
Transcriptome Analysis of Campylobacter jejuni and Campylobacter coli during Cold Stress



Abstract
:1. Introduction
2. Methodology
2.1. Preparation of Retail Meat and Liver Juices
2.2. Bacterial Strains and Growth Conditions
2.3. Preparation of Cells for RNA Isolation
2.4. RNA Extraction
2.5. cDNA Library Preparation, Sequencing and Expression Analysis
2.6. Validation of Differential Gene Expression by qRT-PCR
3. Results
3.1. Overview of RNA-Seq
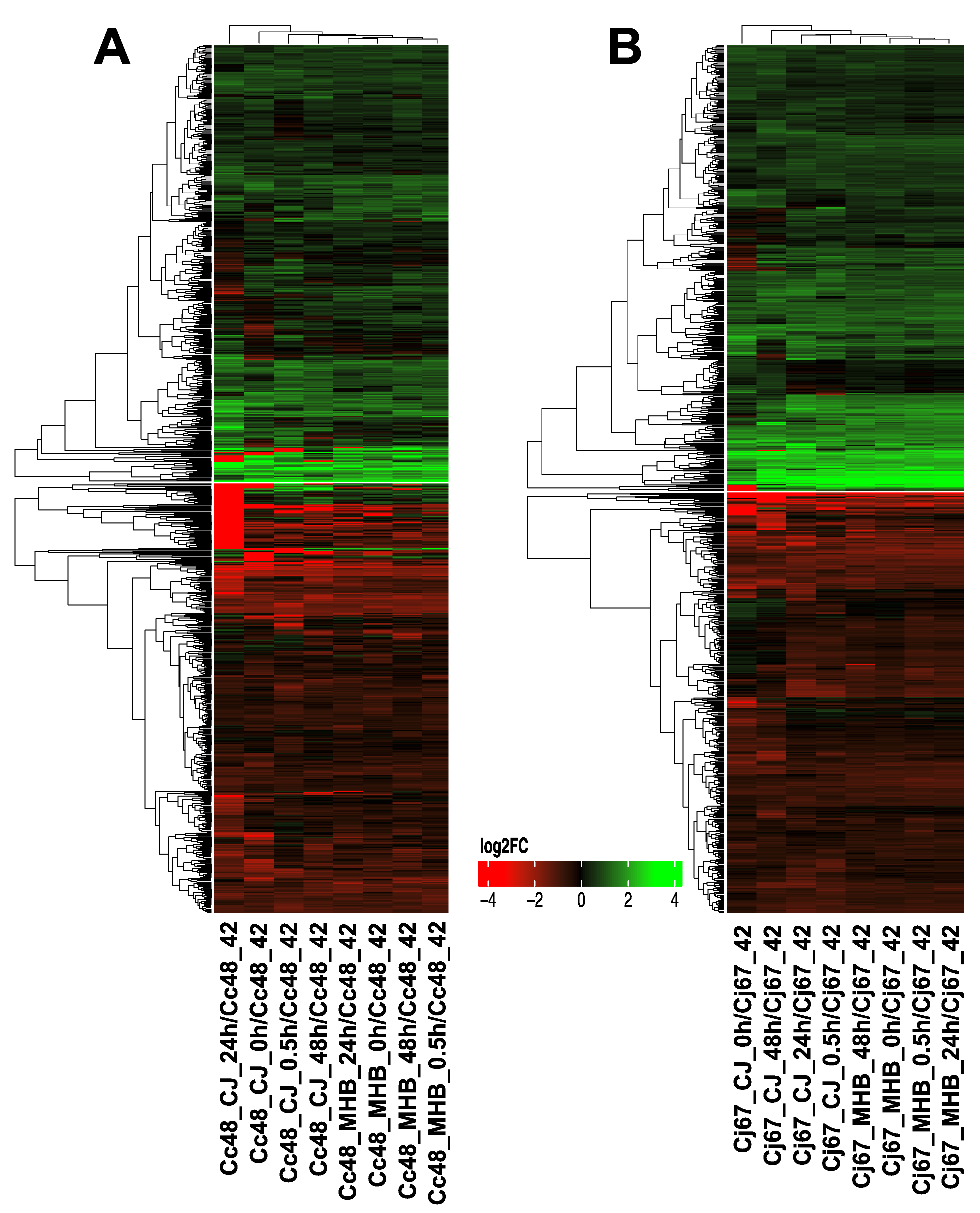
3.2. Influence of Temperature on Gene Expression (4 °C vs. 42 °C)
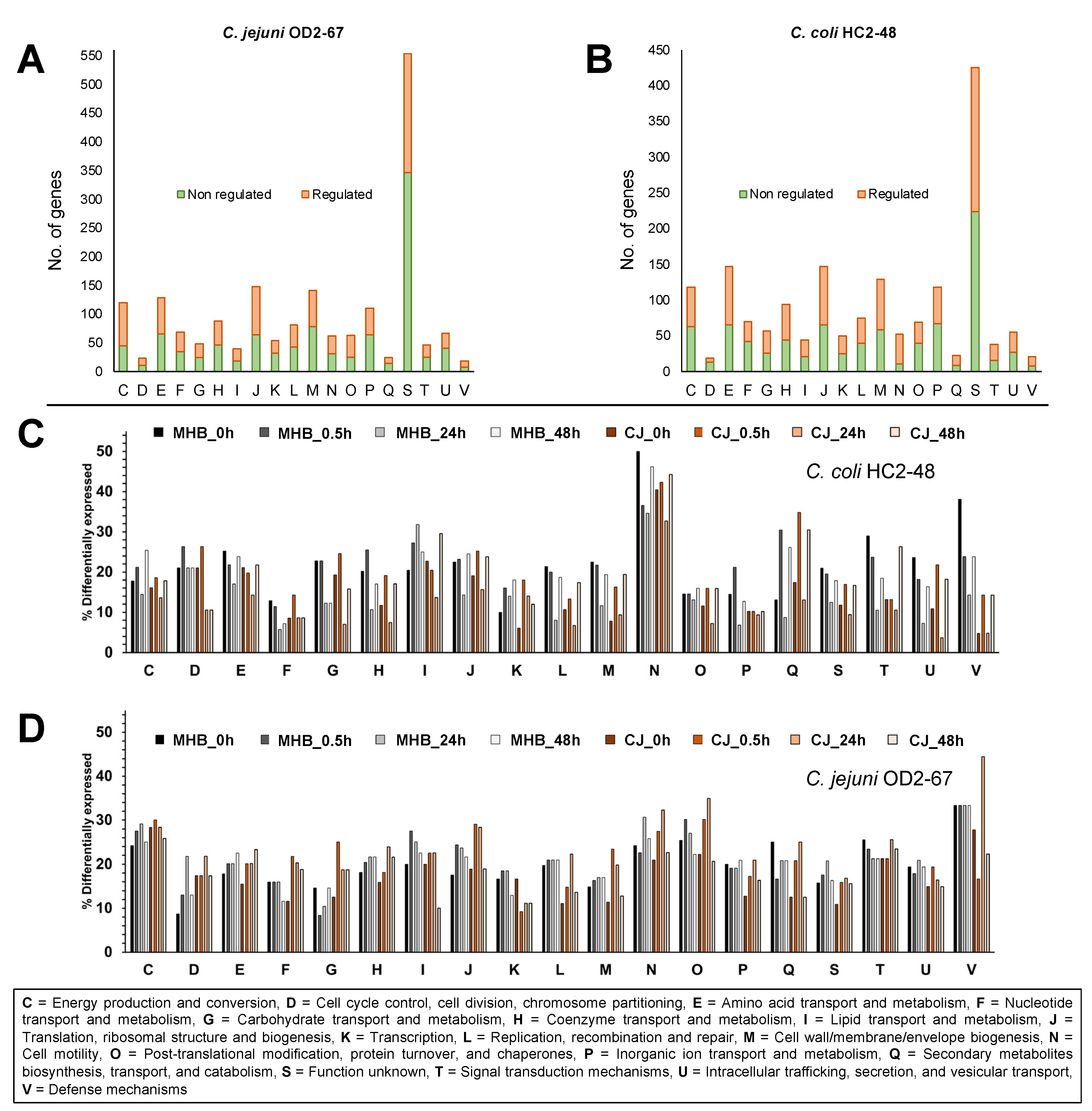
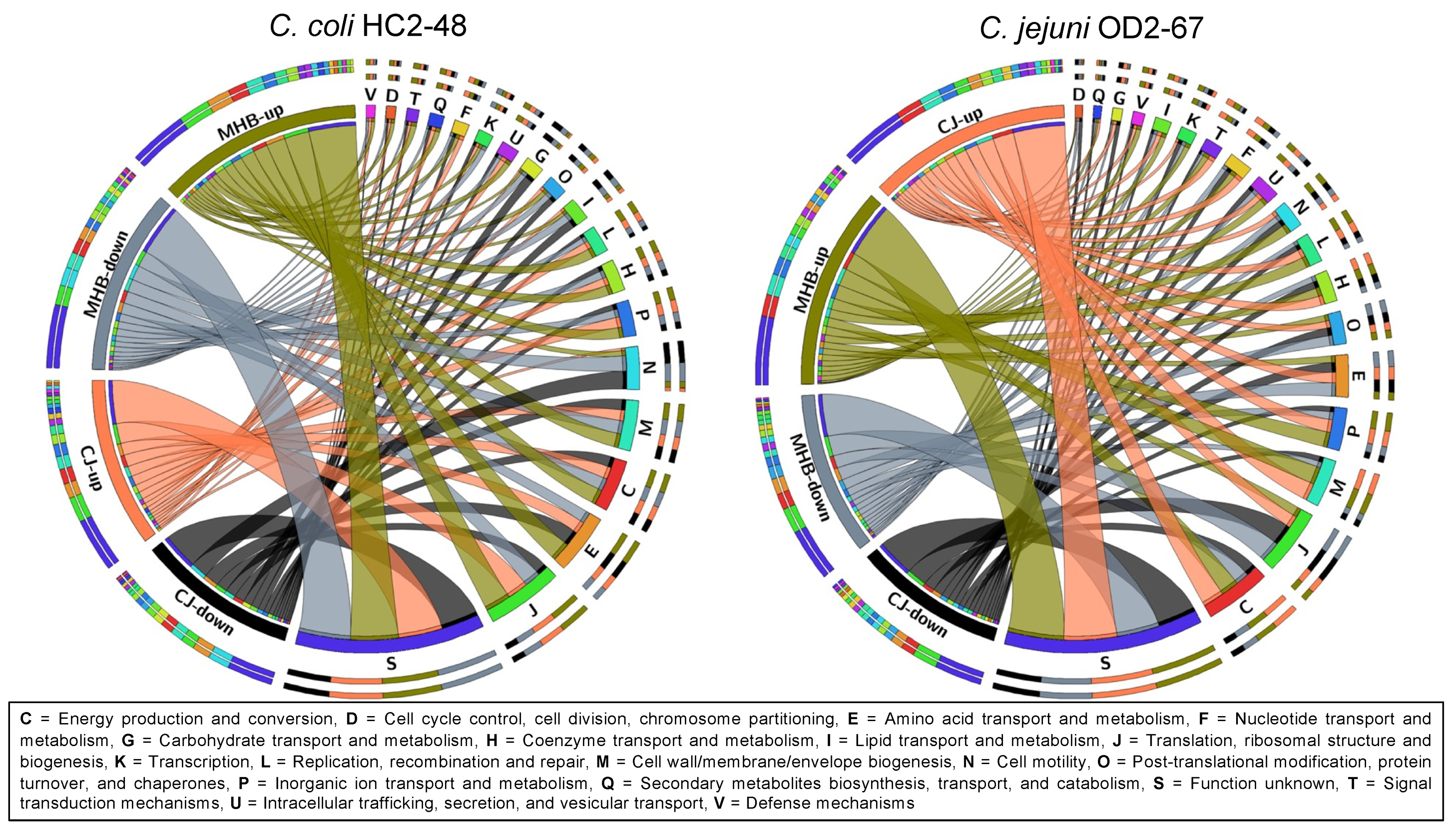
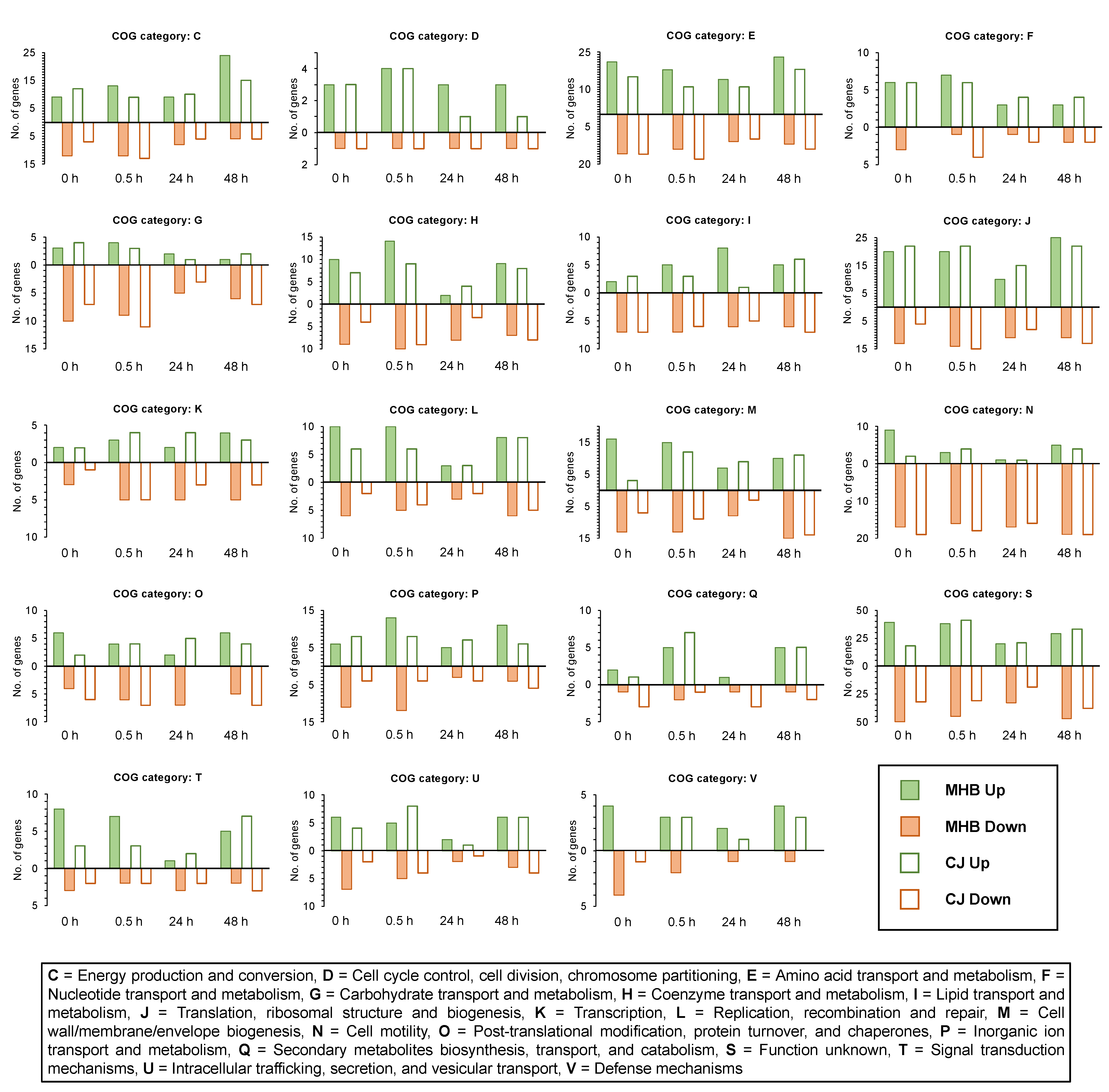
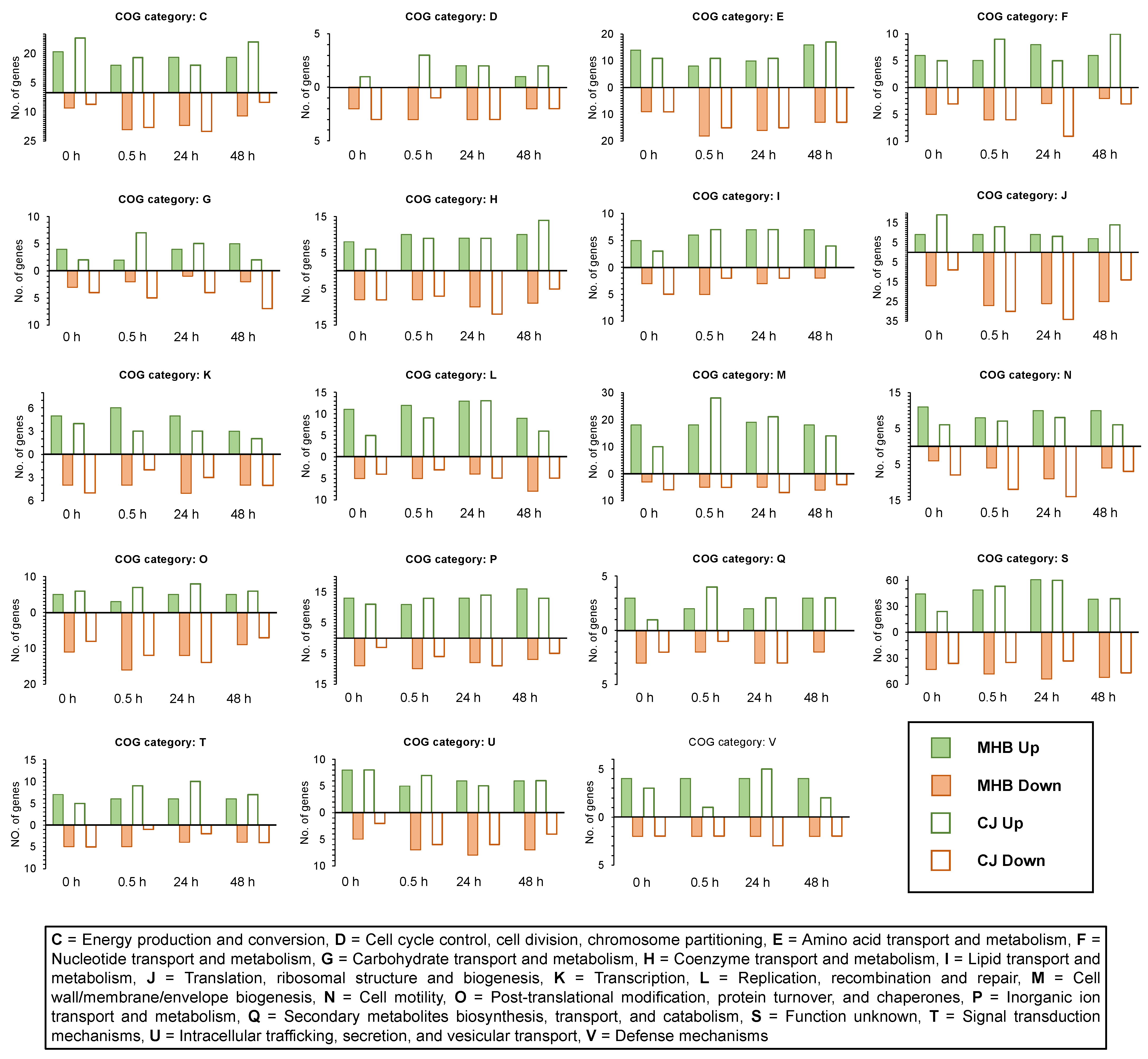
3.3. Functional Group Assignments of DEGs
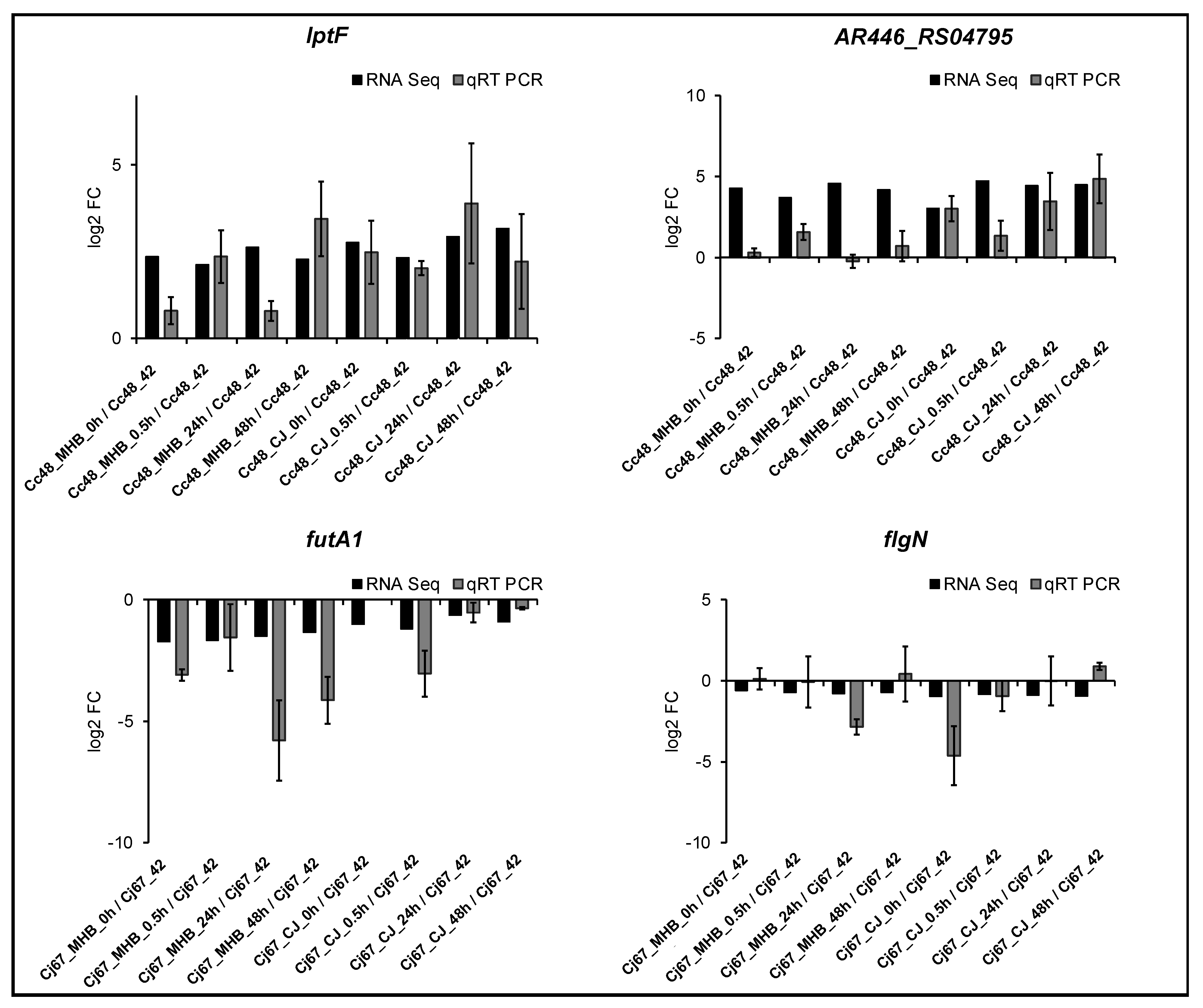
3.4. Validation of RNA-Seq Data by qRT-PCR
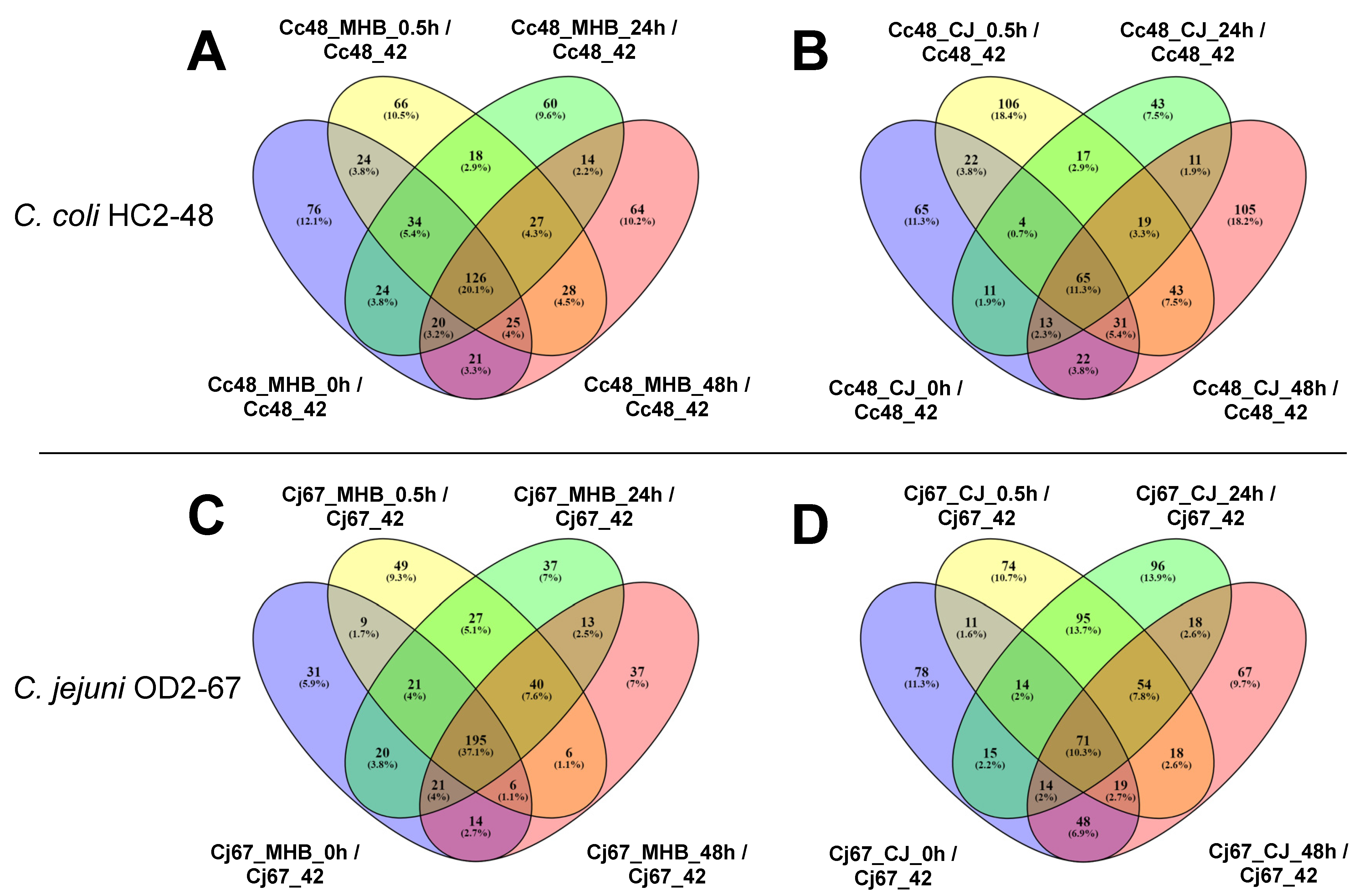
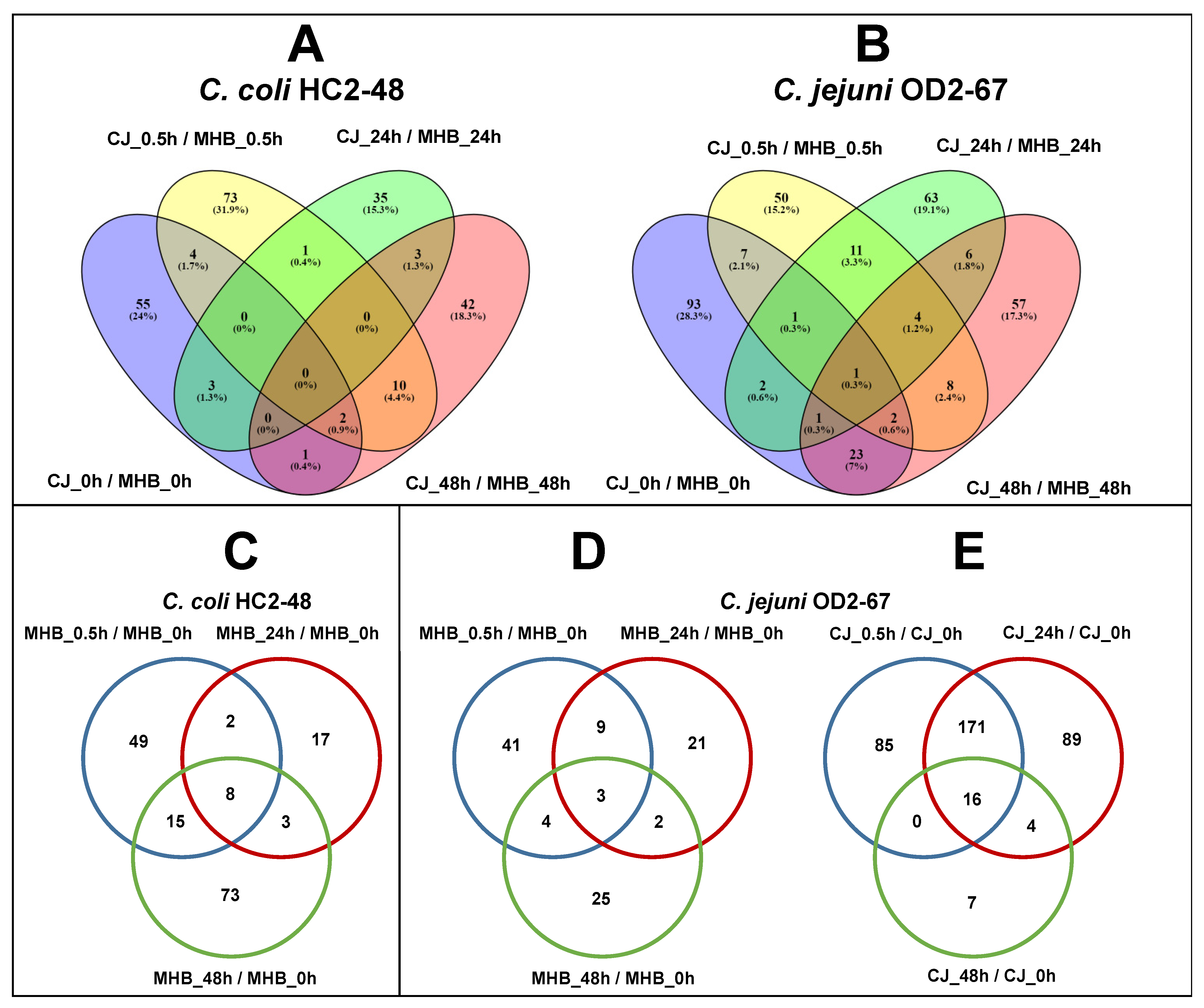
3.5. Common DEGs in C. coli and C. jejuni (4 °C vs. 42 °C)
3.6. Influence of Medium Formulation on Gene Expression (CJ vs. MHB)
3.7. Influence of Incubation Time on Gene Expression (0.5 h vs. 0 h, 24 h vs. 0 h and 48 h vs. 0 h)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karki, A.B.; Wells, H.; Fakhr, M.K. Retail Liver Juices Enhance the Survivability of Campylobacter jejuni and Campylobacter coli at Low Temperatures. Sci. Rep. 2019, 9, 2733. [Google Scholar] [CrossRef] [Green Version]
- Bronowski, C.; James, C.E.; Winstanley, C. Role of Environmental Survival in Transmission of Campylobacter jejuni. FEMS Microbiol. Lett. 2014, 356, 8–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birk, T.; Ingmer, H.; Andersen, M.T.; Jørgensen, K.; Brøndsted, L. Chicken Juice, a Food-Based Model System Suitable to Study Survival of Campylobacter jejuni. Lett. Appl. Microbiol. 2004, 38, 66–71. [Google Scholar] [CrossRef]
- Noormohamed, A.; Fakhr, M.K. A Higher Prevalence Rate of Campylobacter in Retail Beef Livers Compared to Other Beef and Pork Meat Cuts. Int. J. Environ. Res. Public Health 2013, 10, 2058–2068. [Google Scholar] [CrossRef] [Green Version]
- Noormohamed, A.; Fakhr, M.K. Prevalence and Antimicrobial Susceptibility of Campylobacter spp. in Oklahoma Conventional and Organic Retail Poultry. Open Microbiol. J. 2014, 8, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Noormohamed, A.; Fakhr, M.K. Incidence and Antimicrobial Resistance Profiling of Campylobacter in Retail Chicken Livers and Gizzards. Foodborne Pathog. Dis. 2012, 9, 617–624. [Google Scholar] [CrossRef]
- Karki, A.B.; Marasini, D.; Oakey, C.K.; Mar, K.; Fakhr, M.K. Campylobacter coli from Retail Liver and Meat Products Is More Aerotolerant than Campylobacter jejuni. Front. Microbiol. 2018, 9, 2951. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Andrews, K.J.; McMullen, L.M.; Jeon, B. Tolerance to Stress Conditions Associated with Food Safety in Campylobacter jejuni Strains Isolated from Retail Raw Chicken. Sci. Rep. 2019, 9, 11915. [Google Scholar] [CrossRef] [Green Version]
- Moen, B.; Oust, A.; Langsrud, Ø.; Dorrell, N.; Marsden, G.L.; Hinds, J.; Kohler, A.; Wren, B.W.; Rudi, K. Explorative Multifactor Approach for Investigating Global Survival Mechanisms of Campylobacter jejuni under Environmental Conditions. Appl. Environ. Microbiol. 2005, 71, 2086–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, S.P.W.; Gupta, S.; Baig, A.; Wright, E.; Wedley, A.; Jensen, A.N.; Lora, L.L.; Humphrey, S.; Skovgard, H.; MacLeod, K.; et al. Genome-Wide Fitness Analyses of the Foodborne Pathogen Campylobacter jejuni in In Vitro and In Vivo Models. Sci. Rep. 2017, 7, 1251. [Google Scholar] [CrossRef] [Green Version]
- Bronowski, C.; Mustafa, K.; Goodhead, I.; James, C.E.; Nelson, C.; Lucaci, A.; Wigley, P.; Humphrey, T.J.; Williams, N.J.; Winstanley, C. Campylobacter jejuni Transcriptome Changes during Loss of Culturability in Water. PLoS ONE 2017, 12, e0188936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligowska, M.; Cohn, M.T.; Stabler, R.A.; Wren, B.W.; Brøndsted, L. Effect of Chicken Meat Environment on Gene Expression of Campylobacter jejuni and Its Relevance to Survival in Food. Int. J. Food Microbiol. 2011, 145, S111–S115. [Google Scholar] [CrossRef] [PubMed]
- Stintzi, A.; Whitworth, L. Investigation of the Campylobacter jejuni Cold-Shock Response by Global Transcript Profiling. Genome Lett. 2003, 2, 18–27. [Google Scholar] [CrossRef]
- Butcher, J.; Stintzi, A. The Transcriptional Landscape of Campylobacter jejuni under Iron Replete and Iron Limited Growth Conditions. PLoS ONE 2013, 8, e79475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedel, C.; Förstner, K.U.; Püning, C.; Alter, T.; Sharma, C.M.; Gölz, G. Differences in the Transcriptomic Response of Campylobacter coli and Campylobacter Lari to Heat Stress. Front. Microbiol. 2020, 11, 523. [Google Scholar] [CrossRef]
- Marasini, D.; Fakhr, M.K. Complete Genome Sequences of Campylobacter jejuni Strains OD267 and WP2202 Isolated from Retail Chicken Livers and Gizzards Reveal the Presence of Novel 116-Kilobase and 119-Kilobase Megaplasmids with Type VI Secretion Systems. Genome Announc. 2016, 4, e01060-16. [Google Scholar] [CrossRef] [Green Version]
- Marasini, D.; Fakhr, M.K. Complete Genome Sequences of the Plasmid-Bearing Campylobacter coli Strains HC2-48, CF2-75, and CO2-160 Isolated from Retail Beef Liver. Genome Announc. 2016, 4, e01004-16. [Google Scholar] [CrossRef] [Green Version]
- Masek, T.; Vopalensky, V.; Suchomelova, P.; Pospisek, M. Denaturing RNA Electrophoresis in TAE Agarose Gels. Anal. Biochem. 2005, 336, 46–50. [Google Scholar] [CrossRef]
- Carroll, R.K.; Weiss, A.; Shaw, L.N. Rna-Sequencing of Staphylococcus Aureus Messenger RNA. In Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2016; Volume 1373, pp. 131–141. [Google Scholar]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A Hierarchical, Functionally and Phylogenetically Annotated Orthology Resource Based on 5090 Organisms and 2502 Viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. Quantification Strategies in Real-Time PCR. In A-Z of Quantitative PCR; International University Line.: La Jolla, CA, USA, 2004; Volume 29, pp. 87–112. [Google Scholar]
- Boysen, L.; Knøchel, S.; Rosenquist, H. Survival of Campylobacter jejuni in Different Gas Mixtures. FEMS Microbiol. Lett. 2007, 266, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; McMullen, L.M.; Chui, L.; Jeon, B. Differential Survival of Hyper-Aerotolerant Campylobacter jejuni under Different Gas Conditions. Front. Microbiol. 2017, 8, 954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Chin, M.; Nusbaum, C.; Birren, B.W.; Livny, J. How Deep Is Deep Enough for RNA-Seq Profiling of Bacterial Transcriptomes? BMC Genom. 2012, 13, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, D.D.; Madsen, M. Cytolethal Distending Toxins of Campylobacter jejuni: Genetics, Structure, Mode of Action, Epidemiology, and the Role of CDT in Campylobacter Pathogenesis. Genome Lett. 2003, 2, 73–82. [Google Scholar] [CrossRef]
- Palyada, K.; Sun, Y.Q.; Flint, A.; Butcher, J.; Naikare, H.; Stintzi, A. Characterization of the Oxidative Stress Stimulon and PerR Regulon of Campylobacter jejuni. BMC Genom. 2009, 10, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gennaris, A.; Ezraty, B.; Henry, C.; Agrebi, R.; Vergnes, A.; Oheix, E.; Bos, J.; Leverrier, P.; Espinosa, L.; Szewczyk, J.; et al. Repairing Oxidized Proteins in the Bacterial Envelope Using Respiratory Chain Electrons. Nature 2015, 528, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Butcher, J.; Handley, R.A.; van Vliet, A.H.M.; Stintzi, A. Refined Analysis of the Campylobacter jejuni Iron-Dependent/Independent Fur- and PerR-Transcriptomes. BMC Genom. 2015, 16, 498. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A. Pathways of Oxidative Damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Anjem, A.; Imlay, J.A. Mononuclear Iron Enzymes Are Primary Targets of Hydrogen Peroxide Stress. J. Biol. Chem. 2012, 287, 15544–15556. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.; Imlay, J.A. Superoxide Poisons Mononuclear Iron Enzymes by Causing Mismetallation. Mol. Microbiol. 2013, 89, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [Green Version]
- Flint, A.; Stintzi, A.; Saraiva, L.M. Oxidative and Nitrosative Stress Defences of Helicobacter and Campylobacter Species That Counteract Mammalian Immunity. FEMS Microbiol. Rev. 2016, 40, 938–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, K.; Mulholland, F.; Pearson, B.M.; Pin, C.; McNicholl-Kennedy, J.; Ketley, J.M.; Wells, J.M. Campylobacter jejuni Gene Expression in Response to Iron Limitation and the Role of Fur. Microbiology 2005, 151, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Bröms, J.E.; Kumar, R.; Sjöstedt, A. The Role of ClpB in Bacterial Stress Responses and Virulence. Front. Mol. Biosci. 2021, 8, 668910. [Google Scholar] [CrossRef]
- Liaw, J.; Hong, G.; Davies, C.; Elmi, A.; Sima, F.; Stratakos, A.; Stef, L.; Pet, I.; Hachani, A.; Corcionivoschi, N.; et al. The Campylobacter jejuni Type VI Secretion System Enhances the Oxidative Stress Response and Host Colonization. Front. Microbiol. 2019, 10, 2864. [Google Scholar] [CrossRef]
- Marasini, D.; Karki, A.B.; Bryant, J.M.; Sheaff, R.J.; Fakhr, M.K. Molecular Characterization of Megaplasmids Encoding the Type VI Secretion System in Campylobacter jejuni Isolated from Chicken Livers and Gizzards. Sci. Rep. 2020, 10, 12514. [Google Scholar] [CrossRef]
- Ilbert, M.; Méjean, V.; Giudici-Orticoni, M.T.; Samama, J.P.; Iobbi-Nivol, C. Involvement of a Mate Chaperone (TorD) in the Maturation Pathway of Molybdoenzyme TorA. J. Biol. Chem. 2003, 278, 28787–28792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellars, M.J.; Hall, S.J.; Kelly, D.J. Growth of Campylobacter jejuni Supported by Respiration of Fumarate, Nitrate, Nitrite, Trimethylamine-N-Oxide, or Dimethyl Sulfoxide Requires Oxygen. J. Bacteriol. 2002, 184, 4187–4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, K.L.N.; Weiss, D.S. The Escherichia Coli Cell Division Protein FtsW Is Required to Recruit Its Cognate Transpeptidase, FtsI (PBP3), to the Division Site. J. Bacteriol. 2002, 184, 904–912. [Google Scholar] [CrossRef] [Green Version]
- Hamoen, L.W. Cell Division Blockage: But This Time by a Surprisingly Conserved Protein. Mol. Microbiol. 2011, 81, 1–3. [Google Scholar] [CrossRef]
- Richards, J.; Liu, Q.; Pellegrini, O.; Celesnik, H.; Yao, S.; Bechhofer, D.H.; Condon, C.; Belasco, J.G. An RNA Pyrophosphohydrolase Triggers 5′-Exonucleolytic Degradation of MRNA in Bacillus Subtilis. Mol. Cell 2011, 43, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Blanco, D.A.; Shell, S.S. Regulation of MRNA Stability During Bacterial Stress Responses. Front. Microbiol. 2020, 11, 2111. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | MHB | CJ | ||||||
|---|---|---|---|---|---|---|---|---|
| Time of Incubation at 4 °C | 0 h | 0.5 h | 24 h | 48 h | 0 h | 0.5 h | 24 h | 48 h |
| C. coli HC2-48, Total genes analyzed = 1651 | ||||||||
| Number of regulated genes | 350 | 348 | 323 | 325 | 233 | 307 | 183 | 309 |
| Upregulated (%) | 49.14% | 52.01% | 45.82% | 54.46% | 49.79% | 51.47% | 53.55% | 50.81% |
| Downregulated (%) | 50.85% | 47.99% | 54.18% | 45.54% | 50.21% | 48.53% | 46.45% | 49.19% |
| C. jejuni OD2-67, Total genes analyzed = 1774 | ||||||||
| Number of regulated genes | 317 | 353 | 374 | 332 | 270 | 356 | 377 | 309 |
| Upregulated (%) | 56.15% | 47.03% | 51.87% | 50.90% | 56.67% | 57.02% | 50.93% | 57.93% |
| Downregulated (%) | 43.85% | 52.97% | 48.13% | 49.10% | 43.33% | 42.98% | 49.07% | 42.07% |
| Functional Group | Upregulated Genes | Downregulated Genes |
|---|---|---|
| Energy production/conversion | ppa/inorganic diphosphatase petC (AR446_RS02380)/cytochrome c1 | prpC (AR446_RS06780)/citrate synthase/methylcitrate synthase |
| Cell cycle control, cell division, chromosome partitioning | mreB/rod shape-determining protein | pseA/pseudaminic acid biosynthesis protein PseA |
| Amino acid transport/metabolism | hisH/imidazole glycerol phosphate synthase subunit HisH dapF (AR446_RS00670)/diaminopimelate epimerase cysK cysteine synthase A pseC/UDP-4-amino-4, 6-dideoxy-N-acetyl-beta-L-altrosamine transaminase ycaD (AR446_RS04050)/MFS transporter glnH (AR446_RS04215)/transporter-substrate-binding-domain-containing protein | |
| Nucleotide transport/metabolism | purl/phosphoribosylformylglycinamidine synthase subunit PurL | dut (AR446_RS01050)/dUTP diphosphatase |
| Carbohydrate transport/metabolism | pseB/UDP-N-acetylglucosamine 4,6-dehydratase (inverting) # | |
| Coenzyme transport/metabolism | hemE (AR446_RS02015)/uroporphyrinogen decarboxylase | |
| Lipid transport/metabolism | cdsA (AR446_RS01495)/phosphatidate cytidylyltransferase prpE (AR446_RS06790)/propionyl-CoA synthetase dxr/1-deoxy-D-xylulose-5-phosphate reductoisomerase acs acetate—CoA ligase | |
| Translation, ribosomal structure/ biogenesis | rplM/50S ribosomal protein L13 rplQ/50S ribosomal protein L17 rpsI/30S ribosomal protein S9 hisS (AR446_RS04490)/histidine--tRNA ligase mnmE tRNA uridine-5 carboxymethylaminomethyl(34) synthesis GTPase MnmE | tet(O) tetracycline resistance ribosomal protection protein Tet(O) |
| Transcription | rpoA (AR446_RS00265)/DNA-directed RNA polymerase subunit alpha | |
| Cell wall/membrane/envelope biogenesis | rlpA/septal ring lytic transglycosylase RlpA family protein | |
| Cell motility | flhB/flagellar biosynthesis protein FlhB fliD/flagellar filament capping protein FliD flgL/flagellar hook-associated protein FlgL flgG/flagellar basal-body rod protein FlgG flaG (AR446_RS05375)/flagellar protein FlaG fliS/flagellar export chaperone FliS flgH/flagellar basal body L-ring protein FlgH flgB/flagellar basal body rod protein FlgB flgF (AR446_RS04770)/flagellar hook-basal body protein flgK/flagellar hook-associated protein FlgK flgM (AR446_RS00985) */flagellar biosynthesis anti-sigma factor FlgM flgN (AR446_RS00980) */flagellar protein FlgN | |
| Secondary metabolites biosynthesis/transport/catabolism | paaI (AR446_RS03420)/PaaI family thioesterase | |
| Function unknown | AR446_RS07500/hypothetical protein lptF (AR446_RS04170)/LptF/LptG family permease | cj1450 (AR446_RS01055)/hypothetical protein actP (AR446_RS08050)/cation acetate symporter AR446_RS00045/hypothetical protein AR446_RS07515/hypothetical protein AR446_RS08285/hypothetical protein AR446_RS01490/hypothetical protein rny/ribonuclease Y |
| Functional Group | Upregulated Genes | Downregulated Genes |
|---|---|---|
| Energy production/ conversion | nuoA/NAD(P)H-quinone oxidoreductase subunit 3 lldP/L-lactate permease nuoM/NADH-quinone oxidoreductase subunit M atpB/F0F1 ATP synthase subunit A ccpA/cytochrome-c peroxidase hdrC/FAD-binding oxidoreductase | aldA/aldehyde dehydrogenase msrP/protein-methionine-sulfoxide reductase catalytic subunit MsrP |
| Amino acid transport/ metabolism | aroQ/type II 3-dehydroquinate dehydratase | map/type I methionyl aminopeptidase dapA/4-hydroxy-tetrahydrodipicolinate synthase leuA/2-isopropylmalate synthase trpE * (A0W68_RS01665)/anthranilate synthase component I family protein |
| Coenzyme transport/metabolism | Pcm/protein-L-isoaspartate (D-aspartate)O-methyltransferase | |
| Translation, ribosomal structure/biogenesis | ridA (A0W68_RS03730)/RidA family protein | rim/16S rRNA processing protein RimM rpsM/30S ribosomal protein S13 rpsK/30S ribosomal protein S11 |
| Transcription | A0W68_RS06530/response regulator transcription factor | |
| Replication, recombination, repair | dnaN (A0W68_RS00010)/DNA polymerase III subunit beta hup (A0W68_RS04770)/HU family DNA-binding protein | |
| Cell wall/membrane/envelope biogenesis | pseF/pseudaminic acid cytidylyltransferase A0W68_RS0595/hypothetical protein lspA (A0W68_RS01740)/lipoprotein signal peptidase | cfa (A0W68_RS06140)/class I SAM-dependent methyltransferase |
| Cell motility | flip/flagellar type III secretion system pore protein FliP methyl accepting chemotaxis proteins (A0W68_RS08070, A0W68_RS00090) | fliS/flagellar export chaperone FliS flgN */flagellar protein FlgN |
| Post-translational modification, protein turnover and chaperones | clpP/ATP-dependent Clp endopeptidase proteolytic subunit ClpP | grpE/nucleotide exchange factor GrpE cbpA (A0W68_RS06375)/DnaJ family protein |
| Inorganic ion transport and metabolism | Sodium dependent transporter (A0W68_RS04880, A0W68_RS02830) ctf/non-heme ferritin | futA1 (A0W68_RS00880)/Fe(3+) ABC transporter substrate-binding protein |
| Secondary metabolites biosynthesis/transport/catabolism | fahA (A0W68_RS00100)/fumarylacetoacetate hydrolase family protein | |
| Function unknown | A0W68_RS04155/hypothetical protein arsP /organoarsenical efflux permease ArsP pseE1 (A0W68_RS07185)/motility associated factor glycosyltransferase family protein yccA (A0W68_RS01145)/Bax inhibitor-1/YccA family protein A0W68_RS01955/SPOR-domain-containing protein A0W68_RS02215/DUF374-domain-containing protein A0W68_RS07500/hypothetical protein | A0W68_RS01890/hypothetical protein A0W68_RS02435/YkgJ family cysteine cluster protein ciaC/invasion antigen CiaC uup (A0W68_RS04650)/ABC-F family ATP-binding-cassette-domain-containing protein A0W68_RS04080/cupin-domain-containing protein A0W68_RS07830/LysR family transcriptional regulator dba/disulfide bond formation protein Dba A0W68_RS00330/membrane protein), A0W68_RS06720/hypothetical protein |
| Intracellular trafficking, secretion and vesicular transport | dctA (A0W68_RS06185)/cation:dicarboxylase symporter family transporter | yajC/preprotein translocase subunit YajC |
| Defense mechanism | macB (A0W68_RS02860)/ABC transporter permease yokD (A0W68_RS06985)/aminoglycoside N(3)-acetyltransferase |
| Functional Group | Upregulated Genes | Downregulated Genes |
|---|---|---|
| Energy production/conversion | trxB/thioredoxin-disulfide reductase | |
| Cell cycle control, cell division, chromosome partitioning | mreB #/rod shape-determining protein | |
| Coenzyme transport/ metabolism | hemE/uroporphyrinogen decarboxylase | |
| Translation, ribosomal structure/biogenesis | rplM/50S ribosomal protein L13 | |
| Cell motility | flaG/flagellar protein FlaG fliS/flagellar export chaperone FliS flgG/flagellar basal-body rod protein FlgG flgP */flagellar assembly lipoprotein FlgP flgN */flagellar protein FlgN | |
| Post translational modification, chaperones | clpB/AAA family ATPase | |
| Function unknown | AR446_RS06165/A0W68_RS01890/hypothetical protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karki, A.B.; Khatri, B.; Fakhr, M.K. Transcriptome Analysis of Campylobacter jejuni and Campylobacter coli during Cold Stress. Pathogens 2023, 12, 960. https://doi.org/10.3390/pathogens12070960
Karki AB, Khatri B, Fakhr MK. Transcriptome Analysis of Campylobacter jejuni and Campylobacter coli during Cold Stress. Pathogens. 2023; 12(7):960. https://doi.org/10.3390/pathogens12070960
Chicago/Turabian StyleKarki, Anand B., Bhuwan Khatri, and Mohamed K. Fakhr. 2023. "Transcriptome Analysis of Campylobacter jejuni and Campylobacter coli during Cold Stress" Pathogens 12, no. 7: 960. https://doi.org/10.3390/pathogens12070960
APA StyleKarki, A. B., Khatri, B., & Fakhr, M. K. (2023). Transcriptome Analysis of Campylobacter jejuni and Campylobacter coli during Cold Stress. Pathogens, 12(7), 960. https://doi.org/10.3390/pathogens12070960