
Triplex Crystal Digital PCR for the Detection and Differentiation of the Wild-Type Strain and the MGF505-2R and I177L Gene-Deleted Strain of African Swine Fever Virus

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viral Strains
2.2. Clinical Samples
2.3. Design of Primers and Probes
2.4. Extraction of Nucleic Acids
2.5. Construction of the Standard Plasmids
2.6. Optimization of the Reaction Conditions
2.7. Analytical Specificity Analysis
2.8. Analytical Sensitivity Analysis
2.9. Repeatability Analysis
2.10. Detection of the Clinical Samples
3. Results
3.1. Construction of the Standard Plasmids
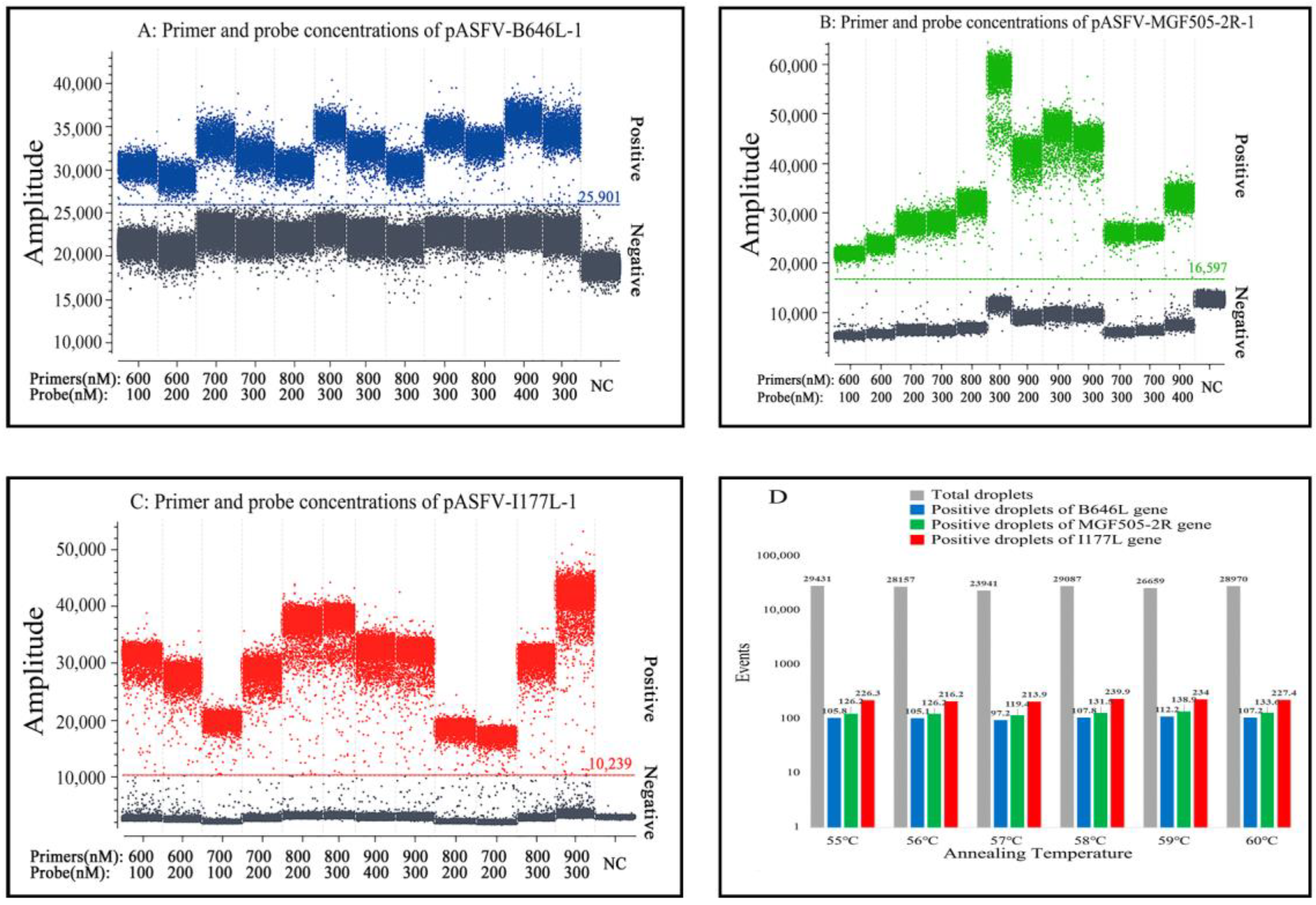
3.2. Determination of the Optimal Parameters
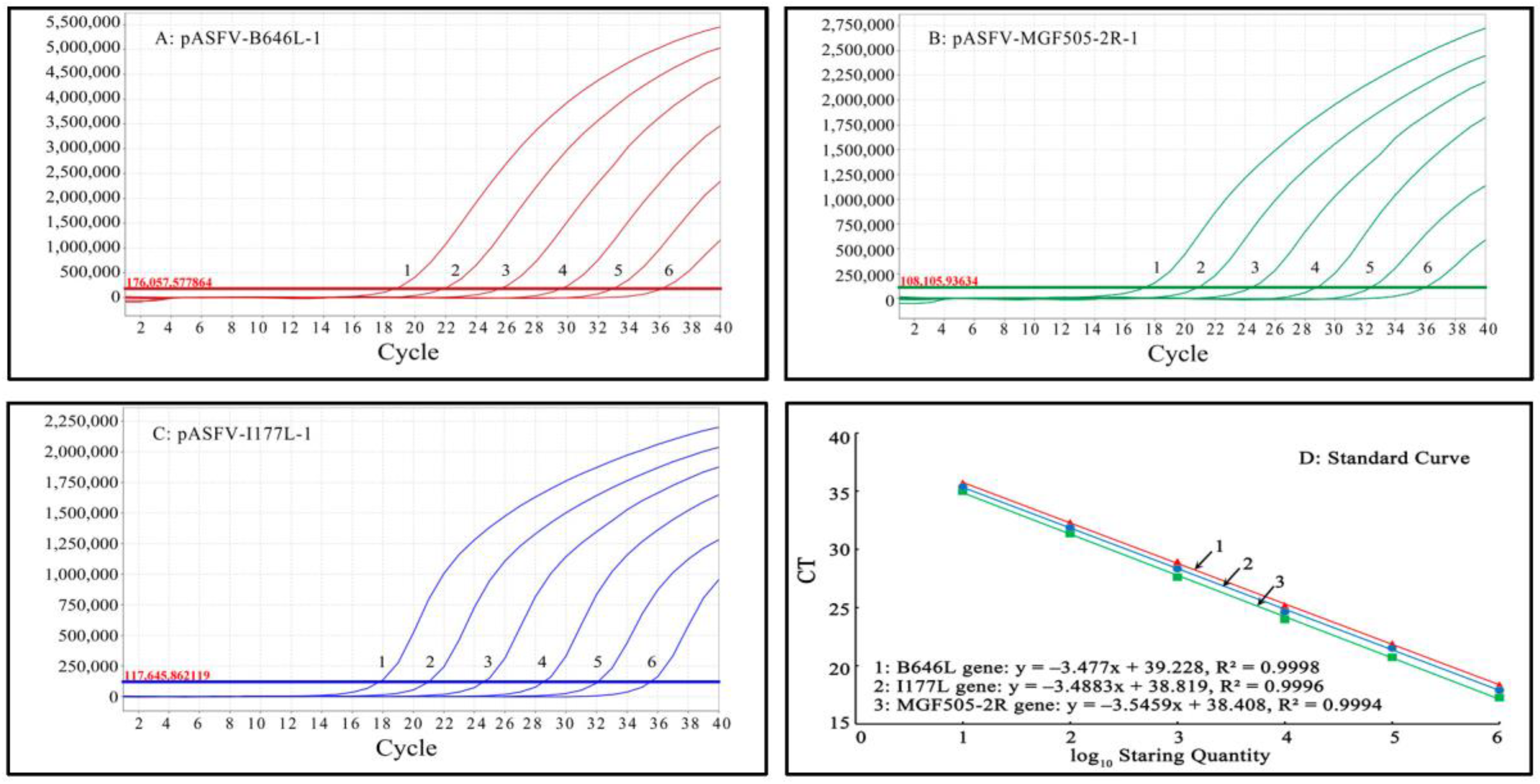
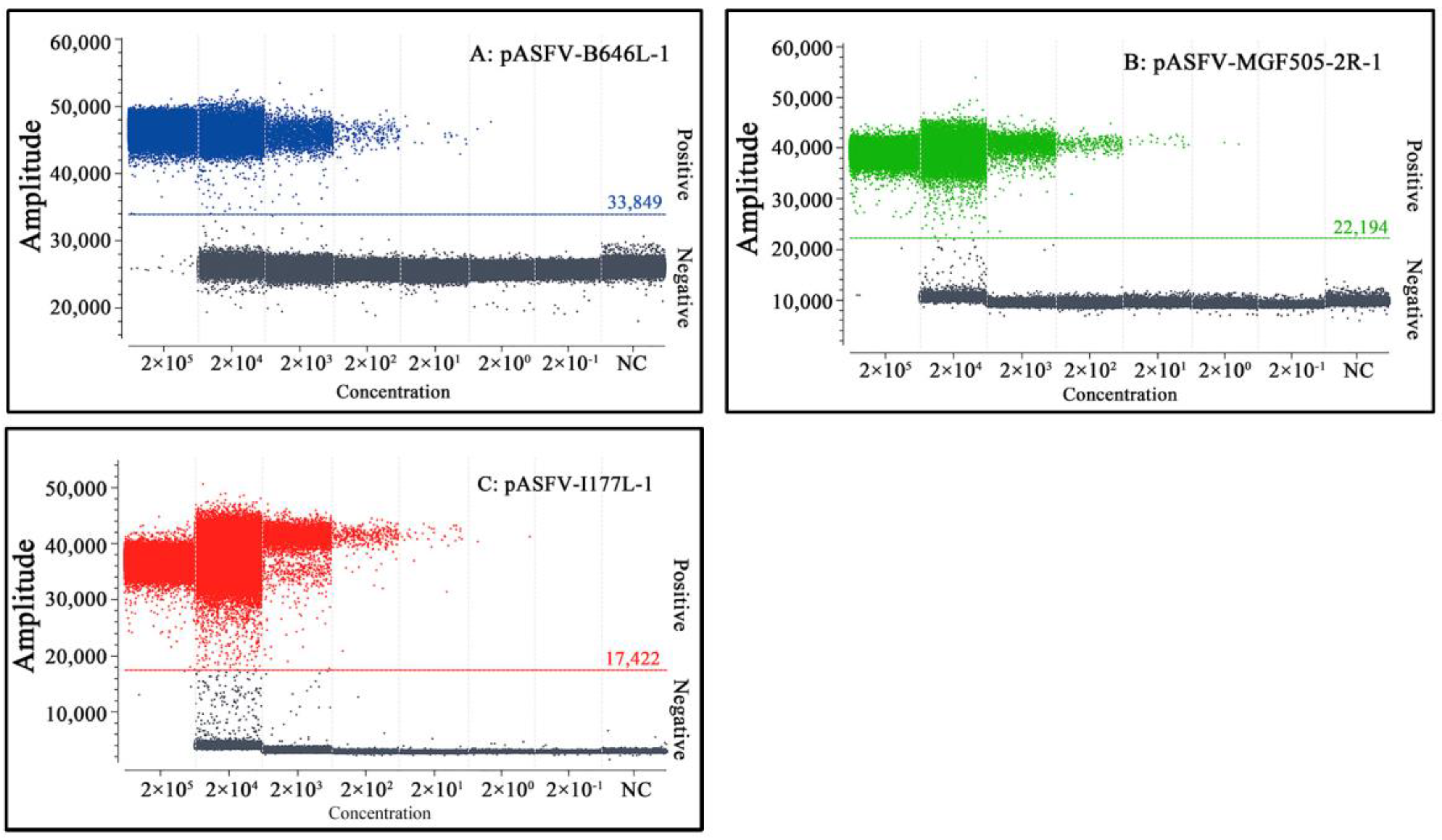
3.3. Generation of the Standard Curves
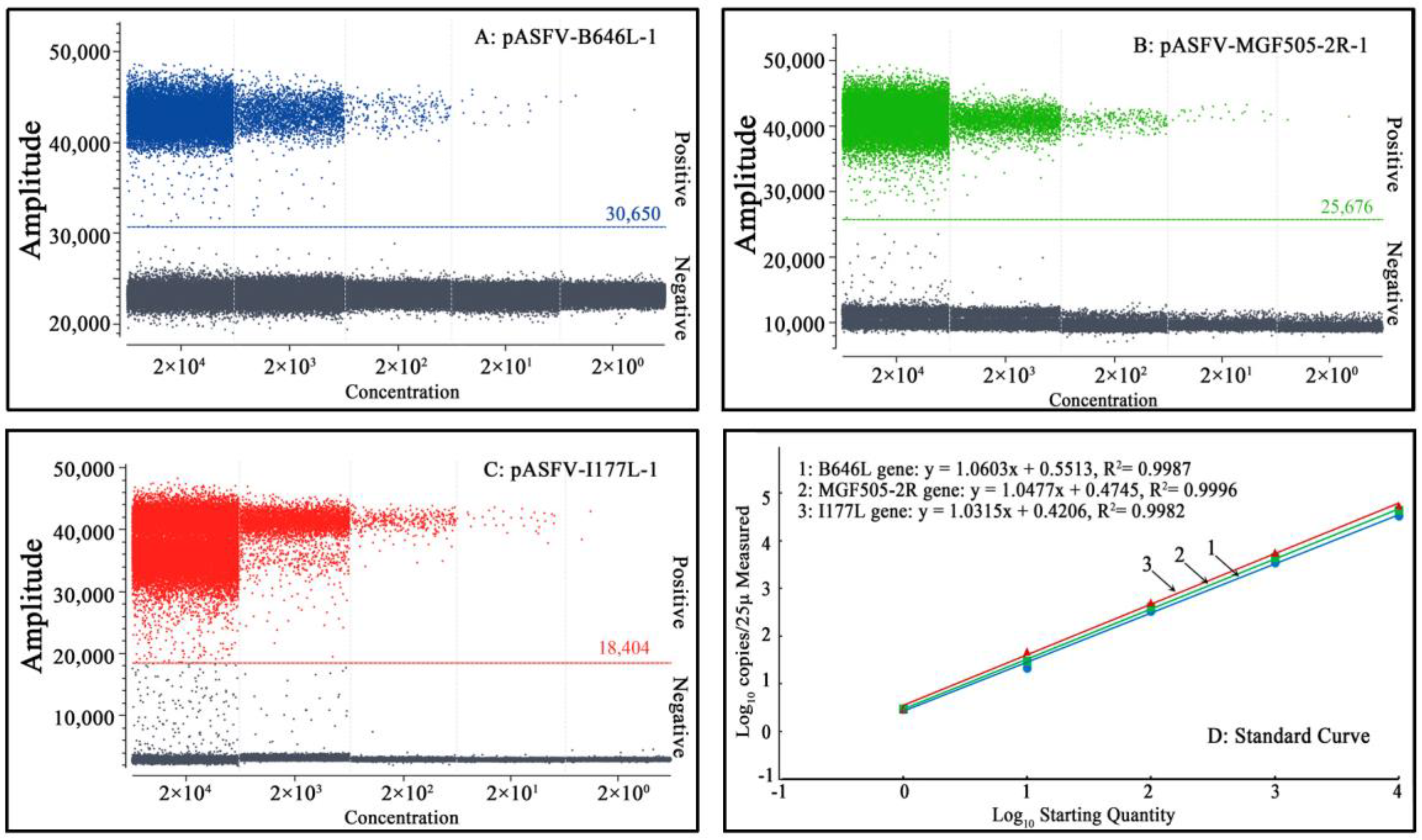
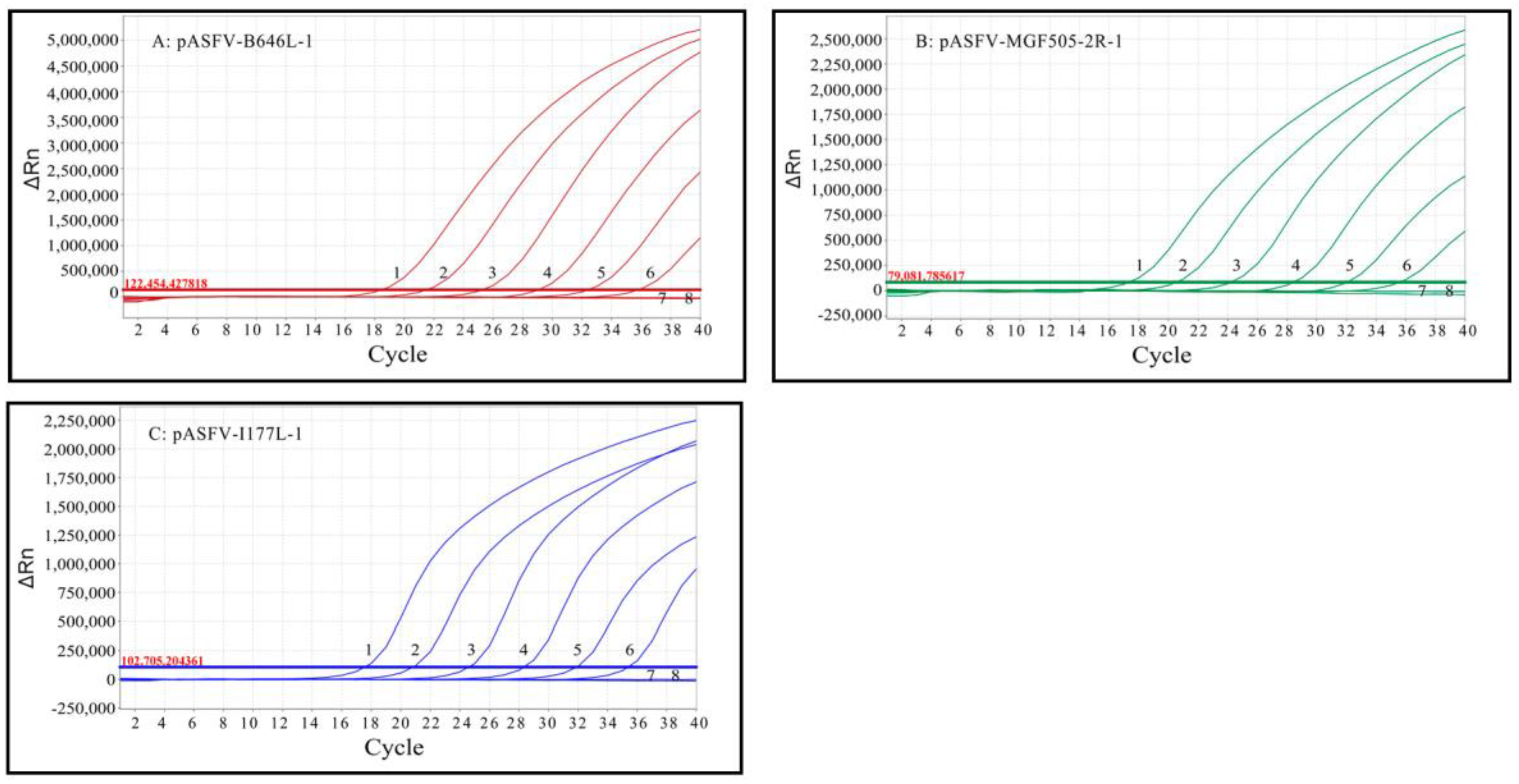
3.4. Sensitivity Analysis
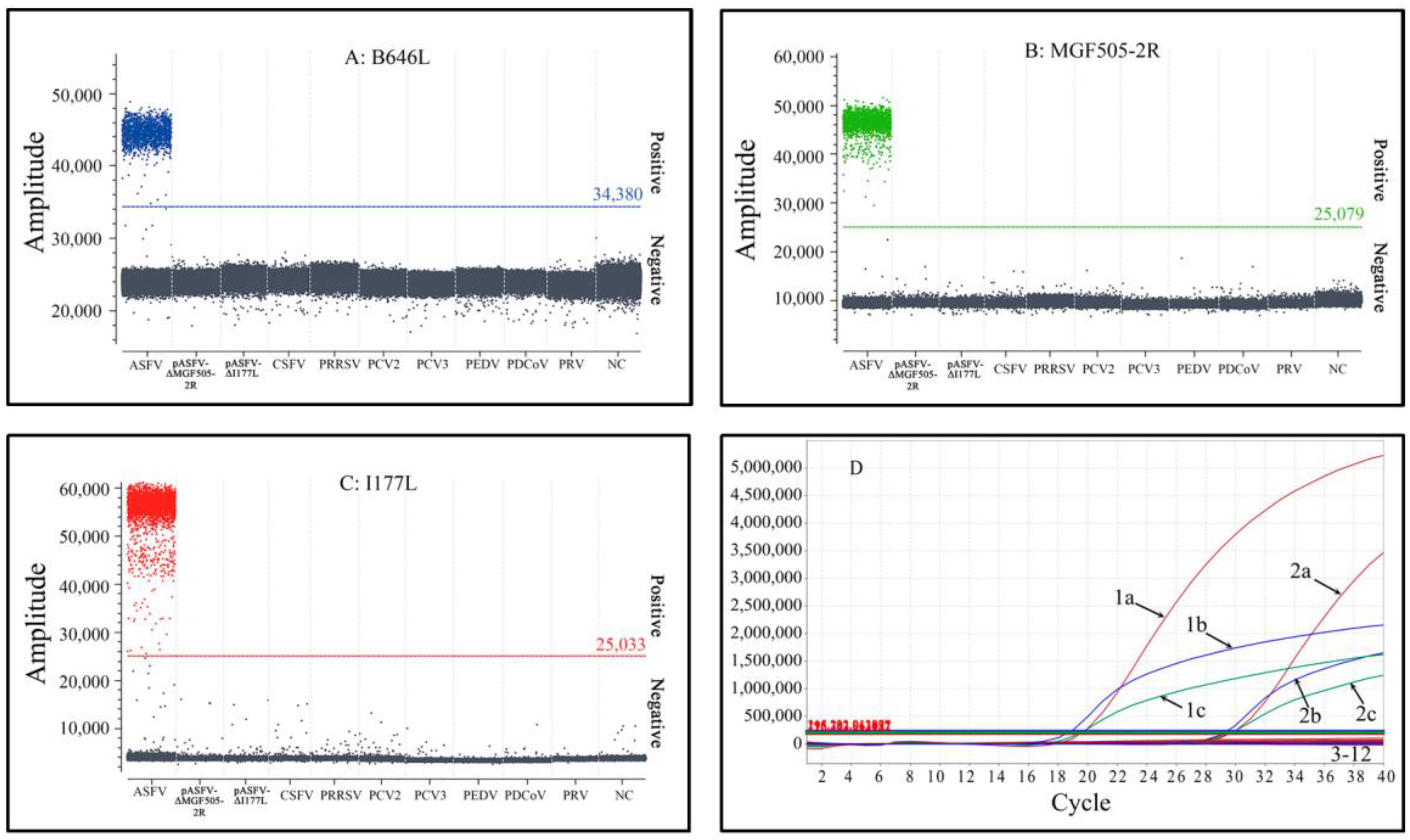
3.5. Specificity Analysis
3.6. Repeatability and Reproducibility Analysis
3.7. Application for the Detection of Clinical Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Li, X.; Zhao, M.; Ding, H.; Fan, S.; et al. African Swine Fever Virus: A Review. Life 2022, 12, 1255. [Google Scholar] [CrossRef] [PubMed]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; ICTV Report Consortium. ICTV Virus Taxonomy Profile Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef]
- Karger, A.; Pérez-Núñez, D.; Urquiza, J.; Hinojar, P.; Alonso, C.; Freitas, F.B.; Revilla, Y.; Le Potier, M.-F.; Montoya, M. An Update on African Swine Fever Virology. Viruses 2019, 11, 864. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Xie, M.; Wu, W.; Chen, Z. Structures and Functional Diversities of ASFV Proteins. Viruses 2021, 13, 2124. [Google Scholar] [CrossRef]
- Cwynar, P.; Stojkov, J.; Wlazlak, K. African Swine Fever Status in Europe. Viruses 2019, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Martínez-López, B. African Swine Fever: An Epidemiological Update. Transbound. Emerg. Dis. 2012, 59 (Suppl. S1), 27–35. [Google Scholar] [CrossRef]
- Danzetta, M.L.; Marenzoni, M.L.; Iannetti, S.; Tizzani, P.; Calistri, P.; Feliziani, F. African Swine Fever: Lessons to Learn From Past Eradication Experiences. A Systematic Review. Front. Veter-Sci. 2020, 7, 296. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African Swine Fever Virus: An Emerging DNA Arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef]
- Tao, D.; Sun, D.; Liu, Y.; Wei, S.; Yang, Z.; An, T.; Shan, F.; Chen, Z.; Liu, J. One year of African swine fever outbreak in China. Acta Trop. 2020, 211, 105602. [Google Scholar] [CrossRef]
- Ito, S.; Bosch, J.; Martínez-Avilés, M.; Sánchez-Vizcaíno, J.M. The evolution of African swine fever in China: A global threat? Front. Vet. Sci. 2022, 9, 828498. [Google Scholar] [CrossRef]
- Mighell, E.; Ward, M.P. African Swine Fever spread across Asia, 2018–2019. Transbound. Emerg. Dis. 2021, 68, 2722–2732. [Google Scholar] [CrossRef] [PubMed]
- Cadenas-Fernández, E.; Ito, S.; Aguilar-Vega, C.; Sánchez-Vizcaíno, J.M.; Bosch, J. The Role of the Wild Boar Spreading African Swine Fever Virus in Asia: Another Underestimated Problem. Front. Vet. Sci. 2022, 9, 844209. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef] [PubMed]
- Penrith, M.-L.; Van Heerden, J.; Heath, L.; Abworo, E.O.; Bastos, A.D.S. Review of the Pig-Adapted African Swine Fever Viruses in and Outside Africa. Pathogens 2022, 11, 1190. [Google Scholar] [CrossRef]
- Njau, E.P.; Domelevo Entfellner, J.-B.; Machuka, E.M.; Bochere, E.N.; Cleaveland, S.; Shirima, G.M.; Kusiluka, L.J.; Upton, C.; Bishop, R.P.; Pelle, R.; et al. The first genotype II African swine fever virus isolated in Africa provides insight into the current Eurasian pandemic. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Shi, K.; Liu, H.; Yin, Y.; Si, H.; Long, F.; Feng, S. Molecular Characterization of African Swine Fever Virus From 2019-2020 Outbreaks in Guangxi Province, Southern China. Front. Vet. Sci. 2022, 9, 912224. [Google Scholar] [CrossRef]
- Qu, H.; Ge, S.; Zhang, Y.; Wu, X.; Wang, Z. A systematic review of genotypes and serogroups of African swine fever virus. Virus Genes 2022, 58, 77–87. [Google Scholar] [CrossRef]
- Sun, E.; Huang, L.; Zhang, X.; Zhang, J.; Shen, D.; Zhang, Z.; Wang, Z.; Huo, H.; Wang, W.; Huangfu, H.; et al. Genotype I African swine fever viruses emerged in domestic pigs in China and caused chronic infection. Emerg. Microbes Infect. 2021, 10, 2183–2193. [Google Scholar] [CrossRef]
- Liu, H.; Shi, K.; Zhao, J.; Yin, Y.; Chen, Y.; Si, H.; Qu, S.; Long, F.; Lu, W. Development of a one-step multiplex qRT-PCR assay for the detection of African swine fever virus, classical swine fever virus and atypical porcine pestivirus. BMC Vet. Res. 2022, 18, 43. [Google Scholar] [CrossRef]
- Sun, E.; Zhang, Z.; Wang, Z.; He, X.; Zhang, X.; Wang, L.; Wang, W.; Huang, L.; Xi, F.; Huangfu, H.; et al. Emergence and prevalence of naturally occurring lower virulent African swine fever viruses in domestic pigs in China in 2020. Sci. China Life Sci. 2021, 64, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Qi, C.; Ge, S.; Li, J.; Hu, Y.; Zhang, X.; Lv, Y.; Han, N.; Wu, X.; Wang, Z.; et al. Genetic variation and evolution of attenuated African swine fever virus strain isolated in the field: A review. Virus Res. 2022, 319, 198874. [Google Scholar]
- Chen, W.; Zhao, D.; He, X.; Liu, R.; Wang, Z.; Zhang, X.; Li, F.; Shan, D.; Chen, H.; Zhang, J.; et al. A seven-gene-deleted African swine fever virus is safe and effective as a live attenuated vaccine in pigs. Sci. China Life Sci. 2020, 63, 623–634. [Google Scholar] [CrossRef]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African Swine Fever Virus Georgia Isolate Harboring Deletions of MGF360 and MGF505 Genes Is Attenuated in Swine and Confers Protection against Challenge with Virulent Parental Virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef]
- Monteagudo, P.L.; Lacasta, A.; López, E.; Bosch, L.; Collado, J.; Pina-Pedrero, S.; Correa-Fiz, F.; Accensi, F.; Navas, M.J.; Vidal, E.; et al. BA71ΔCD2: A New Recombinant Live Attenuated African Swine Fever Virus with Cross-Protective Capabilities. J. Virol. 2017, 91, e01058-17. [Google Scholar] [CrossRef]
- Lopez, E.; Bosch-Camós, L.; Ramirez-Medina, E.; Vuono, E.; Navas, M.J.; Muñoz, M.; Accensi, F.; Zhang, J.; Alonso, U.; Argilaguet, J.; et al. Deletion Mutants of the Attenuated Recombinant ASF Virus, BA71ΔCD2, Show Decreased Vaccine Efficacy. Viruses 2021, 13, 1678. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Gay, C.G.; Gladue, D.P. ASFV-G-∆I177L as an Effective Oral Nasal Vaccine against the Eurasia Strain of Africa Swine Fever. Viruses 2021, 13, 765. [Google Scholar] [CrossRef]
- Tran, X.H.; Le, T.T.P.; Nguyen, Q.H.; Do, T.T.; Nguyen, V.D.; Gay, C.G.; Borca, M.V.; Gladue, D.P. African swine fever virus vaccine candidate ASFV-G-DeltaI177L efficiently protects European and native pig breeds against circulating Vietnamese field strain. Transbound. Emerg. Dis. 2022, 69, e497–e504. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, Z.; Li, Y.; Song, Y.; Di, D.; Liu, J.; Gong, L.; Chen, Z.; Wu, J.; Ye, Z.; et al. Evaluation of an I177L gene-based five-gene-deleted African swine fever virus as a live attenuated vaccine in pigs. Emerg. Microbes Infect. 2023, 12, 2148560. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, S.; Zhang, H.; Qin, Z.; Shan, H.; Cai, X. Vaccines for African swine fever: An update. Front. Microbiol. 2023, 14, 1139494. [Google Scholar] [CrossRef]
- Hawkins, S.F.C.; Guest, P.C. Multiplex Analyses Using Real-Time Quantitative PCR. Methods Mol. Biol. 2017, 1546, 125–133. [Google Scholar] [CrossRef]
- Kuypers, J.; Jerome, K.R. Applications of Digital PCR for Clinical Microbiology. J. Clin. Microbiol. 2017, 55, 1621–1628. [Google Scholar] [CrossRef] [PubMed]
- Hindson, C.M.; Chevillet, J.R.; Briggs, H.A.; Gallichotte, E.N.; Ruf, I.K.; Hindson, B.J.; Vessella, R.L.; Tewari, M. Absolute quantification by droplet digital PCR versus analog real-time PCR. Nat. Methods 2013, 10, 1003–1005. [Google Scholar] [CrossRef]
- Wu, X.; Xiao, L.; Lin, H.; Chen, S.; Yang, M.; An, W.; Wang, Y.; Yang, Z.; Yao, X.; Tang, Z. Development and application of a droplet digital polymerase chain reaction (ddPCR) for detection and investigation of African swine fever virus. Can. J. Vet. Res. 2018, 82, 70–74. [Google Scholar] [PubMed]
- Zhu, J.; Jian, W.; Huang, Y.; Gao, Q.; Gao, F.; Chen, H.; Zhang, G.; Liao, M.; Qi, W. Development and Application of a Duplex Droplet Digital Polymerase Chain Reaction Assay for Detection and Differentiation of EP402R-Deleted and Wild-Type African Swine Fever Virus. Front. Vet. Sci. 2022, 9, 905706. [Google Scholar] [CrossRef]
- Shi, K.; Chen, Y.; Yin, Y.; Long, F.; Feng, S.; Liu, H.; Qu, S.; Si, H. A Multiplex Crystal Digital PCR for Detection of African Swine Fever Virus, Classical Swine Fever Virus, and Porcine Reproductive and Respiratory Syndrome Virus. Front. Vet. Sci. 2022, 9, 926881. [Google Scholar] [CrossRef]
- Jia, R.; Zhang, G.; Liu, H.; Chen, Y.; Zhou, J.; Liu, Y.; Ding, P.; Wang, Y.; Zang, W.; Wang, A. Novel Application of Nanofluidic Chip Digital PCR for Detection of African Swine Fever Virus. Front. Vet. Sci. 2020, 7, 621840. [Google Scholar] [CrossRef]
- Wang, A.; Jia, R.; Liu, Y.; Zhou, J.; Qi, Y.; Chen, Y.; Liu, D.; Zhao, J.; Shi, H.; Zhang, J.; et al. Development of a novel quantitative real-time PCR assay with lyophilized powder reagent to detect African swine fever virus in blood samples of domestic pigs in China. Transbound. Emerg. Dis. 2020, 67, 284–297. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, L.; Noll, L.; Stoy, C.; Porter, E.; Fu, J.; Feng, Y.; Peddireddi, L.; Liu, X.; Dodd, K.A.; et al. Development of a real-time PCR assay for detection of African swine fever virus with an endogenous internal control. Transbound. Emerg. Dis. 2020, 67, 2446–2454. [Google Scholar] [CrossRef]
- Zhan, Y.; Zhang, L.-H.; Lin, Y.; Cai, Y.-F.; Zou, Y.-W.; Hao, Z.-Y.; Luo, Z.-H.; Wang, N.-D.; Deng, Z.-B.; Yang, Y.; et al. Development and preliminary testing of a probe-based duplex real-time PCR assay for the detection of African swine fever virus. Mol. Cell. Probes 2021, 59, 101764. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, Y.; Liu, P.; Zhu, Z.; Liu, P.; Chen, C.; Wu, X. Development and application of a duplex real-time PCR assay for differentiation of genotypes I and II African swine fever viruses. Transbound. Emerg. Dis. 2022, 69, 2971–2979. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Lu, H.; Wu, Z.; Zhu, S. A duplex fluorescent quantitative PCR assay to distinguish the genotype I and II strains of African swine fever virus in Chinese epidemic strains. Front. Vet. Sci. 2022, 9, 998874. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Feng, Y.; Yang, Y.; Luo, Y.; Gong, T.; Wang, H.; Gong, L.; Zhang, G.; Zheng, Z. Establishment of a Dual Real-Time PCR Assay for the Identification of African Swine Fever Virus Genotypes I and II in China. Front. Vet. Sci. 2022, 9, 882824. [Google Scholar] [CrossRef]
- Yang, H.; Peng, Z.; Song, W.; Zhang, C.; Fan, J.; Chen, H.; Hua, L.; Pei, J.; Tang, X.; Chen, H.; et al. A triplex real-time PCR method to detect African swine fever virus gene-deleted and wild type strains. Front. Vet. Sci. 2022, 9, 943099. [Google Scholar] [CrossRef]
- Zhao, K.; Shi, K.; Zhou, Q.; Xiong, C.; Mo, S.; Zhou, H.; Long, F.; Wei, H.; Hu, L.; Mo, M. The Development of a Multiplex Real-Time Quantitative PCR Assay for the Differential Detection of the Wild-Type Strain and the MGF505-2R, EP402R and I177L Gene-Deleted Strain of the African Swine Fever Virus. Animals 2022, 12, 1754. [Google Scholar] [CrossRef]
- Lin, Y.; Cao, C.; Shi, W.; Huang, C.; Zeng, S.; Sun, J.; Wu, J.; Hua, Q. Development of a triplex real-time PCR assay for detection and differentiation of gene-deleted and wild-type African swine fever virus. J. Virol. Methods 2020, 280, 113875. [Google Scholar] [CrossRef]
- Guo, Z.; Li, K.; Qiao, S.; Chen, X.-X.; Deng, R.; Zhang, G. Development and evaluation of duplex TaqMan real-time PCR assay for detection and differentiation of wide-type and MGF505-2R gene-deleted African swine fever viruses. BMC Vet. Res. 2020, 16, 428. [Google Scholar] [CrossRef]
- Ge, S.; Li, J.; Fan, X.; Liu, F.; Li, L.; Wang, Q.; Ren, W.; Bao, J.; Liu, C.; Wang, H.; et al. Molecular Characterization of African Swine Fever Virus, China, 2018. Emerg. Infect. Dis. 2018, 24, 2131–2133. [Google Scholar] [CrossRef]
- Ata, E.B.; Li, Z.-J.; Shi, C.-W.; Yang, G.-L.; Yang, W.-T.; Wang, C.-F. African swine fever virus: A raised global upsurge and a continuous threaten to pig husbandry. Microb. Pathog. 2022, 167, 105561. [Google Scholar] [CrossRef]
- Madic, J.; Zocevic, A.; Senlis, V.; Fradet, E.; Andre, B.; Muller, S.; Dangla, R.; Droniou, M.E. Three-color crystal digital PCR. Biomol. Detect. Quantif. 2016, 10, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.L.; Loganathan, N.; Agarwalla, S.; Yang, C.; Yuan, W.; Zeng, J.; Wu, R.; Wang, W.; Duraiswamy, S. Current commercial dPCR platforms: Technology and market review. Crit. Rev. Biotechnol. 2023, 43, 433–464. [Google Scholar] [CrossRef]
- Whale, A.S.; Cowen, S.; Foy, C.A.; Huggett, J.F. Methods for Applying Accurate Digital PCR Analysis on Low Copy DNA Samples. PLoS ONE 2013, 8, e58177. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shi, K.; Liu, H.; Yin, Y.; Zhao, J.; Long, F.; Lu, W.; Si, H. Development of a multiplex qRT-PCR assay for detection of African swine fever virus, classical swine fever virus and porcine reproductive and respiratory syndrome virus. J. Vet. Sci. 2021, 22, e87. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Qi, W.; Yang, Y.; Liu, Z.; An, T.; Wu, X.; Chen, J. Prevention and Control Strategies of African Swine Fever and Progress on Pig Farm Repopulation in China. Viruses 2021, 13, 2552. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Stahl, K.; Jori, F.; Vial, L.; Pfeiffer, D.U. African Swine Fever Epidemiology and Control. Annu. Rev. Anim. Biosci. 2020, 8, 221–246. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeted Gene | Name | Sequences (5′→3′) | Amplicon (bp) |
|---|---|---|---|
| B646L | ASFV-B646L-F | GGCGTATAAAAAGTCCAGGAAATTC | 79 |
| ASFV-B646L-R | TTCGGCGAGCGCTTTATC | ||
| ASFV-B646L-P | FAM-TCACCAAATCCTTTTGCGATGCAAGCT-BHQ1 | ||
| MGF505-2R | ASFV-MGF505-F | AGTCATGCACGGCATATACAA | 153 |
| ASFV-MGF505-R | GGTTTAAACCGTGCCACATCC | ||
| ASFV-MGF505-P | VIC-ACGCGGCCACCCAATTCAGAGAC-BHQ1 | ||
| I177L | ASFV-I177L-F | GGCATAATTATCAAATGCGAAGGG | 122 |
| ASFV-I177L-R | TGGAAAGTTAATGATCAGGGCTT | ||
| ASFV-I177L-P | Cy5-AATCCTAGCTTGCCGGTAATGGCT-BHQ2 |
| Reagent | Triplex cdPCR | Triplex qPCR | ||
|---|---|---|---|---|
| Volume (μL) | Final Concentration (nM) | Volume (μL) | Final Concentration (nM) | |
| PerfeCta Multiplex qPCR ToughMix (2×) | 12.5 | 1× | / | / |
| Fluorescein Sodium Salt (1 µM) | 2.5 | 100 | / | / |
| Premix Ex Taq (Probe qPCR) (2×) | / | / | 12.5 | 1× |
| ASFV-B646L-F (25 µM) | 0.8 | 800 | 0.3 | 300 |
| ASFV-B646L-R (25 µM) | 0.8 | 800 | 0.3 | 300 |
| ASFV-B646L-P (25 µM) | 0.3 | 300 | 0.3 | 300 |
| ASFV-MGF505-2R-F (25 µM) | 0.8 | 800 | 0.4 | 400 |
| ASFV-MGF505-2R-R (25 µM) | 0.8 | 800 | 0.4 | 400 |
| ASFV-MGF505-2R-P (25 µM) | 0.3 | 300 | 0.4 | 400 |
| ASFV-I177L-F (25 µM) | 0.8 | 800 | 0.4 | 400 |
| ASFV-I177L-R (25 µM) | 0.8 | 800 | 0.4 | 400 |
| ASFV-I177L-P (25 µM) | 0.3 | 300 | 0.4 | 400 |
| Total nucleic acids | 2.5 | / | 2.5 | / |
| Nuclease-free distilled water | Up to 25 | / | Up to 25 | / |
| Plasmid | Concentration (Copies/μL) | Intra-Assay for Repeatability | Inter-Assay for Reproducibility | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Triplex cdPCR (Copies/Reaction) | Triplex qPCR (Ct) | Triplex cdPCR (Copies/Reaction) | Triplex qPCR (Ct) | ||||||||||
| SD | CV (%) | SD | CV (%) | SD | CV (%) | SD | CV (%) | ||||||
| pASFV-B646L-1 | 2.0 × 104 | 33,983.33 | 500.83 | 1.47 | 25.33 | 0.19 | 0.75 | 34,083.33 | 780.36 | 2.29 | 25.46 | 0.27 | 1.06 |
| 2.0 × 103 | 3463.33 | 41.26 | 1.19 | 28.68 | 0.23 | 0.80 | 3455.00 | 56.35 | 1.63 | 28.48 | 0.40 | 1.40 | |
| 2.0 × 102 | 331.67 | 6.29 | 1.90 | 32.20 | 0.36 | 1.12 | 335.00 | 9.01 | 2.69 | 32.07 | 0.59 | 1.84 | |
| pASFV-MGF505-2R-1 | 2.0 × 104 | 43,225.00 | 650.00 | 1.50 | 24.44 | 0.17 | 0.70 | 43,418.33 | 698.75 | 1.61 | 24.38 | 0.23 | 0.94 |
| 2.0 × 103 | 4389.17 | 68.66 | 1.56 | 28.38 | 0.20 | 0.70 | 4370.83 | 91.66 | 2.10 | 28.68 | 0.30 | 1.05 | |
| 2.0 × 102 | 390.83 | 5.20 | 1.33 | 31.53 | 0.31 | 0.98 | 395.00 | 10.00 | 2.53 | 32.03 | 0.52 | 1.62 | |
| pASFV-I177L-1 | 2.0 × 104 | 54,550.00 | 912.41 | 1.67 | 23.66 | 0.21 | 0.89 | 54,041.67 | 1179.07 | 2.18 | 23.56 | 0.30 | 1.27 |
| 2.0 × 103 | 5607.50 | 90.93 | 1.62 | 28.28 | 0.24 | 0.85 | 5612.50 | 103.11 | 1.84 | 28.28 | 0.24 | 0.85 | |
| 2.0 × 102 | 515.00 | 5.00 | 0.97 | 31.49 | 0.18 | 0.57 | 509.17 | 11.27 | 2.21 | 31.69 | 0.20 | 0.63 | |
| Pathogen | Number | Triplex qPCR | Triplex cdPCR | Coincidence Rate (%) | Kappa | ||
|---|---|---|---|---|---|---|---|
| Positive | Positive Rate (%) | Positive | Positive Rate (%) | ||||
| ASFV | 1510 | 196 | 12.98 | 214 | 14.17 | 98.81 | 0.95 |
| ASFV-ΔMGF505-2R | 1510 | 5 | 0.33 | 5 | 0.33 | 100 | / |
| ASFV-ΔI177L | 1510 | 0 | 0 | 0 | 0 | 100 | / |
| Triplex qPCR | Triplex cdPCR | Coincidence Rate (%) | Kappa | ||
|---|---|---|---|---|---|
| Positive | Negative | Total | |||
| Positive | 196 | 0 | 196 | 98.81 | 0.95 |
| Negative | 18 | 1296 | 1314 | ||
| Total | 214 | 1296 | 1510 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, K.; Zhao, K.; Wei, H.; Zhou, Q.; Shi, Y.; Mo, S.; Long, F.; Hu, L.; Feng, S.; Mo, M. Triplex Crystal Digital PCR for the Detection and Differentiation of the Wild-Type Strain and the MGF505-2R and I177L Gene-Deleted Strain of African Swine Fever Virus. Pathogens 2023, 12, 1092. https://doi.org/10.3390/pathogens12091092
Shi K, Zhao K, Wei H, Zhou Q, Shi Y, Mo S, Long F, Hu L, Feng S, Mo M. Triplex Crystal Digital PCR for the Detection and Differentiation of the Wild-Type Strain and the MGF505-2R and I177L Gene-Deleted Strain of African Swine Fever Virus. Pathogens. 2023; 12(9):1092. https://doi.org/10.3390/pathogens12091092
Chicago/Turabian StyleShi, Kaichuang, Kang Zhao, Haina Wei, Qingan Zhou, Yuwen Shi, Shenglan Mo, Feng Long, Liping Hu, Shuping Feng, and Meilan Mo. 2023. "Triplex Crystal Digital PCR for the Detection and Differentiation of the Wild-Type Strain and the MGF505-2R and I177L Gene-Deleted Strain of African Swine Fever Virus" Pathogens 12, no. 9: 1092. https://doi.org/10.3390/pathogens12091092
APA StyleShi, K., Zhao, K., Wei, H., Zhou, Q., Shi, Y., Mo, S., Long, F., Hu, L., Feng, S., & Mo, M. (2023). Triplex Crystal Digital PCR for the Detection and Differentiation of the Wild-Type Strain and the MGF505-2R and I177L Gene-Deleted Strain of African Swine Fever Virus. Pathogens, 12(9), 1092. https://doi.org/10.3390/pathogens12091092