain P1/7 (10%) and
S. parasuis strain BS26 (20%) (
Figure S2).
3.3. Two S. parasuis Strains Induced Neurological Symptoms in Infected Mice from 24 h Post-Infection
To reduce the mortality of the infected mice and obtain enough observations, the infection dose decreased to 2 × 107 CFUs for each strain. In the present study, 100% (40/40), 77% (22/30), 45% (9/20), and 40% (4/10) of mice infected with S. parasuis strain BS26 survived at 12 h, 24 h, 48 h, and 72 h post-infection, respectively. The survival rates of mice infected with NN1 at 12 h, 24 h, 48 h, and 72 h post-infection were 100% (40/40), 100% (30/30), 80% (16/20), and 60% (6/10), respectively. Compared with the two S. parasuis strains, S. suis strain P1/7 possessed a higher capacity to induce lethal infection at 24 h post-infection; the survival rates at 12 h, 24 h, 48 h, and 72 h post-infection were 83% (33/40), 27% (8/30), 20% (4/20), and 20% (2/10), respectively.
The survival mice infected with BS26 and NN1 displayed neurological symptoms from 24 h post-infection. The appearance time and proportion of neurological symptoms in survival mice differed between mice infected with BS26 and NN1. Among the mice infected with BS26, half (20/40) of the survival mice displayed ataxia of mainly paresis of the hindlimb (median time 24 h), and 30% (6/20) of the survival mice with paresis of the hindlimb also displayed bending of the head toward one side and walking in circles as the infection progressed (median time 60 h). Among the mice infected with NN1, 45% (18/40) of the survival mice displayed ataxia of mainly paresis of the hindlimb (median time 48 h), and over 20% (4/18) of the survival mice with paresis of the hindlimb also displayed bending of the head toward one side and walking in circles as the infection progressed (median time 72 h). However, we could not rule out the possibility that the mice sacrificed at earlier time points would have developed neurological symptoms at later time points; thus, the proportion of survival mice with neurological symptoms may be underestimated.
Notably, no neurological symptoms were observed in survival mice infected with
S. suis strain P1/7 before 72 h post-infection. This result was in agreement with a previous study, which reported the appearance of clinical symptoms of meningitis in
S. suis ST1-infected mice from four days post-infection [
19].
3.4. Investigation of Histopathological Changes in the Brains of Survival Mice with Neurological Symptoms
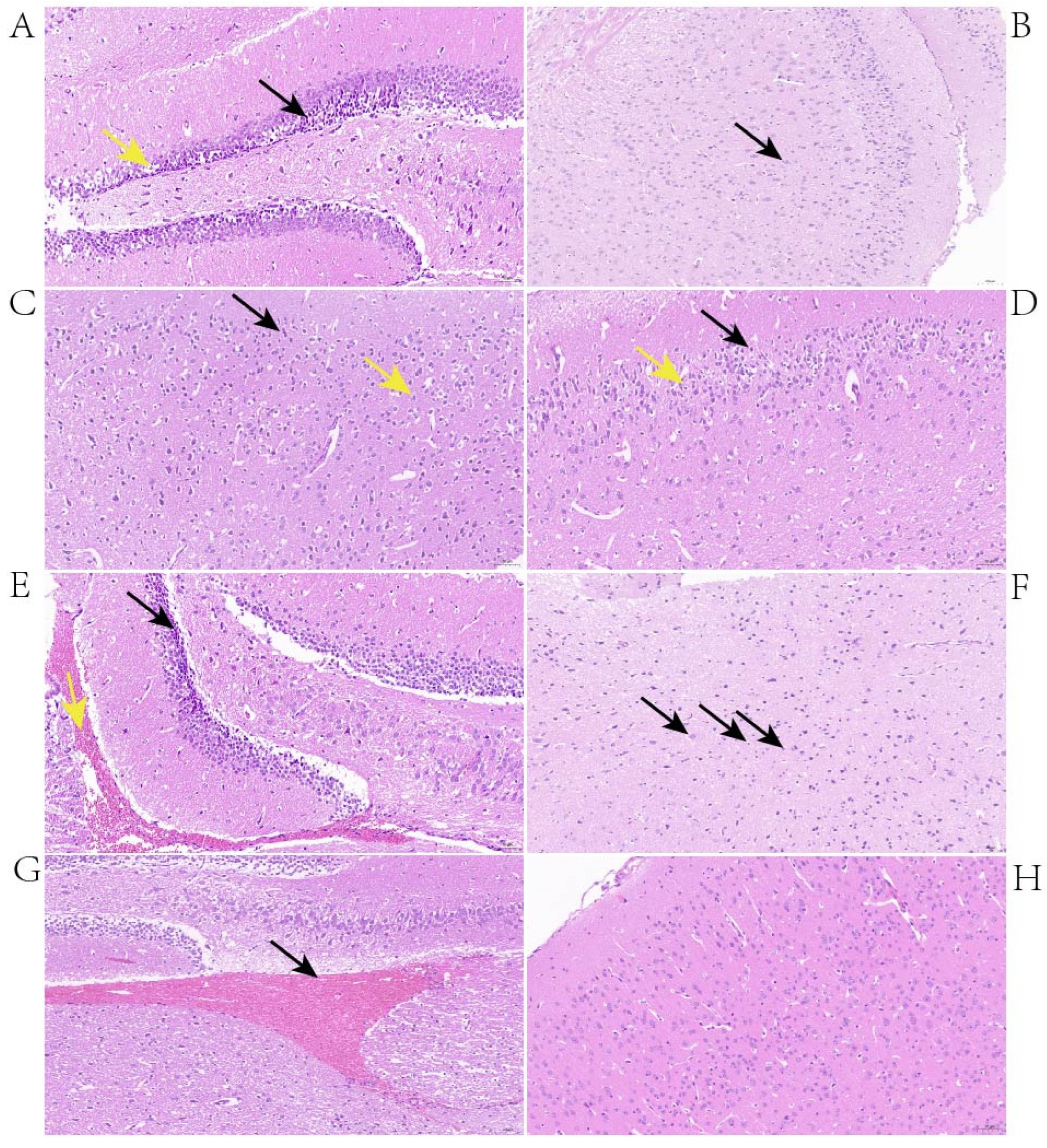
The histopathological changes in the brains of mice with neurological symptoms were investigated. At 24 h post-infection, the changes observed in the brains of mice infected with BS26 were neuronal atrophy, neuronal deformation, and vacuolization of the neuronal cytoplasm in the hippocampus (
Figure 1A). In addition, the brains of BS26-infected mice exhibited the engulfment of neuronal debris by microglia from 24 h post-infection (
Figure 1B). At the same time point, the histopathological changes in mice infected with NN1 appeared in the cerebral cortex and included neuronal deformation, vacuolization of the neuronal cytoplasm, and microgliosis (
Figure 1C,D). At 48 h post-infection, the aforementioned neuronal atrophy, neuronal deformation, and vacuolization of the neuronal cytoplasm appeared in both the hippocampus and the cerebral cortex of mice infected with BS26. In contrast, at the same time point, the significant histopathological changes which appeared in the brains of NN1-infected mice were hemorrhagic foci (
Figure 1E). At 72 h post-infection, the significant histopathological changes in the brains of BS26-infected mice with neurological symptoms were slight neutrophil infiltration (
Figure 1F). In contrast, hemorrhagic foci had appeared in the brains of almost all NN1-infected mice with neurological symptoms at the same time point (
Figure 1G).
3.5. Both S. parasuis Strains Induced Rapid Upregulation of Proinflammatory Mediators and TLR Genes Transcription in the Brains of Mice with Neurological Symptoms
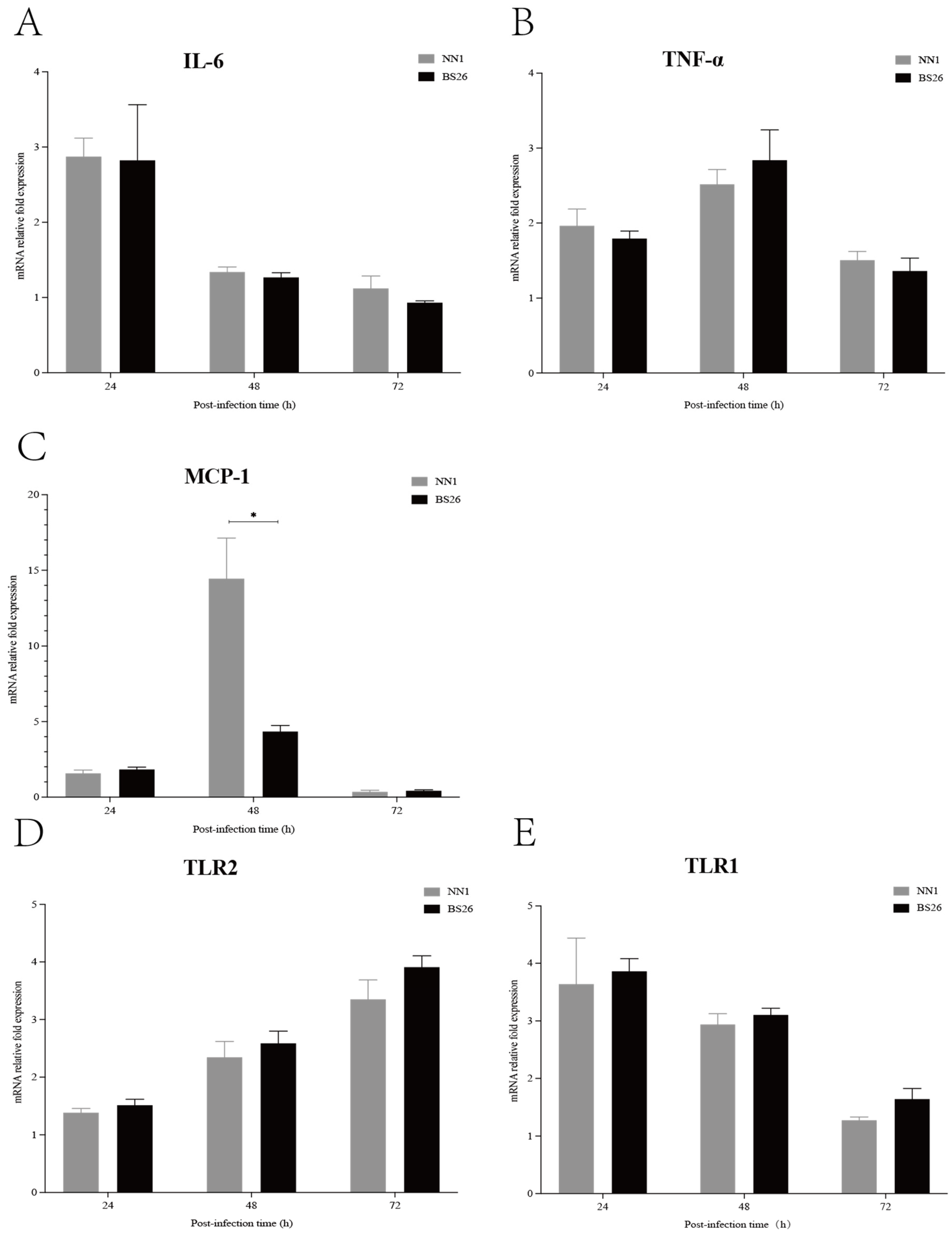
The transcription levels of IL-6, TNF-α, and MCP-1 genes in the brains of mice with neurological symptoms were investigated. The transcription levels of the IL-6 gene in the brains of mice infected with both
S. parasuis strains were slightly upregulated at 24 h post-infection and then gradually returned to the baseline level at 48 h post-infection (
Figure 2A). The transcription patterns of the TNF-α and MCP-1 genes were similar in mice infected with the two
S. parasuis strains that showed neurological symptoms, which slightly upregulated at 24 h post-infection, peaked at 48 h post-infection and then gradually returned to the baseline level at 72 h post-infection (
Figure 2B,C). No significant differences in transcription levels of the IL-6 and TNF-α genes were observed between mice infected with BS26 and NN1 throughout the experiment (
Figure 2A,B), whereas a significantly higher transcription level of the MCP-1 gene was observed in NN1-infected mice at 48 h post-infection (
Figure 2C).
TLRs play a pivotal role in the innate immune response by recognizing the pathogen-associated molecular patterns (PAMPs) of invading pathogens. In the present study, we investigated the kinetics of TLR1, TLR2, TLR6, and TLR9 gene transcription in the brains of infected mice with neurological symptoms. TLR2 gene transcription patterns in the brains of BS26- and NN1-infected mice with neurological symptoms were upregulated at 48 h post-infection and peaked at 72 h post-infection (
Figure 2D). Notably, TLR1 gene transcription was also upregulated in the brains of BS26- and NN1-infected mice with neurological symptoms, but a different transcription pattern in which transcription peaked at 24 h post-infection and then gradually decreased to the baseline level at 72 h post-infection was observed (
Figure 2E). The transcription levels of the TLR1 and TLR2 genes in the brains of BS26- and NN1-infected mice with neurological symptoms were no different (
Figure 2D,E).
3.6. The Capacity of S. parasuis Strains to Induce Proinflammatory Mediators Production by Primary Astrocytes and BV2 Cells Was Higher Than That of S. suis Strain P1/7 at the Early Phase of Interaction
Activated astrocytes and microglia played critical roles in the CNS inflammatory response against
S. suis strains infection by producing proinflammatory mediators [
14,
20,
21]. In this study, the capacity of
S. parasuis strains to induce proinflammatory cytokine and chemokine production by primary astrocytes and BV2 cells was investigated and compared with that of
S. suis strain P1/7.
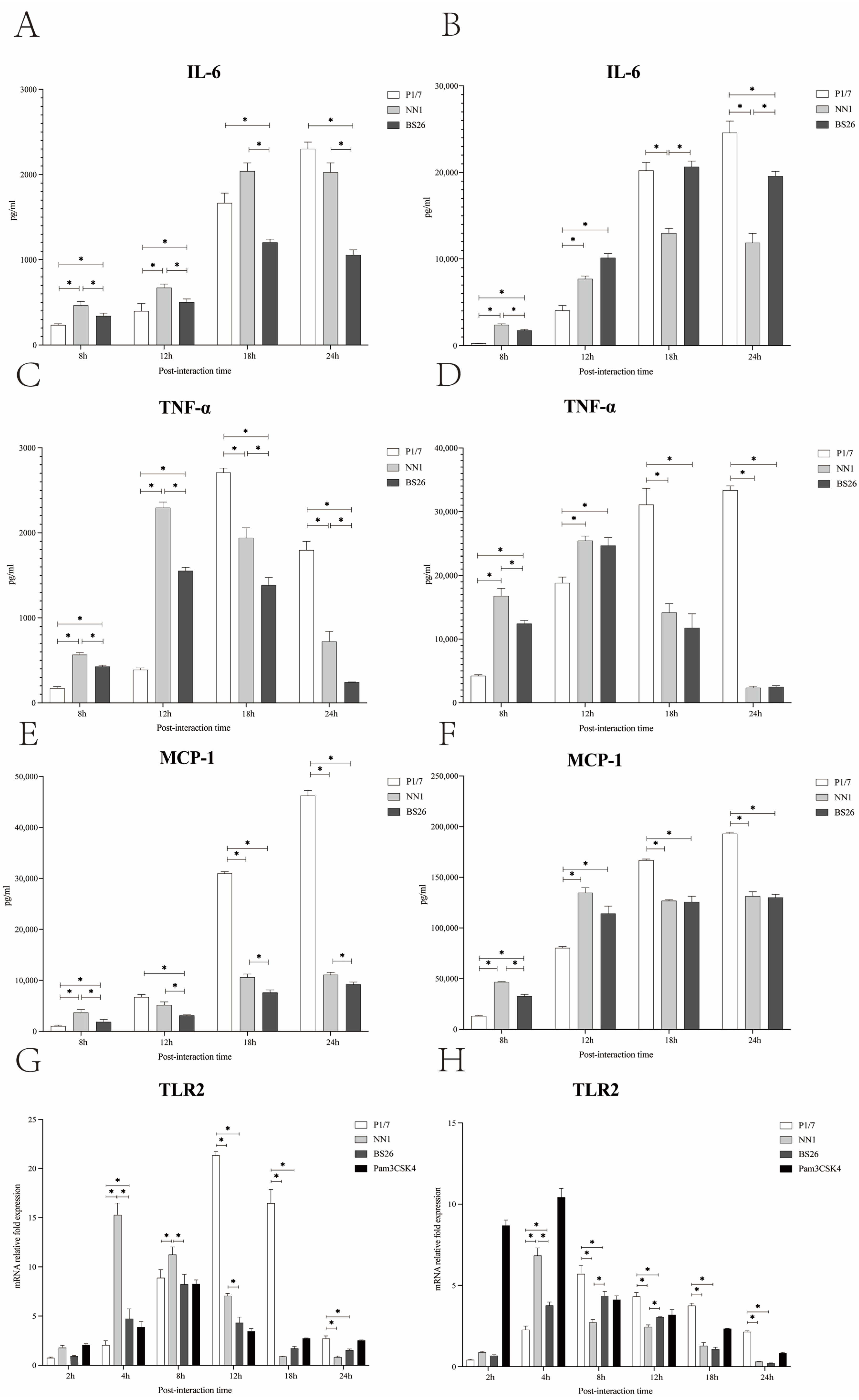
The levels of IL-6 in both primary astrocytes and BV2 cells infected with the two
S. parasuis strains peaked at 18 h post-infection and then gradually decreased at 24 h post-infection (
Figure 3A,B). The levels of TNF-α in both primary astrocytes and BV2 cells infected with the two
S. parasuis strains peaked at 12 h post-infection and gradually decreased thereafter (
Figure 3C,D). Of the two
S. parasuis strains, NN1 possessed a higher capacity to induce IL-6, TNF-α, and MCP-1 production by primary astrocytes throughout the experiment, and by BV2 cells at 8 h post-infection, whereas BS26 induced higher IL-6 production by BV2 cells from 18 h post-infection (
Figure 3A–F).
The differences between
S. suis strain P1/7 and the two
S. parasuis strains in the kinetics of the proinflammatory mediators secreted by primary astrocytes and BV2 cells were observed. At 8 h and 12 h post-infection, the levels of TNF-α, IL-6, and MCP-1 in both primary astrocytes and BV2 cells infected with the two
S. parasuis strains were significantly higher than those of cells infected with
S. suis strain P1/7, except for the MCP-1 in primary astrocytes at 12 h post-infection (
Figure 3A–F). As the infection progressed, TNF-α and MCP-1 levels in both primary astrocytes and BV2 cells infected with
S. suis strain P1/7 were significantly higher than those of cells infected with the two
S. parasuis strains at 18 h and 24 h post-infection (
Figure 3C–F). The levels of IL-6 in both primary astrocytes and BV2 cells infected with
S. suis strain P1/7 peaked at 24 h post-infection (
Figure 3A,B). At 24 h post-infection,
S. suis strain P1/7 induced a significantly higher level of IL-6 production by BV2 cells than the two
S. parasuis strains (
Figure 3B).
3.7. The Activation of Primary Astrocytes and BV2 Cells by S. parasuis Strains Was Mediated by TLR2
In this study, the kinetics of the TLR1, 2, 6, and 9 genes transcription in primary astrocytes and BV2 cells infected with the two
S. parasuis strains were investigated and compared with those of corresponding glial cells infected with
S. suis strain P1/7. The TLR2 gene transcription levels in primary astrocytes and BV2 cells infected with NN1 peaked at 4 h post-infection, whereas the TLR2 gene transcription levels in glial cells infected with BS26 peaked at 8 h post-infection (
Figure 3G,H). Of the two
S. parasuis strains, NN1 induced higher transcription levels of the TLR2 gene in primary astrocytes before 18 h post-infection and in BV2 cells at 4 h post-infection, whereas BS26 induced higher transcription levels of the TLR2 gene in BV2 cells at 8 h and 12 h post-infection (
Figure 3G,H).
At 4 h post-infection, the transcription levels of the TLR2 gene in both primary astrocytes and BV2 cells infected with two
S. parasuis strains were significantly higher than those of cells infected with
S. suis strain P1/7; conversely,
S. suis strain P1/7 induced the highest transcription levels of the TLR2 gene among three strains in primary astrocytes and BV2 cells from 12 h and 8 h post-infection, respectively (
Figure 3G,H).
Neither S. suis strain P1/7 nor the two S. parasuis strains upregulated the transcription levels of the TLR1, 6, and 9 genes in both glial cells.
3.8. The Capacity of S. parasuis Strains to Resist Phagocytosis by BV2 Cells Was Lower Than That of S. suis Strain P1/7 under Various Conditions
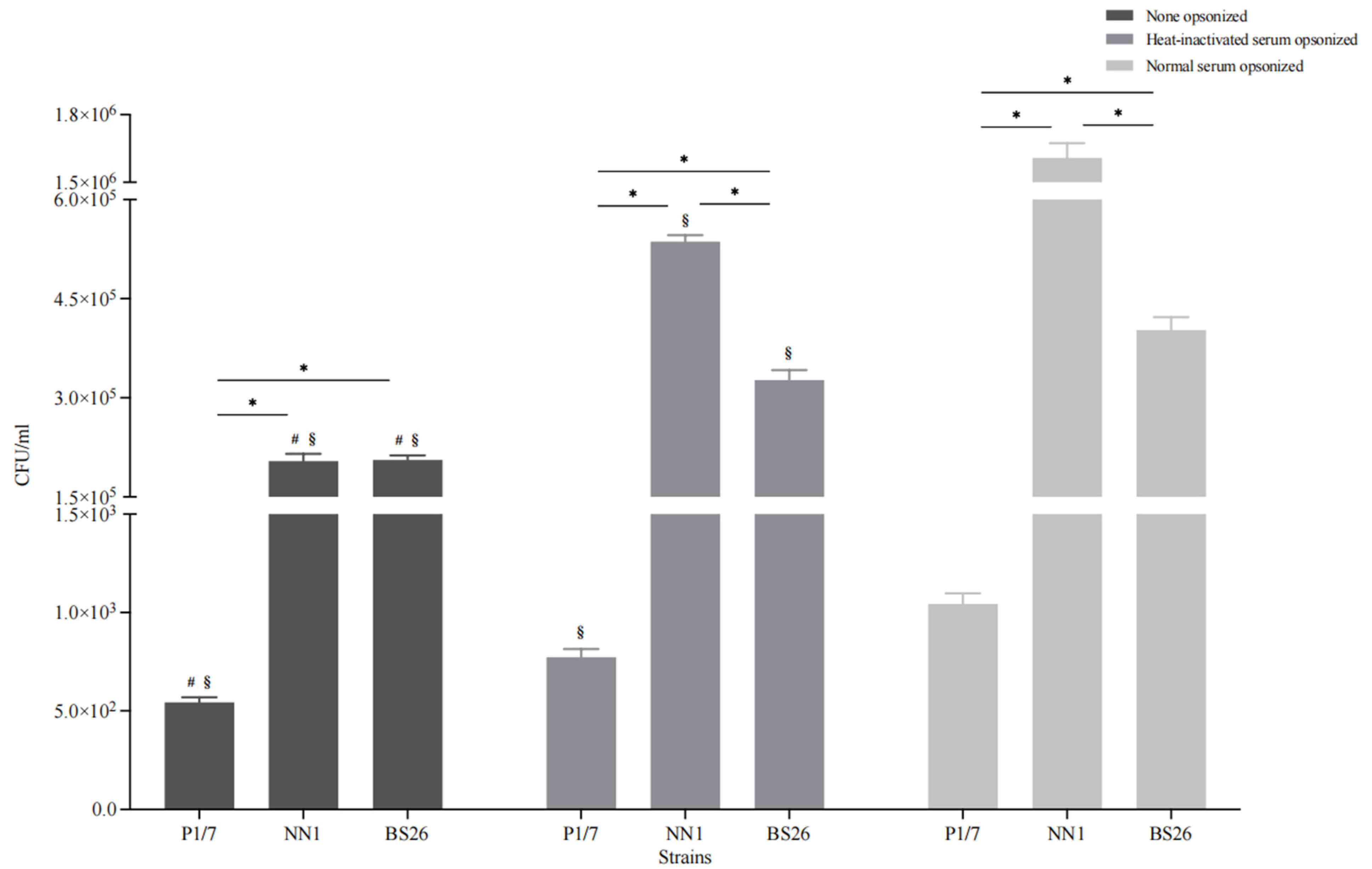
Microglia are phagocytes that play critical roles in the removal of invading pathogens [
22]. Under non-opsonic conditions, the phagocytosis levels of the two
S. parasuis strains (more than 10% of the initial inoculum for both strains) were significantly higher than that of
S. suis strain P1/7 (less than 0.1% of the initial inoculum) (
Figure 4). In addition, the capacity of BS26 to resist phagocytosis by BV2 cells was significantly higher than that of NN1 (
Figure 4).
To evaluate the contribution of the complement pathway to the phagocytosis of
S. suis strain P1/7 and two
S. parasuis strains, bacterial opsonization with normal and heat-inactivated mouse serum was performed prior to the experiment, respectively. Normal serum pre-opsonization resulted in an eightfold increase in the internalization of
S. parasuis strain NN1, whereas the internalization of
S. suis strain P1/7 and
S. parasuis strain BS26 increased by twofold (
Figure 4). Notably, the phagocytosis levels of the three heat-inactivated serum pre-opsonized strains were significantly lower than those of corresponding normal serum pre-opsonized strains, although they were significantly higher than those of corresponding non-opsonized strains (
Figure 4).
4. Discussion
To date, three strains of
S. parasuis, an emerging zoonotic pathogen, have been isolated from the blood cultures of patients with peritonitis, pneumonia, and arthritis. The putative
S. suis virulence-related genes present in
S. parasuis clinical strains were widespread in intermediately and lowly pathogenic
S. suis strains [
23]. Interestingly, none of the
S. parasuis genomes harbored “classical”
S. suis virulence markers
mrp,
sly, and
epf. Virulence genes preferentially present in highly pathogenic and epidemic
S. suis strain, such as
ofs,
sao,
nisK,
nisR,
salK,
salR,
revS,
virB4,
virD4, and
SSU05_0473 genes [
23,
24,
25], were absent in three
S. parasuis clinical strains. It indicated that the pathogenic mechanism of
S. parasuis clinical strains may differ from that of highly pathogenic
S. suis strains. The survival assay revealed substantial differences in virulence levels of the three
S. parasuis clinical strains, even though they exhibited phylogenetic affinity and a nearly identical distribution of putative virulence genes.
A better understanding of the pathogenesis of
S. parasuis is important to establish effective anti-inflammatory strategies. Hitherto, information on the pathogenicity of
S. parasuis and the mechanisms underlying the host inflammatory response initiated by the
S. parasuis infection is still scarce.
S. suis has a phylogenetic affinity with
S. parasuis. Meningitis is the most common clinical syndrome of
S. suis infection in humans [
26]. Evaluation on the potential of
S. parasuis to initiate inflammation in CNS is critically needed. The presence and replication in the CNS are the first steps for pathogens to establish infection. Our previous study demonstrated the capacity of
S. parasuis strains to enter the CNS of infected mice [
7]. However, the cerebral immune response following
S. parasuis infection and its subsequent damage to the CNS tissue remain unclear. In the present study, we investigated the features of the cerebral inflammatory response against
S. parasuis infection in survival mice with neurological symptoms and the roles of resident glial cells in the inflammation.
The two S. parasuis strains induced neurological symptoms in infected C57BL/6 mice from 24 h post-infection. The major neurological symptoms of S. parasuis-infected mice were paresis of the hindlimb, bending of the head toward one side, and walking in circles. It is noteworthy that a higher proportion and more rapid appearance of neurological symptoms were observed in survival mice infected with S. parasuis strain BS26 than those of survival mice infected with S. parasuis strain NN1. Compared with survival mice infected with S. parasuis strains, no neurological symptoms were observed in survival mice infected with S. suis strain P1/7. This may be partially attributable to the high mortality of mice infected with S. suis strain P1/7 from 12 h post-infection.
Various histopathological changes were observed in the brains of mice with neurological symptoms infected with the two S. parasuis strains. At the early phase of infection, the common histopathological changes of mice infected with both S. parasuis strains were neuronal atrophy, neuronal deformation, and vacuolization of the neuronal cytoplasm. However, these histopathological changes were mainly present in the cerebral cortex of mice infected with S. parasuis strain NN1, whereas they appeared in both the hippocampus and cerebral cortex of mice infected with S. parasuis strain BS26. From 24 h post-infection, microgliosis in the cerebral cortex and the engulfment of neuronal debris by microglia were observed in the brains of mice infected with NN1 and BS26, respectively. At the later phase of infection, a higher proportion of hemorrhage and slight neutrophil infiltration was present in the brains of NN1- and BS26-infected mice, respectively. These findings indicate that the two S. parasuis strains induced different histopathological changes in the CNS, and the related pathogenesis of these changes may differ between the two strains.
The rapid and effective initiation of the host immune response is critical for the CNS to clear invading pathogens, but it also results in neurological dysfunction by producing high levels of proinflammatory cytokines and chemokines [
27]. In the present study, the two
S. parasuis strains sequentially induced the upregulation of IL-6, TNF-α, and MCP-1 gene transcription in the brains of mice with neurological symptoms from 24 h post-infection. The significantly higher CSF concentrations of IL-6, TNF-α, and MCP-1 are usually found in the acute phase of patients with bacterial meningitis, especially in patients with a fatal outcome [
28,
29]. This indicates that proinflammatory mediators may play critical roles in the histopathological changes and related neurological symptoms induced by
S. parasuis strains.
The upregulation of IL-6 gene transcription was more rapid than that of the TNF-α and MCP-1 genes in the brains of both
S. parasuis strains infected mice with neurological symptoms. This indicates that IL-6, an important inducer of acute phase proteins, may play a key role in the initiation and amplification of the inflammatory response against invading
S. parasuis strains.
S. parasuis strain NN1 induced a significantly higher transcription level of the MCP-1 gene than
S. parasuis strain BS26 did at 48 h post-infection, whereas no differences in the transcription levels of the TNF-α and IL-6 genes were observed between the two
S. parasuis strains. MCP-1 is responsible for the recruitment of neutrophils and mononuclear cells from the peripheral circulation into the inflammation site of the CNS [
30]. However, no obvious neutrophil infiltration in the brains of
S. parasuis strain NN1-infected mice with neurological symptoms was observed. This may be partially attributable to the drastic decline in the MCP-1 transcription level after reaching the peak.
TLRs play a critical role in inflammation by activating downstream signaling pathways, leading to the expression of a variety of proinflammation cytokines and chemokines [
31]. TLR2 activation was involved in the pathogenesis of meningitis induced by
S. suis strains [
14,
19,
21]. TLR1 and TLR6 binded with TLR2 to form receptor clusters (heterodimers) in response to different G
+ bacterial components [
32]. In the present study, the upregulation of TLR1 and TLR2 gene transcription was observed in the brains of two
S. parasuis strains-infected mice with neurological symptoms.
S. parasuis strains induced a rapid upregulation of TLR1 gene transcription, whereas they induced a comparatively delayed upregulation of TLR2 gene transcription. The delayed expression of TLR2 was also observed in CD1 mice infected with
S. suis ST1 strain [
19]. It is possible that TLR1 played an important role in the initiation of cerebral inflammation induced by the two
S. parasuis strains. Although only TLR1 and TLR2 gene transcription was upregulated, we cannot exclude the possibility that other receptors were involved in the pathogenesis of cerebral inflammation induced by
S. parasuis strains. Further research is needed to investigate the immune pathways exploited by
S. parasuis strains to induce the production of various proinflammation cytokines and chemokines in the CNS.
Resident glial cells play critical roles in the inflammatory response following bacterial invasion of the brain. Previous studies have shown that primary astrocytes and BV2 cells played key roles in the CNS inflammation triggered by
S. suis strains [
14,
20,
21]. In the present study, the two
S. parasuis strains induced higher levels of proinflammatory cytokine and chemokine production by primary astrocytes and BV2 cells compared with
S. suis strain P1/7 at the early phase of interaction. This may partially lead to the high proportion of survival mice infected with two
S. parasuis strains developing neurological symptoms at the early phase of infection. In contrast,
S. suis strain P1/7 induced significantly higher levels of proinflammatory cytokine and chemokine production by primary astrocytes and BV2 cells at the later phase of interaction. No clinical symptoms of meningitis were observed in P1/7-infected mice at the early phase of infection, which may be partially attributed to the delayed production of proinflammatory mediators by astrocytes and microglia.
NN1 possessed a higher capacity to induce proinflammatory cytokine and chemokine production by primary astrocytes throughout the experiment, whereas BS26 induced higher levels of IL-6 production by BV2 cells at the later phase of interaction. We propose that activated astrocytes may play a critical role in the NN1-induced cerebral inflammatory response, whereas the role of microglia may be more crucial in the BS26-induced CNS inflammation.
The rapid production of proinflammatory mediators by astrocytes and BV2 cells which interacted with the two S. parasuis strains may be partially due to the significant upregulation of TLR2 gene transcription at the very early phase of interaction. Moreover, the transcription levels of the TLR2 gene in both primary astrocytes and BV2 cells which interacted with the two S. parasuis strains were significantly higher than those of cells that interacted with S. suis strain P1/7 at the very early phase of interaction. These results underline the importance of TLR2 in mediating S. parasuis-induced cerebral inflammation. Neither S. parasuis strains upregulated TLR1 gene transcription in primary astrocytes or BV2 cells. It is possible that endothelial and ependymal cells, rather than resident microglia and astrocytes, were the main cellular sources of upregulated TLR1 gene transcription in the CNS.
The transience of proinflammatory mediator’s transcription upregulation in the brains of two
S. parasuis strains infected mice with neurological symptoms may be partially due to their low capacity to resist serum-mediated opsonic phagocytosis. Conversely,
S. suis strain P1/7 possessed a significantly higher capacity to resist serum-mediated opsonic phagocytosis than the two
S. parasuis strains did. The bacterial burden and duration in the brains of mice infected with
S. suis strain P1/7 were significantly greater than those of mice infected with the
S. parasuis strains [
7]. The combination of the higher capacity to activate glial cells at the later phase of infection and higher bacterial burden and duration in the brain led to a prolonged and stronger immune response in the brains of mice infected with
S. suis strain P1/7. This is supported by the findings of a previous study, in which severe symptoms of meningitis and histopathological lesions were observed in CD1 mice infected with
S. suis strain ST1 from 4 days to 9 days post-infection [
19].
The complement system is an important element of the innate immune defense against many bacterial pathogens. Our results indicated that the loss of complement activity resulted in a significant reduction in the internalization of
S. parasuis strains. Furthermore, the observed differences in phagocytosis levels between heat-inactivated serum opsonic and non-opsonic conditions indicate that other heat-stable serum factors, such as albumin and fibronectin, are also involved in opsonic phagocytosis of
S. parasuis strains. BS26 possessed a significantly higher capacity to resist serum-mediated opsonic phagocytosis than NN1. Kozel et al. proposed that complement opsonization and capsule size have an inverse relationship with phagocytosis: when the capsule size increases, the efficacy of complement-mediated phagocytosis decreases [
33]. The
cps gene cluster of
S. parasuis strain NN1 shared a 99.2% sequence similarity with
S. parasuis strain BS26. Further studies are needed to investigate the differences in CPS expression in vivo between the two
S. parasuis strains.
In conclusion, S. parasuis clinical strains BS26 and NN1 induced transient and significant innate immunity response in the brain, mainly through the sequential activation of TLR1 and TLR2 at the early phase of infection, which then resulted in histopathological lesions and the appearance of neurological symptoms in mice. TLR2-mediated activation of astrocytes and microglia played important roles in the cerebral inflammatory response induced by BS26 and NN1, respectively. Our data suggest that S. parasuis clinical strains possess a high potential to induce cerebral inflammation in susceptible people at the early phase of infection. Further studies are needed to investigate the differences in the pathogenesis of inducing cerebral inflammation between S. parasuis strains BS26 and NN1.



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}