
Interaction of Microbes with Microplastics and Nanoplastics in the Agroecosystems—Impact on Antimicrobial Resistance



Abstract
:1. Introduction
2. The Physiologic Implications of Microbe–MNP Interactions
2.1. MNP-Induced Gene Expressions and Gene Transfers
2.2. MP-Associated Biofilms and Cluster Formation
3. Influence of MNPs in the Propagation of AMR in Agroecosystem
3.1. Impacts on Resistome
3.1.1. In the Wastewater Treatment System
3.1.2. In the Water Ecosystem
3.1.3. In Soil
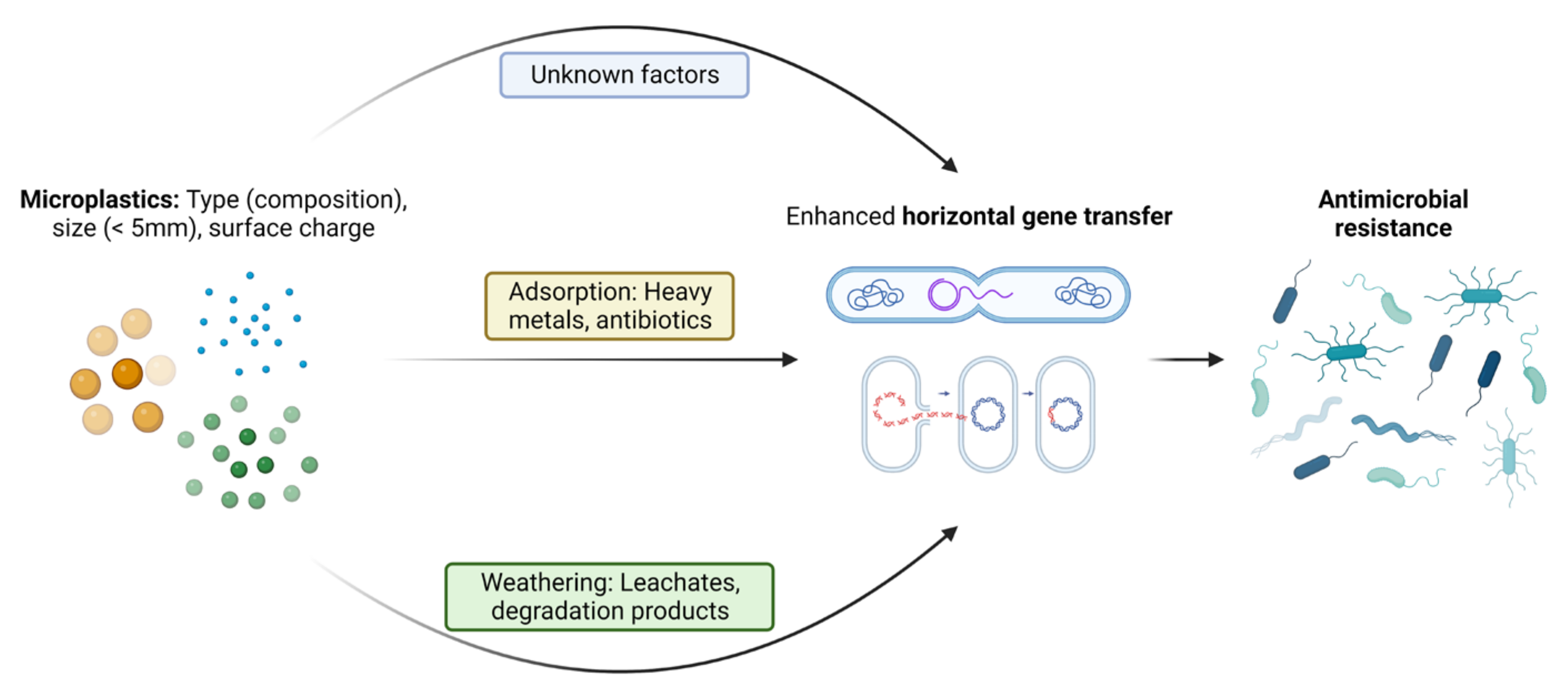
3.2. Impacts on the HGT
3.2.1. Size of MPs
3.2.2. Type and Concentration of MPs
3.2.3. Aging of MPs
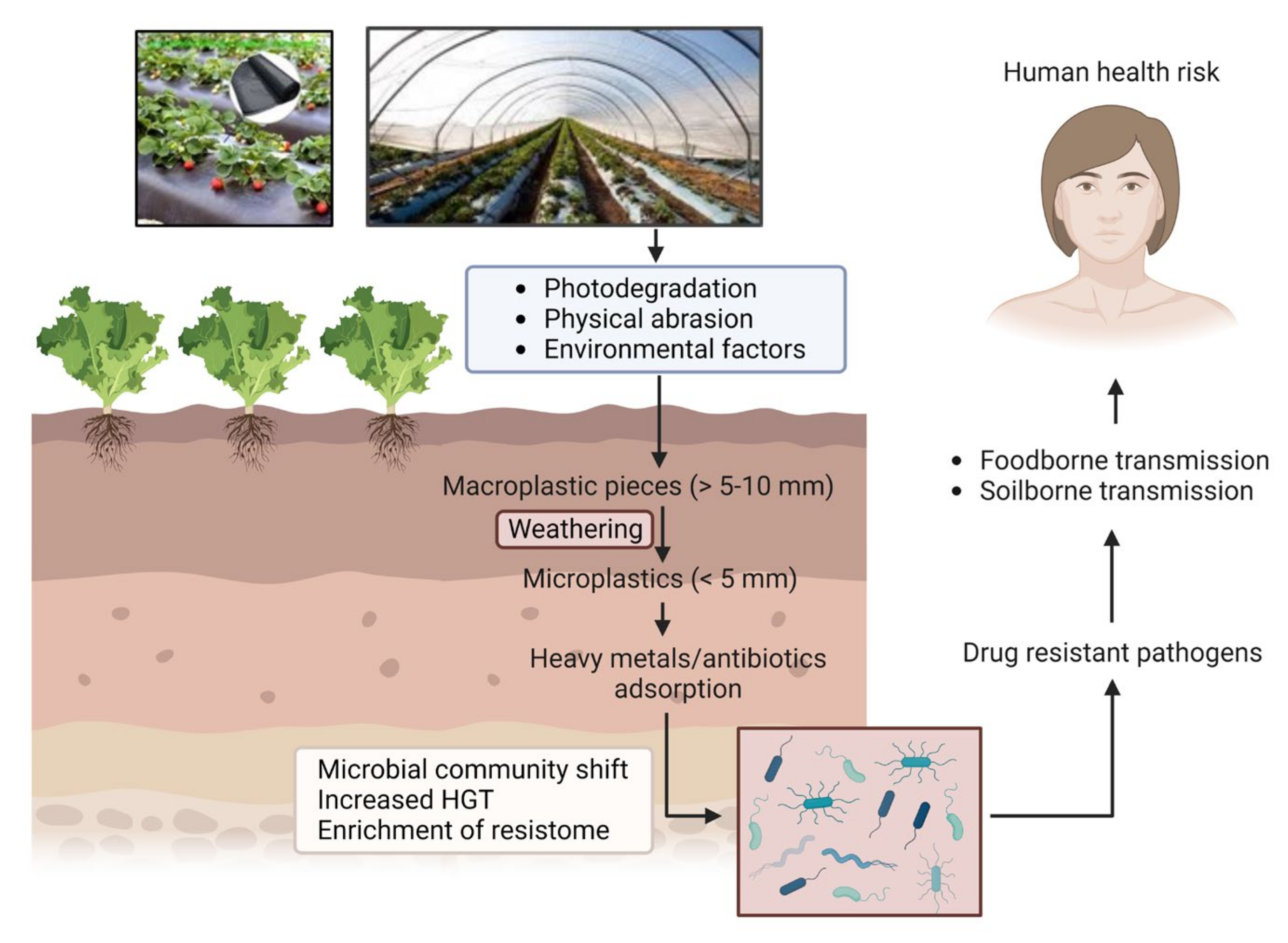
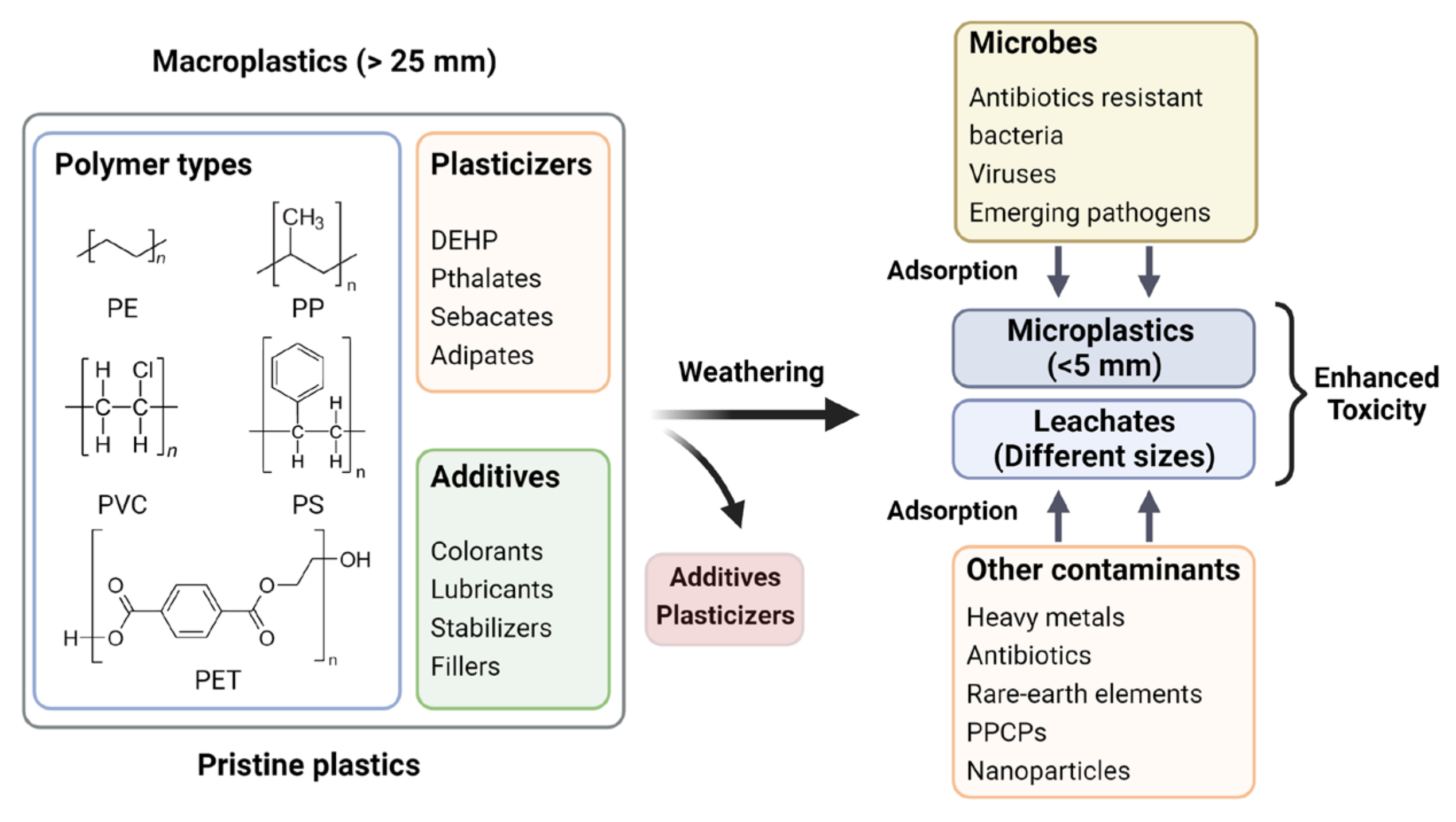
4. Impacts of Long-Term Persistence of MPs on Agroecosystem
Adsorption of Co-Contaminants
5. Rapid Detection, Identification, and Quantification of MPs from Environmental Matrices
5.1. Methodological Challenges
5.2. Recent Developments in Analytical Methods for MNP Detection
6. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef]
- Bråte, I.L.N.; Hurley, R.; Iversen, K.; Beyer, J.; Thomas, K.V.; Steindal, C.C.; Green, N.W.; Olsen, M.; Lusher, A. Mytilus spp. as sentinels for monitoring microplastic pollution in Norwegian coastal waters: A qualitative and quantitative study. Environ. Pollut. 2018, 243, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Digka, N.; Tsangaris, C.; Torre, M.; Anastasopoulou, A.; Zeri, C. Microplastics in mussels and fish from the Northern Ionian Sea. Mar. Pollut. Bull. 2018, 135, 30–40. [Google Scholar] [CrossRef]
- Gedik, K.; Eryaşar, A.R. Microplastic pollution profile of Mediterranean mussels (Mytilus galloprovincialis) collected along the Turkish coasts. Chemosphere 2020, 260, 127570. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, T.; Liu, P.; Li, H.; Hu, F. The formation of specific bacterial communities contributes to the enrichment of antibiotic resistance genes in the soil plastisphere. J. Hazard. Mater. 2022, 436, 129247. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Mi, J.; Yan, Q.; Wen, X.; Zhou, S.; Wang, Y.; Ma, B.; Zou, Y.; Liao, X.; Wu, Y. Animal manures application increases the abundances of antibiotic resistance genes in soil-lettuce system associated with shared bacterial distributions. Sci. Total Environ. 2021, 787, 147667. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.P.; Tsang, C.F.; Li, L.; Yu, R.M.K.; Kong, R.Y.C. Microplastics act as a carrier for wastewater-borne pathogenic bacteria in sewage. Chemosphere 2022, 301, 134692. [Google Scholar] [CrossRef]
- Dai, Y.; Shi, J.; Zhang, N.; Pan, Z.; Xing, C.; Chen, X. Current research trends on microplastics pollution and impacts on agro-ecosystems: A short review. Sep. Sci. Technol. 2022, 57, 656–669. [Google Scholar] [CrossRef]
- Isari, E.A.; Papaioannou, D.; Kalavrouziotis, I.K.; Karapanagioti, H.K. Microplastics in Agricultural Soils: A Case Study in Cultivation of Watermelons and Canning Tomatoes. Water 2021, 13, 2168. [Google Scholar] [CrossRef]
- Lang, M.; Wang, G.; Yang, Y.; Zhu, W.; Zhang, Y.; Ouyang, Z.; Guo, X. The occurrence and effect of altitude on microplastics distribution in agricultural soils of Qinghai Province, northwest China. Sci. Total Environ. 2022, 810, 152174. [Google Scholar] [CrossRef]
- Castan, S.; Henkel, C.; Hüffer, T.; Hofmann, T. Microplastics and nanoplastics barely enhance contaminant mobility in agricultural soils. Commun. Earth Environ. 2021, 2, 193. [Google Scholar] [CrossRef]
- Helmberger, M.S.; Tiemann, L.K.; Grieshop, M.J. Towards an ecology of soil microplastics. Funct. Ecol. 2020, 34, 550–560. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Lozano, Y.M.; Rillig, M.C. Microplastics increase soil pH and decrease microbial activities as a function of microplastic shape, polymer type, and exposure time. Front. Environ. Sci. 2021, 9, 675803. [Google Scholar] [CrossRef]
- Ding, L.; Huang, D.; Ouyang, Z.; Guo, X. The effects of microplastics on soil ecosystem: A review. Curr. Opin. Environ. Sci. Health 2022, 26, 100344. [Google Scholar] [CrossRef]
- Joos, L.; De Tender, C. Soil under stress: The importance of soil life and how it is influenced by (micro) plastic pollution. Comput. Struct. Biotechnol. J. 2022, 20, 1554–1566. [Google Scholar] [CrossRef]
- Tian, L.; Jinjin, C.; Ji, R.; Ma, Y.; Yu, X. Microplastics in agricultural soils: Sources, effects, and their fate. Curr. Opin. Environ. Sci. Health 2022, 25, 100311. [Google Scholar] [CrossRef]
- Maity, S.; Guchhait, R.; Sarkar, M.B.; Pramanick, K. Occurrence and distribution of micro/nanoplastics in soils and their phytotoxic effects: A review. Plant Cell Environ. 2022, 45, 1011–1028. [Google Scholar] [CrossRef]
- He, F.; Shi, H.; Liu, R.; Tian, G.; Qi, Y.; Wang, T. Randomly-shaped nanoplastics induced stronger biotoxicity targeted to earthworm Eisenia fetida species: Differential effects and the underlying mechanisms of realistic and commercial polystyrene nanoplastics. Sci. Total Environ. 2023, 877, 162854. [Google Scholar] [CrossRef]
- Pazienza, P.; De Lucia, C. For a new plastics economy in agriculture: Policy reflections on the EU strategy from a local perspective. J. Clean. Prod. 2020, 253, 119844. [Google Scholar] [CrossRef]
- Scarascia-Mugnozza, G.; Sica, C.; Russo, G. Plastic materials in European agriculture: Actual use and perspectives. J. Agric. Eng. 2011, 42, 15–28. [Google Scholar] [CrossRef]
- Battulga, B.; Kawahigashi, M.; Oyuntsetseg, B. Characterization of biofilms formed on polystyrene microplastics (PS-MPs) on the shore of the Tuul River, Mongolia. Environ. Res. 2022, 212, 113329. [Google Scholar] [CrossRef] [PubMed]
- Sagawa, N.; Kawaai, K.; Hinata, H. Abundance and size of microplastics in a coastal sea: Comparison among bottom sediment, beach sediment, and surface water. Mar. Pollut. Bull. 2018, 133, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Ma, J.; Li, G.; Rillig, M.C.; Zhu, Y.-G. Soil plastispheres as hotspots of antibiotic resistance genes and potential pathogens. ISME J. 2022, 16, 521–532. [Google Scholar] [CrossRef]
- Xu, B.; Liu, F.; Cryder, Z.; Huang, D.; Lu, Z.; He, Y.; Wang, H.; Lu, Z.; Brookes, P.C.; Tang, C.; et al. Microplastics in the soil environment: Occurrence, risks, interactions and fate–a review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2175–2222. [Google Scholar] [CrossRef]
- Rillig, M.C.; Leifheit, E.; Lehmann, J. Microplastic effects on carbon cycling processes in soils. PLoS Biol. 2021, 19, e3001130. [Google Scholar] [CrossRef] [PubMed]
- Junaid, M.; Liu, X.; Wu, Y.; Wang, J. Selective enrichment of antibiotic resistome and bacterial pathogens by aquatic microplastics. J. Hazard. Mater. Adv. 2022, 7, 100106. [Google Scholar] [CrossRef]
- Wu, X.; Pan, J.; Li, M.; Li, Y.; Bartlam, M.; Wang, Y. Selective enrichment of bacterial pathogens by microplastic biofilm. Water Res. 2019, 165, 114979. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, J.; Li, W. Conjugative antibiotic-resistant plasmids promote bacterial colonization of microplastics in water environments. J. Hazard. Mater. 2022, 430, 128443. [Google Scholar] [CrossRef]
- Imran, M.; Das, K.R.; Naik, M.M. Co-selection of multi-antibiotic resistance in bacterial pathogens in metal and microplastic contaminated environments: An emerging health threat. Chemosphere 2019, 215, 846–857. [Google Scholar] [CrossRef]
- Yuan, Q.; Sun, R.; Yu, P.; Cheng, Y.; Wu, W.; Bao, J.; Alvarez, P.J. UV-aging of microplastics increases proximal ARG donor-recipient adsorption and leaching of chemicals that synergistically enhance antibiotic resistance propagation. J. Hazard. Mater. 2022, 427, 127895. [Google Scholar] [CrossRef]
- Zhao, T.; Tan, L.; Han, X.; Wang, X.; Zhang, Y.; Ma, X.; Lin, K.; Wang, R.; Ni, Z.; Wang, J.; et al. Microplastic-induced apoptosis and metabolism responses in marine Dinoflagellate, Karenia mikimotoi. Sci. Total Environ. 2022, 804, 150252. [Google Scholar] [CrossRef]
- Feng, L.-J.; Sun, X.-D.; Zhu, F.-P.; Feng, Y.; Duan, J.-L.; Xiao, F.; Li, X.-Y.; Shi, Y.; Wang, Q.; Sun, J.-W.; et al. Nanoplastics promote microcystin synthesis and release from cyanobacterial Microcystis aeruginosa. Environ. Sci. Technol. 2020, 54, 3386–3394. [Google Scholar] [CrossRef] [PubMed]
- Zha, Y.; Li, Z.; Zhong, Z.; Ruan, Y.; Sun, L.; Zuo, F.; Li, L.; Hou, S. Size-dependent enhancement on conjugative transfer of antibiotic resistance genes by micro/nanoplastics. J. Hazard. Mater. 2022, 431, 128561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-J.; Zeng, Y.-H.; Zhu, J.-M.; Cai, Z.-H.; Zhou, J. The structure and assembly mechanisms of plastisphere microbial community in natural marine environment. J. Hazard. Mater. 2022, 421, 126780. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, J.; Dai, H.; Zhao, Y.; Li, D.; Duan, W.; Guo, Y. Microplastics affect the ammonia oxidation performance of aerobic granular sludge and enrich the intracellular and extracellular antibiotic resistance genes. J. Hazard. Mater. 2021, 409, 124981. [Google Scholar] [CrossRef]
- Huang, H.; Feng, G.; Wang, M.; Liu, C.; Wu, Y.; Dong, L.; Feng, L.; Zheng, X.; Chen, Y. Nitric oxide: A neglected driver for the conjugative transfer of antibiotic resistance genes among wastewater microbiota. Environ. Sci. Technol. 2022, 56, 6466–6478. [Google Scholar] [CrossRef]
- Lenaker, P.L.; Corsi, S.R.; Mason, S.A. Spatial distribution of microplastics in surficial benthic sediment of Lake Michigan and Lake Erie. Environ. Sci. Technol. 2020, 55, 373–384. [Google Scholar] [CrossRef]
- Steinman, A.D.; Scott, J.; Green, L.; Partridge, C.; Oudsema, M.; Hassett, M.; Kindervater, E.; Rediske, R.R. Persistent organic pollutants, metals, and the bacterial community composition associated with microplastics in Muskegon Lake (MI). J. Great Lakes Res. 2020, 46, 1444–1458. [Google Scholar] [CrossRef]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef]
- Lami, R. Quorum sensing in marine biofilms and environments. In Quorum Sensing; Academic Press: Cambridge, MA, USA, 2019; pp. 55–96. [Google Scholar] [CrossRef]
- Liu, S.; Shi, J.; Wang, J.; Dai, Y.; Li, H.; Li, J.; Liu, X.; Chen, X.; Wang, Z.; Zhang, P. Interactions Between Microplastics and Heavy Metals in Aquatic Environments: A Review. Front. Microbiol. 2021, 12, 652520. [Google Scholar] [CrossRef]
- Cai, L.; Wu, D.; Xia, J.; Shi, H.; Kim, H. Influence of physicochemical surface properties on the adhesion of bacteria onto four types of plastics. Sci. Total Environ. 2019, 671, 1101–1107. [Google Scholar] [CrossRef]
- Xu, X.; Wang, S.; Gao, F.; Li, J.; Zheng, L.; Sun, C.; He, C.; Wang, Z.; Qu, L. Marine microplastic-associated bacterial community succession in response to geography, exposure time, and plastic type in China’s coastal seawaters. Mar. Pollut. Bull. 2019, 145, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Reddy, S.; Barathe, P.; Oak, U.; Shriram, V.; Kharat, S.S.; Govarthanan, M.; Kumar, V. Microplastic-associated pathogens and antimicrobial resistance in environment. Chemosphere 2021, 291, 133005. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Z.; Li, M.; Bartlam, M.; Wang, Y. Integrated metagenomic and metatranscriptomic analysis reveals actively expressed antibiotic resistomes in the plastisphere. J. Hazard. Mater. 2022, 430, 128418. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cao, N.; Duan, C.; Wang, Q.; Ding, C.; Wang, J. Selection of antibiotic resistance genes on biodegradable and non-biodegradable microplastics. J. Hazard. Mater. 2021, 409, 124979. [Google Scholar] [CrossRef]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot spots of horizontal gene transfer (HGT) in aquatic environments, with a focus on a new HGT mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Zhou, C.-S.; Wu, J.-W.; Liu, B.-F.; Ma, W.-L.; Yang, S.-S.; Cao, G.-L. (Micro) nanoplastics promote the risk of antibiotic resistance gene propagation in biological phosphorus removal system. J. Hazard. Mater. 2022, 431, 128547. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Chen, Y.; Meng, X.; Dieketseng, M.Y.; Wang, X.; Yan, S.; Wang, B.; Zhou, L.; Zheng, G. A neglected risk of nanoplastics as revealed by the promoted transformation of plasmid-borne ampicillin resistance gene by Escherichia coli. Environ. Microbiol. 2022, 24, 4946–4959. [Google Scholar] [CrossRef]
- Li, R.; Zhu, L.; Yang, K.; Li, H.; Zhu, Y.-G.; Cui, L. Impact of Urbanization on Antibiotic Resistome in Different Microplastics: Evidence from a Large-Scale Whole River Analysis. Environ. Sci. Technol. 2021, 55, 8760–8770. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, J.; Wu, J.; Wang, J.; Luo, Y. Potential risks of microplastics combined with superbugs: Enrichment of antibiotic resistant bacteria on the surface of microplastics in mariculture system. Ecotoxicol. Environ. Saf. 2020, 187, 109852. [Google Scholar] [CrossRef] [PubMed]
- Galafassi, S.; Sabatino, R.; Sathicq, M.B.; Eckert, E.M.; Fontaneto, D.; Fontana, G.D.; Mossotti, R.; Corno, G.; Volta, P.; Di Cesare, A. Contribution of microplastic particles to the spread of resistances and pathogenic bacteria in treated wastewaters. Water Res. 2021, 201, 117368. [Google Scholar] [CrossRef]
- Xu, C.; Lu, J.; Shen, C.; Wang, J.; Li, F. Deciphering the mechanisms shaping the plastisphere antibiotic resistome on riverine microplastics. Water Res. 2022, 225, 119192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, J.; Zhao, Y.; Dai, H.; Jia, J.; Zhang, D. Plastisphere enrich antibiotic resistance genes and potential pathogenic bacteria in sewage with pharmaceuticals. Sci. Total Environ. 2021, 768, 144663. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, Q.; Zhao, S.; Zhao, P.; Yang, X.; Huang, Q.; Su, J. Watershed urbanization enhances the enrichment of pathogenic bacteria and antibiotic resistance genes on microplastics in the water environment. Environ. Pollut. 2022, 313, 120185. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.-M.; Lu, P.-Z.; Liu, X.-P. Fate and abundance of antibiotic resistance genes on microplastics in facility vegetable soil. Sci. Total Environ. 2020, 709, 136276. [Google Scholar] [CrossRef] [PubMed]
- Basili, M.; Quero, G.M.; Giovannelli, D.; Manini, E.; Vignaroli, C.; Avio, C.G.; De Marco, R.; Luna, G.M. Major role of surrounding environment in shaping biofilm community composition on marine plastic debris. Front. Mar. Sci. 2020, 7, 262. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, Z.; Zhu, J.; Shi, J.; Wei, H.; Xie, B.; Shi, H. Microplastics act as vectors for antibiotic resistance genes in landfill leachate: The enhanced roles of the long-term aging process. Environ. Pollut. 2021, 270, 116278. [Google Scholar] [CrossRef]
- Zhao, R.; Yu, K.; Zhang, J.; Zhang, G.; Huang, J.; Ma, L.; Deng, C.; Li, X.; Li, B. Deciphering the mobility and bacterial hosts of antibiotic resistance genes under antibiotic selection pressure by metagenomic assembly and binning approaches. Water Res. 2020, 186, 116318. [Google Scholar] [CrossRef]
- Wiedner, K.; Polifka, S. Effects of microplastic and microglass particles on soil microbial community structure in an arable soil (Chernozem). Soil 2020, 6, 315–324. [Google Scholar] [CrossRef]
- Bosker, T.; Bouwman, L.J.; Brun, N.R.; Behrens, P.; Vijver, M.G. Microplastics accumulate on pores in seed capsule and delay germination and root growth of the terrestrial vascular plant Lepidium sativum. Chemosphere 2019, 226, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Hu, G.; Fan, X.; Jia, H. Sorption properties of cadmium on microplastics: The common practice experiment and A two-dimensional correlation spectroscopic study. Ecotoxicol. Environ. Saf. 2020, 190, 110118. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yang, C.; Zhu, Z.; Bai, X.; Ma, J. Adsorption behavior of organic pollutants and metals on micro/nanoplastics in the aquatic environment. Sci. Total Environ. 2019, 694, 133643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, J.; Zhou, B.; Zhou, Y.; Dai, Z.; Zhou, Q.; Christie, P.; Luo, Y. Enhanced adsorption of oxytetracycline to weathered microplastic polystyrene: Kinetics, isotherms and influencing factors. Environ. Pollut. 2018, 243 Pt B, 1550–1557. [Google Scholar] [CrossRef]
- Fan, X.; Zou, Y.; Geng, N.; Liu, J.; Hou, J.; Li, D.; Yang, C.; Li, Y. Investigation on the adsorption and desorption behaviors of antibiotics by degradable MPs with or without UV ageing process. J. Hazard. Mater. 2021, 401, 123363. [Google Scholar] [CrossRef]
- Ho, W.-K.; Law, J.C.-F.; Lo, J.C.-W.; Chng, I.K.-X.; Hor, C.H.-H.; Leung, K.S.-Y. Sorption Behavior, Speciation, and Toxicity of Microplastic-Bound Chromium in Multisolute Systems. Environ. Sci. Technol. Lett. 2023, 10, 27–32. [Google Scholar] [CrossRef]
- Pompei, C.M.E.; Campos, L.C.; da Silva, B.F.; Fogo, J.C.; Vieira, E.M. Occurrence of PPCPs in a Brazilian water reservoir and their removal efficiency by ecological filtration. Chemosphere 2019, 226, 210–219. [Google Scholar] [CrossRef]
- Fang, S.; Yu, W.; Li, C.; Liu, Y.; Qiu, J.; Kong, F. Adsorption behavior of three triazole fungicides on polystyrene microplastics. Sci. Total Environ. 2019, 691, 1119–1126. [Google Scholar] [CrossRef]
- Zarfl, C.; Matthies, M. Are marine plastic particles transport vectors for organic pollutants to the Arctic? Mar. Pollut. Bull. 2010, 60, 1810–1814. [Google Scholar] [CrossRef]
- Elizalde-Velázquez, A.; Subbiah, S.; Anderson, T.A.; Green, M.J.; Zhao, X.; Cañas-Carrell, J.E. Sorption of three common nonsteroidal anti-inflammatory drugs (NSAIDs) to microplastics. Sci. Total Environ. 2020, 715, 136974. [Google Scholar] [CrossRef]
- Yamashita, R.; Tanaka, K.; Yeo, B.G.; Takada, H.; van Franeker, J.A.; Dalton, M.; Dale, E. Hazardous Chemicals in Plastics in Marine Environments: International Pellet Watch; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Puckowski, A.; Cwięk, W.; Mioduszewska, K.; Stepnowski, P.; Białk-Bielińska, A. Sorption of pharmaceuticals on the surface of microplastics. Chemosphere 2021, 263, 127976. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, M.; Sha, W.; Wang, Y.; Hao, H.; Dou, Y.; Li, Y. Sorption Behavior and Mechanisms of Organic Contaminants to Nano and Microplastics. Molecules 2020, 25, 1827. [Google Scholar] [CrossRef] [Green Version]
- Xiao, F.; Zhang, X.; Penn, L.; Gulliver, J.S.; Simcik, M.F. Effects of Monovalent Cations on the Competitive Adsorption of Perfluoroalkyl Acids by Kaolinite: Experimental Studies and Modeling. Environ. Sci. Technol. 2011, 45, 10028–10035. [Google Scholar] [CrossRef]
- Bundschuh, M.; Weyers, A.; Ebeling, M.; Elsaesser, D.; Schulz, R. Narrow pH Range of Surface Water Bodies Receiving Pesticide Input in Europe. Bull. Environ. Contam. Toxicol. 2016, 96, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.V.; Carter, L.J.; Agatz, A.; Boxall, A.B. Novel Approach for Characterizing pH-Dependent Uptake of Ionizable Chemicals in Aquatic Organisms. Environ. Sci. Technol. 2017, 51, 6965–6971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Zheng, M.; Wang, L.; Zhao, Q.; Lou, Y.; Shi, L.; Qu, L. Sorption of polyhalogenated carbazoles (PHCs) to microplastics. Mar. Pollut. Bull. 2019, 146, 718–728. [Google Scholar] [CrossRef]
- Xu, B.; Liu, F.; Brookes, P.C.; Xu, J. The sorption kinetics and isotherms of sulfamethoxazole with polyethylene microplastics. Mar. Pollut. Bull. 2018, 131, 191–196. [Google Scholar] [CrossRef]
- Joo, S.H.; Liang, Y.; Kim, M.; Byun, J.; Choi, H. Microplastics with adsorbed contaminants: Mechanisms and Treatment. Environ. Chall. 2021, 3, 100042. [Google Scholar] [CrossRef]
- Zhao, L.; Rong, L.; Xu, J.; Lian, J.; Wang, L.; Sun, H. Sorption of five organic compounds by polar and nonpolar microplastics. Chemosphere 2020, 257, 127206. [Google Scholar] [CrossRef]
- Seidensticker, S.; Grathwohl, P.; Lamprecht, J.; Zarfl, C. A combined experimental and modeling study to evaluate pH-dependent sorption of polar and non-polar compounds to polyethylene and polystyrene microplastics. Environ. Sci. Eur. 2018, 30, 30. [Google Scholar] [CrossRef]
- Mári, Á.; Bordós, G.; Gergely, S.; Büki, M.; Háhn, J.; Palotai, Z.; Besenyő, G.; Szabó, É.; Salgó, A.; Kriszt, B.; et al. Validation of microplastic sample preparation method for freshwater samples. Water Res. 2021, 202, 117409. [Google Scholar] [CrossRef]
- Grause, G.; Kuniyasu, Y.; Chien, M.-F.; Inoue, C. Separation of microplastic from soil by centrifugation and its application to agricultural soil. Chemosphere 2022, 288, 132654. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, N.; Anquetil, C.; Dris, R.; Gasperi, J.; Tassin, B.; Derenne, S. Quantification of microplastics by pyrolysis coupled with gas chromatography and mass spectrometry in sediments: Challenges and implications. Microplastics 2022, 1, 229–239. [Google Scholar] [CrossRef]
- De Frond, H.; Hampton, L.T.; Kotar, S.; Gesulga, K.; Matuch, C.; Lao, W.; Weisberg, S.B.; Wong, C.S.; Rochman, C.M. Monitoring microplastics in drinking water: An interlaboratory study to inform effective methods for quantifying and characterizing microplastics. Chemosphere 2022, 298, 134282. [Google Scholar] [CrossRef]
- Lv, L.; He, L.; Jiang, S.; Chen, J.; Zhou, C.; Qu, J.; Lu, Y.; Hong, P.; Sun, S.; Li, C. In situ surface-enhanced Raman spectroscopy for detecting microplastics and nanoplastics in aquatic environments. Sci. Total Environ. 2020, 728, 138449. [Google Scholar] [CrossRef]
- Mowla, M.; Shakiba, S.; Louie, S.M. Selective quantification of nanoplastics in environmental matrices by asymmetric flow field-flow fractionation with total organic carbon detection. Chem. Commun. 2021, 57, 12940–12943. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, R.; Tsuchiya, M.; Lindsay, D.J.; Kitahashi, T.; Fujikura, K.; Fukushima, T. A new small device made of glass for separating microplastics from marine and freshwater sediments. PeerJ 2019, 7, e7915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoffo, E.D.; Ribeiro, F.; O’Brien, J.W.; O’Brien, S.; Tscharke, B.J.; Gallen, M.; Samanipour, S.; Mueller, J.F.; Thomas, K.V. Identification and quantification of selected plastics in biosolids by pressurized liquid extraction combined with double-shot pyrolysis gas chromatography–mass spectrometry. Sci. Total Environ. 2020, 715, 136924. [Google Scholar] [CrossRef]
- Primpke, S.; Fischer, M.; Lorenz, C.; Gerdts, G.; Scholz-Böttcher, B.M. Comparison of pyrolysis gas chromatography/mass spectrometry and hyperspectral FTIR imaging spectroscopy for the analysis of microplastics. Anal. Bioanal. Chem. 2020, 412, 8283–8298. [Google Scholar] [CrossRef]
- Saliu, F.; Biale, G.; Raguso, C.; La Nasa, J.; Degano, I.; Seveso, D.; Galli, P.; Lasagni, M.; Modugno, F. Detection of plastic particles in marine sponges by a combined infrared micro-spectroscopy and pyrolysis-gas chromatography-mass spectrometry approach. Sci. Total Environ. 2022, 819, 152965. [Google Scholar] [CrossRef]
- Sullivan, G.L.; Gallardo, J.D.; Jones, E.W.; Hollliman, P.J.; Watson, T.M.; Sarp, S. Detection of trace sub-micron (nano) plastics in water samples using pyrolysis-gas chromatography time of flight mass spectrometry (PY-GCToF). Chemosphere 2020, 249, 126179. [Google Scholar] [CrossRef]
- Weckhuysen, B.; ten Have, I.; Meirer, F.; Oord, R.; Zettler, E.; Van Sebille, E.; Amaral-Zettler, L. Nanoscale Infrared Spectroscopy Reveals Nanoplastics at 5000 m Depth in the South Atlantic Ocean. Phys. Sci. 2021; preprint. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Y.; Qiu, Z.; He, Y.; Zhang, Y. Towards a fast and generalized microplastic quantification method in soil using terahertz spectroscopy. Sci. Total Environ. 2022, 841, 156624. [Google Scholar] [CrossRef]
- Urso, M.; Ussia, M.; Novotný, F.; Pumera, M. Trapping and detecting nanoplastics by MXene-derived oxide microrobots. Nat. Commun. 2022, 13, 3573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, Y.; Ouyang, X.; Weng, L.; Ma, J.; Islam, S.; Li, Y. Resolving natural organic matter and nanoplastics in binary or ternary systems via UV–Vis analysis. J. Colloid Interface Sci. 2023, 632, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.-S.; Jeong, E.T.; Lee, S.; Kim, I.S.; Chung, S.; Han, S.; Choi, I.; Ryu, Y.-S. Real-Time Underwater Nanoplastic Detection beyond the Diffusion Limit and Low Raman Scattering Cross-Section via Electro-Photonic Tweezers. ACS Nano 2022, 17, 2114–2123. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Skoczynska, E.; van Putten, R.-J.; Leslie, H.A.; Gruter, G.-J.M. Quantification of polyethylene terephthalate micro-and nanoplastics in domestic wastewater using a simple three-step method. Sci. Total Environ. 2023, 857, 159209. [Google Scholar] [CrossRef]
- Moraz, A.; Breider, F. Detection and Quantification of Nonlabeled Polystyrene Nanoparticles Using a Fluorescent Molecular Rotor. Anal. Chem. 2021, 93, 14976–14984. [Google Scholar] [CrossRef]
- Ivleva, N.P. Chemical analysis of microplastics and nanoplastics: Challenges, advanced methods, and perspectives. Chem. Rev. 2021, 121, 11886–11936. [Google Scholar] [CrossRef]
- Jia, W.; Karapetrova, A.; Zhang, M.; Xu, L.; Li, K.; Huang, M.; Wang, J.; Huang, Y. Automated identification and quantification of invisible microplastics in agricultural soils. Sci. Total Environ. 2022, 844, 156853. [Google Scholar] [CrossRef]
- Prata, J.C.; da Costa, J.P.; Girão, A.V.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Identifying a quick and efficient method of removing organic matter without damaging microplastic samples. Sci. Total Environ. 2019, 686, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Amin, A.; Gibson, C.T.; Chuah, C.; Tang, Y.; Naidu, R.; Fang, C. Raman imaging of microplastics and nanoplastics generated by cutting PVC pipe. Environ. Pollut. 2022, 298, 118857. [Google Scholar] [CrossRef] [PubMed]
- Cella, C.; La Spina, R.; Mehn, D.; Fumagalli, F.; Ceccone, G.; Valsesia, A.; Gilliland, D. Detecting micro-and nanoplastics released from food packaging: Challenges and analytical strategies. Polymers 2022, 14, 1238. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, J.; Yu, J.; Zhou, Q.; Wu, W.; Fu, L.; Yin, C.; Fernandez, C.; Karimi-Maleh, H. Current development and future challenges in microplastic detection techniques: A bibliometrics-based analysis and review. Sci. Prog. 2022, 105, 00368504221132151. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protective cultivation films: covering vineyards/orchards direct covering greenhouses and tunnels high and low tunnels mulching nursery films | Piping, irrigation/drainage: channel lining drainage pipes drippers irrigation tapes and pipes micro irrigation water reservoir | Nets: anti-bird anti-hail nets for olives/nuts harvesting shading wind-breaking |
| Other films: fumigation films silage films | Packaging: fertilizers sacks agro-chemical cans/bottles containers tanks for liquid storage crates | Other: baling twines baling wraps nursery pots strings and ropes |
| Type of MPs | Model Bacteria | Research Setup or Test Matrices | Possible Mechanisms | Reference |
|---|---|---|---|---|
| Polystyrene (PS-MPs/PS-NPs) | E. coli DH5α (recipient) plasmid pUC19 (ampR) | Transformation of plasmid borne ARG into recipient bacteria E. coli DH5α, in co-presence of humic acid or Fe3+. | NPs (10–500 mg/L) significantly enhanced the transformation efficiency. NPs induced ROS overproduction, activated SOS response, increased cell membrane permeability, and changed the secretion systems, thereby facilitating the uptake of exogenous DNA by bacteria. | [50] |
| Polystyrene (PS-MPs) | E. coli MG1655 (recipient) E. coli DH5α | Effects of Pristine PS-MPs and UV-irradiated PS-MPs on HGT. Differential gene expression analysis of donor bacteria for conjugation regulating genes, recipient bacteria for genes regulating bacterial sensitivity to phage lambda and genes associated with intracellular ROS production | UV-aged MPs enhanced horizontal ARG transfer compared to pristine MPs. Conjugation was the dominant mechanism. MPs interact via hydrophobic attraction and π-π stacking. Aged MPs upregulated genes regulating conjugation, associated with cell permeability, and intracellular ROS production | [30] |
| Polystyrene (PS) MPs | E. coli DH5α (RP4 plasmid) (Donor) E. coli K12 MG1655 (recipient) | Effect of MNPs size and concentration on HGT. ROS production, cell membrane permeability and viability assay. Analysis of gene expression upon exposure to MPs, through qPCR | The gene transfer efficiency depends on mating time, ratio of bacteria and size, concentration of MNPs. MNPs increased ROS production and cell membrane permeability of the donor and the recipient. NPs at concentration 100 mg/L led to excessive ROS effecting viability of bacteria. | [33] |
| Polystyrene (PS) MPs | Phosphorus removing bacteria | Lab-scale batch reactors (working volume 1 L) operated at 20 °C, for 30 days containing different concentrations of MPs. After of MPs exposure, microbial communities explored by high-throughput sequencing. Quantification of ARGs also performed. | MP/NPs promoted the propagation of ARGs in biological phosphorus removal system. Microbial community shift rather than HGT was the main factor promoting ARG propagation under MP/NP pressure. | [49] |
| Polyvinyl chloride (PVC), Polyamide (PA), Polyethylene (PE), Polystyrene (PS) MP beads (30 μm) | Soil microbial communities | Soil microcosm experiments, to study the effects of manure, temperature, and moisture on microbiome and antibiotic resistome. Amplicon sequencing and HT-qPCR to detect ARGs, and the bacterial community of plastispheres. Field experiment with metagenomic sequencing to study antibiotic resistome of different plastispheres. | The MPs select for microbial communities in the plastisphere (depending on type and size), and these microbiota are involved in a variety of ecological processes in the soil ecosystem. Plastispheres represents a hotspot of increased potential for the HGT. | [23] |
| Polystyrene (PS MPs (diameter 3 mm, height 4 mm). | E. coli HB101 (plasmid RP4) (Donor) E. coli NK5449 resistant to rifampicin and Nalidixic acid (Recipient) | Two types of representative materials, nanoalumina (nanomaterial, conjugative pili promoter) and FNA (conjugative pili inhibitor), were used to verify effect on HGT. Assay of intracellular ATP concentration, and bacterial colonization in MPs using SEM. | RP4 plasmid promotes bacterial colonization on MPs, promoting biofilm development. Negligible effect of donor bacteria alone, on bacterial colonization. When donors and recipient coexist, intracellular ATP concentration and intracellular energy supply increase, simultaneously increasing the expression level of conjugative pili synthesis genes. | [8] |
| Low density polyethylene (LDPE), Polypropylene (PP), Polystyrene (PS) | Soil bacteria | Soil microcosm experiment with 100 g soil in sterilized 350 mL glass jar (incubated at 25 °C for 14 days to activate the microbiota). Three types of MPs (sizes 75 and 550 µm) were added at 2% concentration, incubation for 90 days. DNA was extracted from 0.5 g soil after experiment and 0.2 g of extracted MPs from soil, sequenced and aligned against the Comprehensive Antibiotic Resistance Database for ARG identification | Total abundance of different ARGs was significantly higher in the plastispheres compared to the surrounding soil under all treatments. Enrichment of ARGs in the plastisphere (resistome) varied across MP types, but not much on MPs size, which also effected the microbial community structure. | [5] |
| Polyvinyl chloride (PVC), polyamide (PA), polystyrene (PS), and polyethylene (PE) MPs | Ammonia-oxidizing, Nitrite-oxidizing bacteria | The sequencing batch reactor operated for a total of 84 days with added MPs at different concentration. The microbial community of aerobic granular sludge from reactor and MPs associated biofilm were explored by Illumina Miseq sequencing. | PVC, PA and PS stimulated the secretion of extracellular polymeric substances and ROS species. Shifts of nitrification genes in aerobic granular sludge and on MPs biofilms | [35] |
| Polyvinyl chloride (PVC) MP pellets (3 mm) | Biofilm | Sterilized MPs, rock, and leaf wrapped into different aggreates, used in bioreactor fed with 5 L river water, incubated for 2 weeks. Biofilms formed on different substrates was investigated using SEM. DNA extracted from biofilms, followed by 16S rRNA gene sequencing and shotgun metagenomics to study the relative abundance of ARGs type. | Biomass of MPs associated biofilm was more than rock biofilm but less than leaf biofilm. MP associated biofilm had distinctive microbial communities structure compared with rock and leaf biofilms. The ARG abundance of biofilm was ~three-fold higher than that of river water, indicating high diversity ARGs enriched by biofilm cultured with water sampled from river. | [27] |
| Targeted Environmental Matrix | Extraction Methods Used | Detection and Quantification Techniques | Type of MPs/NPs Detected | Detection Limit | Reference |
|---|---|---|---|---|---|
| Agricultural soil | Centrifugation | Fluorescence microscopy, Nile red staining, Image processing with Fiji and Python | LDPE, PP, PS, PVC, PET | 6–20 mg/µg | [84] |
| Estuarian muddy sediment | Homogenization, freeze drying | Pyrolysis-GC/MS, FTIR/Raman spectroscopy | PP, PS, PET, PVC | Particles per µg of sample | [85] |
| Drinking water | Filtering/sieving | Stereomicroscopy, FTIR/Raman spectroscopy | PE, PS, PET, PVC | % of particle recovery | [86] |
| Agricultural soil | Ultrasonication, filtration | ATR-FTIR, LDIR | PE, PP, PVC, PA, PTFE | 105 particles per kg soil | [31] |
| Water (pure water and sea water) | Filtration, dialysis | TEM, DLS, SERS | PS, PP, PE | 40 µg/mL (100 nm sized) | [87] |
| Organic clay colloids and humic acid | Asymmetric flow field-flow fractionation (AF4) | AF4-TOC (Total organic carbon), FLD, dRI, UV, Nile red staining | PS | Comparison of different methods | [88] |
| Sediment | Density separation JAMSTEC microplastic-sediment separator (JAMSS) unit | Japan Agency for Marine-Earth Science and Technology (JAMSTEC) | PE, PP, PS, PVC, PET | 94–98% of MPs < 1000 µm | [89] |
| Biosolids | Pressurized liquid extraction (PLE) | Double shot pyrolysis GC/MS | PC, PE, PS, PP, PET, PVC, PMMA | 2.8–6.6 mg/g | [90] |
| Surface water and sediment | Density separation-MPSS | Pyrolysis-GC/MS, Hyperspectral FTIR imaging spectroscopy | PE, PEST, PP, PS, PVC, PC, PMMA, PUR, PA | Mass of MPs in µg m−3 | [91] |
| Marine sponge | Pressurized solvent extraction (PSE) | Pyrolysis-GC/MS, Micro-FTIR | PS, PP, PE, PVC, PC, PL, PA | 6.6–30.2 ng/g | [92] |
| Powdered plastic | Filtration | Pyrolysis GC-ToF, Thermo-gravitimetric analysis, GC-ToF | PP, PS, PVC | <50 µg/L | [93] |
| Aquatic environment | Photo-induced force microscopy (PiFM), Infrared spectroscopy | PET | Count of NP size ~20 nm | [94] | |
| Soil | Density gradient separation | Transmission-type terahertz time-domain spectrometer and NIR hyperspectral imaging system | PS, PVC | 1.12% (tetrahertz) 3.34% (NIR) | [95] |
| Water | self-propelled light-powered MXene-derived γ-Fe2O3/Pt/TiO2 microrobots | SEM, Electrochemical impedance spectroscopy (EIS), XPS, NTA | NPs | 106 NPs per mL | [96] |
| Natural organic matter | Absorptive fractionation | UV-Vis | PSNPs | 7.4 mgC/L | [97] |
| Underwater | On-site detection (NO extraction) | DEP-ACEO-Raman tweezer (DART) (Dieletrophoresis and AC electro-osmosis) | NPs (PS, PMMA) | 1.17 μg·L−1 | [98] |
| Waste-water | Drying | PET depolymerization, LC-MS | MNPs (PET) | Influent-2.0 µg/L, effluent-1.2 µg/L | [99] |
| Water, radish seeds, mussels | Drying (vegetal samples), centrifugation, chemical digestion, filtration (animal samples) | Fluorescent Molecular Rotor (FMR) | PS NPs | 475–563 µg/L in pure water | [100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nath, J.; De, J.; Sur, S.; Banerjee, P. Interaction of Microbes with Microplastics and Nanoplastics in the Agroecosystems—Impact on Antimicrobial Resistance. Pathogens 2023, 12, 888. https://doi.org/10.3390/pathogens12070888
Nath J, De J, Sur S, Banerjee P. Interaction of Microbes with Microplastics and Nanoplastics in the Agroecosystems—Impact on Antimicrobial Resistance. Pathogens. 2023; 12(7):888. https://doi.org/10.3390/pathogens12070888
Chicago/Turabian StyleNath, Jayashree, Jayita De, Shantanu Sur, and Pratik Banerjee. 2023. "Interaction of Microbes with Microplastics and Nanoplastics in the Agroecosystems—Impact on Antimicrobial Resistance" Pathogens 12, no. 7: 888. https://doi.org/10.3390/pathogens12070888
APA StyleNath, J., De, J., Sur, S., & Banerjee, P. (2023). Interaction of Microbes with Microplastics and Nanoplastics in the Agroecosystems—Impact on Antimicrobial Resistance. Pathogens, 12(7), 888. https://doi.org/10.3390/pathogens12070888