1. Introduction
Hantavirus cardiopulmonary syndrome (HCPS) is one of the zoonotic viral diseases caused by the family of viruses of the order
Bunyavirales, within which are the Sin Nombre virus (SNV) and the Andes virus (ANDV) [
1], which are the most common zoonotic agents that cause HCPS and are found mainly in rodents such as
Peromyscus maniculatus (deer mouse) or
Oligoryzomys longicaudatus (long-tailed mouse) [
2,
3]. Only in Argentina and Chile are HCPS associated with ANDV infection through the long-tailed mouse [
2]. Because of the genus
Hantavirus, in addition to HCPS, other hantaviruses can cause hemorrhagic fever with renal syndrome (HFRS).
HCPS occurs mainly in the Americas, while HFRS occurs in Asia and Europe [
4]. HCPS was discovered in the United States in 1993, when SNV was identified, with an initial mortality of 50%, and to date there is no cure [
2,
5,
6]. At first, it was called hantavirus pulmonary syndrome; however, it was redefined as hantavirus cardiopulmonary syndrome because the leading cause of death is myocardial depression [
7].
HCPS has a mortality rate of
of cases infected by ANDV [
8], with the lethality of this hantavirus disease between 30 and
[
5]. The high lethality can be correlated with the short intervention period since, on average, one has three days between the first symptoms and the first consultation, and two more days between the first consultation and death; that is to say, it is very brief [
6]. Furthermore, there is a relationship between socioeconomic level and this indicator, since more significant economic vulnerability increases the probability of death [
5].
Transmission of the virus from rodent to humans occurs primarily through inhalation of viral particles found in the fluids of this infected rodent, such as urine, faeces, or saliva [
9,
10]. This contagion occurs in work activities (particularly forest workers and farmers), or recreational or domestic scenarios [
2,
11,
12,
13]. There are also other types of infection, but these are more isolated cases, such as bites or by eating an infected rodent. There are cases of person-to-person transmission, occurring mainly in Argentina and Chile [
9,
14,
15,
16]. Still, these have been isolated cases, except for what happened in Argentina between November 2018 and February 2019, resulting in 34 confirmed cases of contagion of the virus, ANDV among people, and 11 deaths [
15].
The number of people infected each year varies; this can occur due to the ecosystem variations of recent years produced by climate change, such as forest fires, among others, increasing the frequency of high-impact events [
2]. In addition, the flowering of the
Chusquea quila and the
Chusquea colihue (the main food of the rodent) has boosted the increase in the reservoir population [
17].
The countries in America with the highest incidence of hantavirus infection (HI) correspond to Brazil, Argentina, and Chile [
2]. In Argentina and Chile, the main reservoirs are found among mice:
Oligoryzomys longicaudatus, Abrothrix olivaceus, and
Akodon longipilis; the first of these was found with a more infected population in these two mentioned countries [
18]. It should be noted that in Argentina in 2021 a new reservoir was found,
Scapteromys aquaticus, further expanding the diversity of reservoirs [
19]. In Brazil, there are many hantavirus reservoirs;
Oligoryzomys nigripi and
Necromys lasiurus are some of them [
20].
Most mathematical models study the dynamics of the hantavirus among rodents [
21,
22,
23,
24,
25,
26]. Some distinguish between males and females [
21,
23]; others compare direct with indirect transmission, involving demographic, environmental, or seasonal variables [
24,
25,
27]; and in [
28], they predict the territorial distribution of infected rodents. However, there are models focused on cases of HI in humans; for example, in [
29], the human population is divided into agricultural workers and others, while in [
30], with a statistical approach, the article projects possible future cases according to the environmental variable. Our study presents a mathematical-epidemiological generalist model representing the territorial dynamics between humans and rodents (main novelty). It aims to analyze this territorial distribution impact on the disease (HI) spread in the human co-inhabiting population.
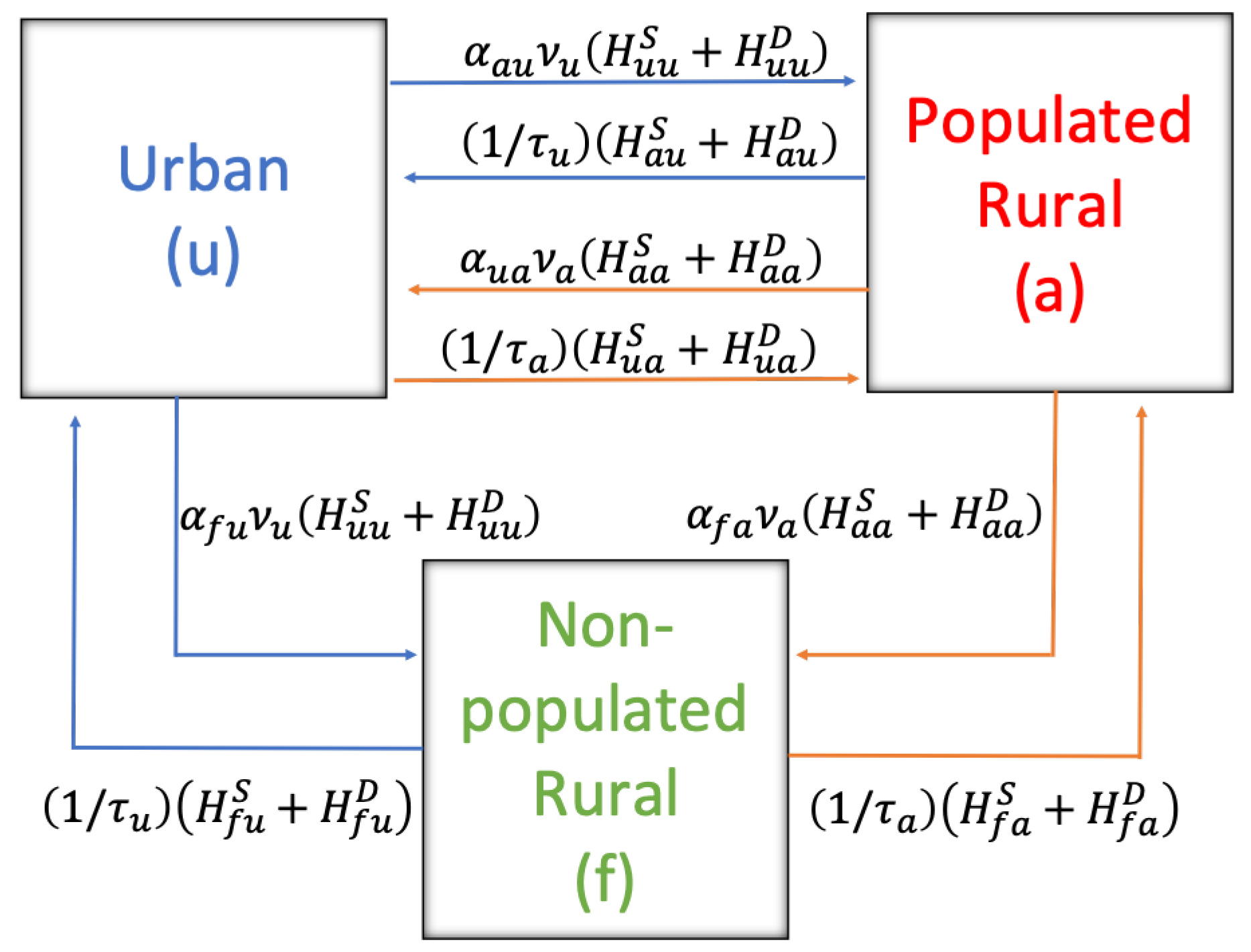
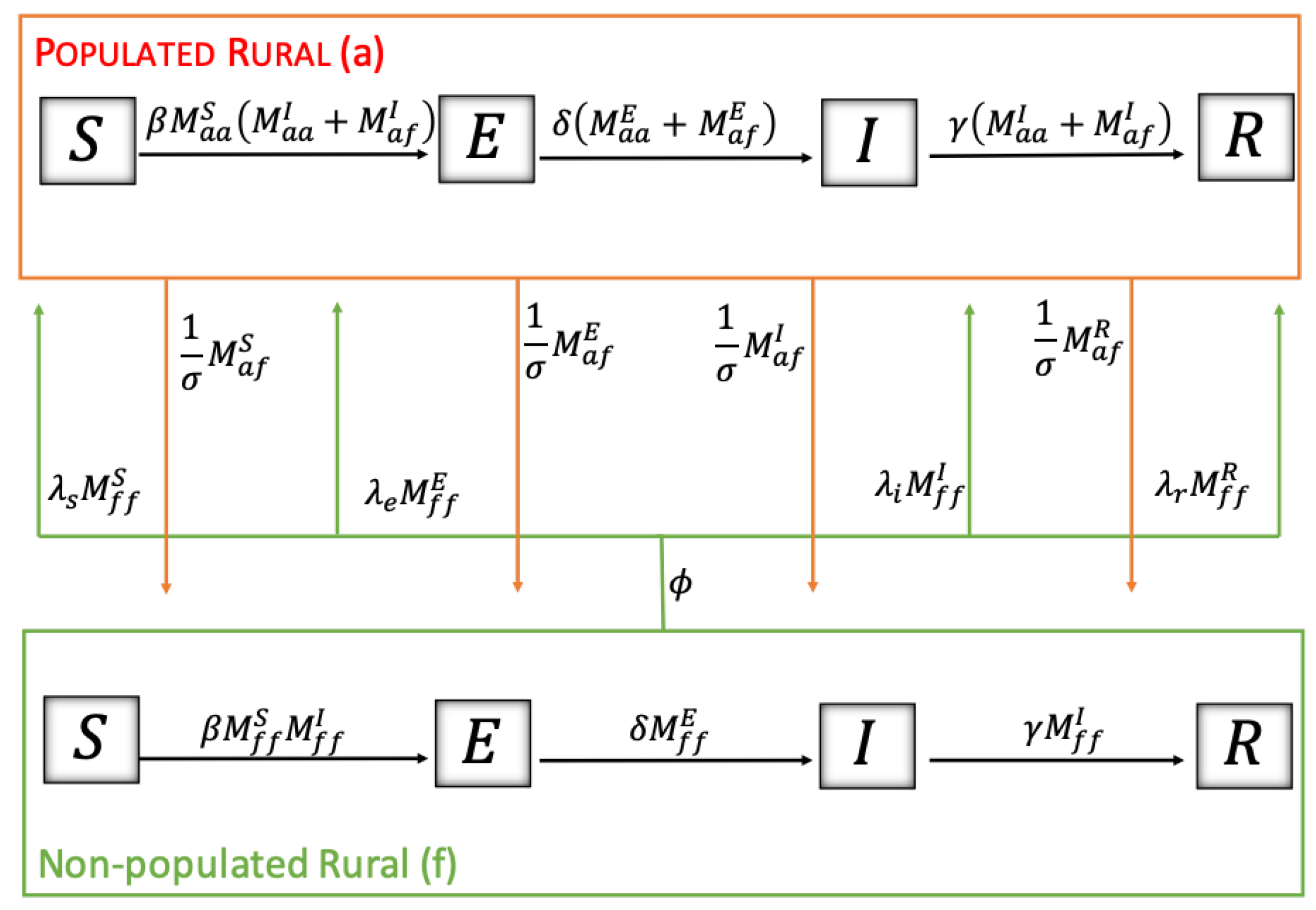
To express the dynamics of the disease among rodents, we rely on a compartmental model type SEIR (Susceptible-Exposed-Infectious-Removed) [
31]. At the same time, for the sanitary condition of humans toward infection, there are two states: susceptible or infected (non-infectious); although there are reported cases of contagion among people, these are particular cases and more studies are needed, so for simplicity of the model, transmission between people will not be considered, only their mobility.
Next, relevant data on HI propagation will be presented (
Section 2.1), as well as the mathematical model of HI together with the study of the proposed system (
Section 2.2), different numerical simulations based on the exposed model (
Section 3), and finally (
Section 4), the conclusions obtained from the research will be discussed.
3. Results
To express the dynamics of the model presented, the numerical simulations (using the ode45 function of MATLAB) will be carried out with the data associated with Chile, particularly with the Maule region, whose total population, according to the 2017 census, corresponds to 1,044,950 inhabitants, where approximately 70% of the population lives in the urban sector and 30% in the rural sector [
35].
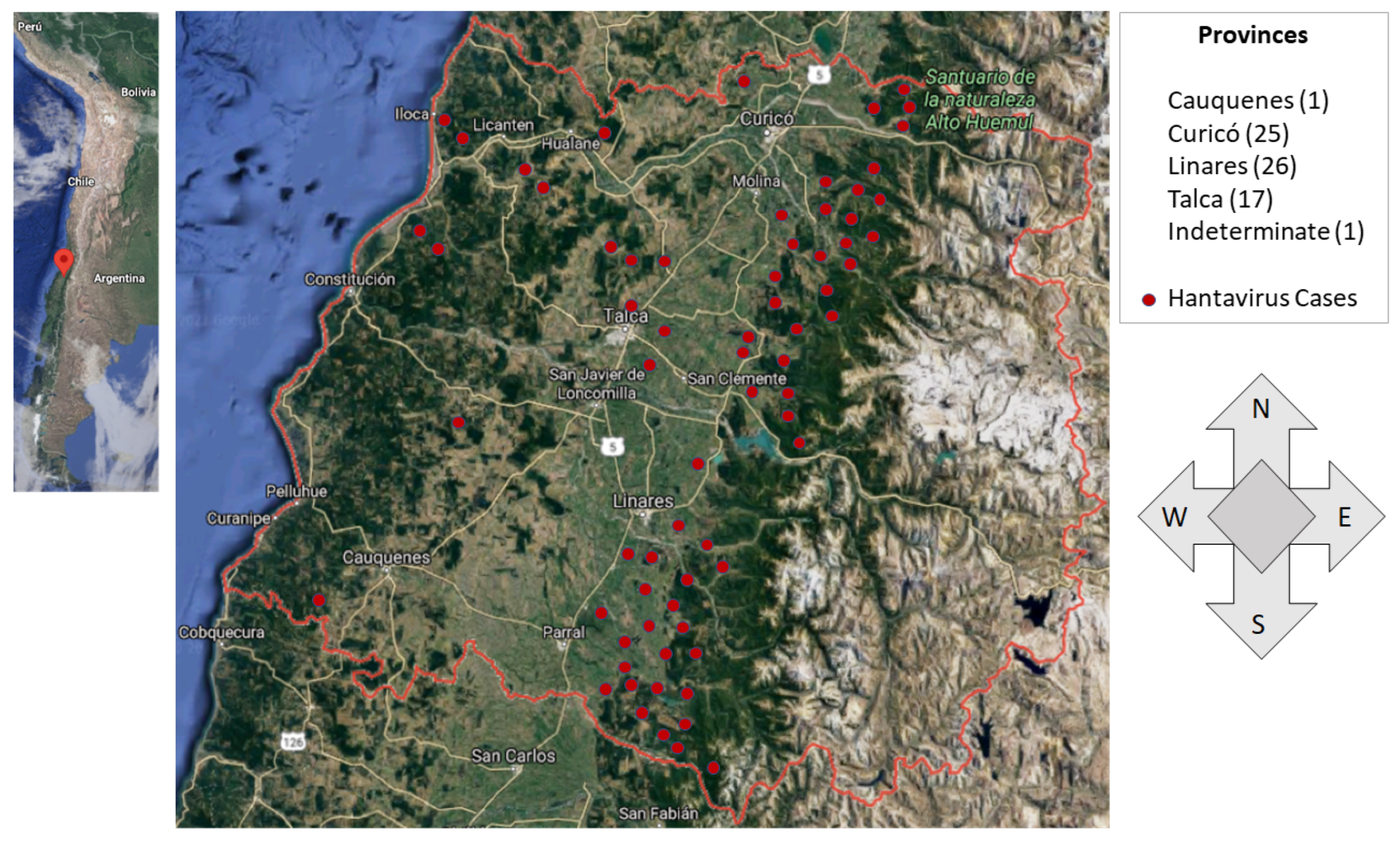
According to the data provided by the SEREMI of Health Maule region [
9], a region located in south-central Chile, we can classify the residence and the place of infection among the three sectors of our model: Urban (
u), Rural populated (
a), and Rural non-populated (
f), which shows that the greatest cases occur among people who live in the rural populated sector (61%), followed by people who live in the urban sector and are infected in the rural populated (23%). They are followed by people living in the rural populated and urban sectors who are infected in the rural unpopulated sector, with 12% and 4%, respectively; see
Figure 5.
The numerical values associated with the different rates presented in the model have been extracted from other investigations [
21,
23,
26,
29]. Based on the information provided in
Figure 5 (see
Table 6), these have been chosen to obtain a projection treating to incorporate more realism.
Thus, with the chosen rates, we obtain that (see
Figure 6) the percentage of the human population infected by HI during a year amounts to
, that is, approximately 0.0012% of the population, which is expressed in the Maule region as a total of 12 infected people (value according to the data provided in
Table 4), and which, when distributed among the sectors of the model, is 58, 25, 11, and 6 (%) for
,
,
, and
, respectively (values very close to those provided in
Figure 5). Mortality corresponds to
, which is equivalent to 30% of the infected population (four cases).
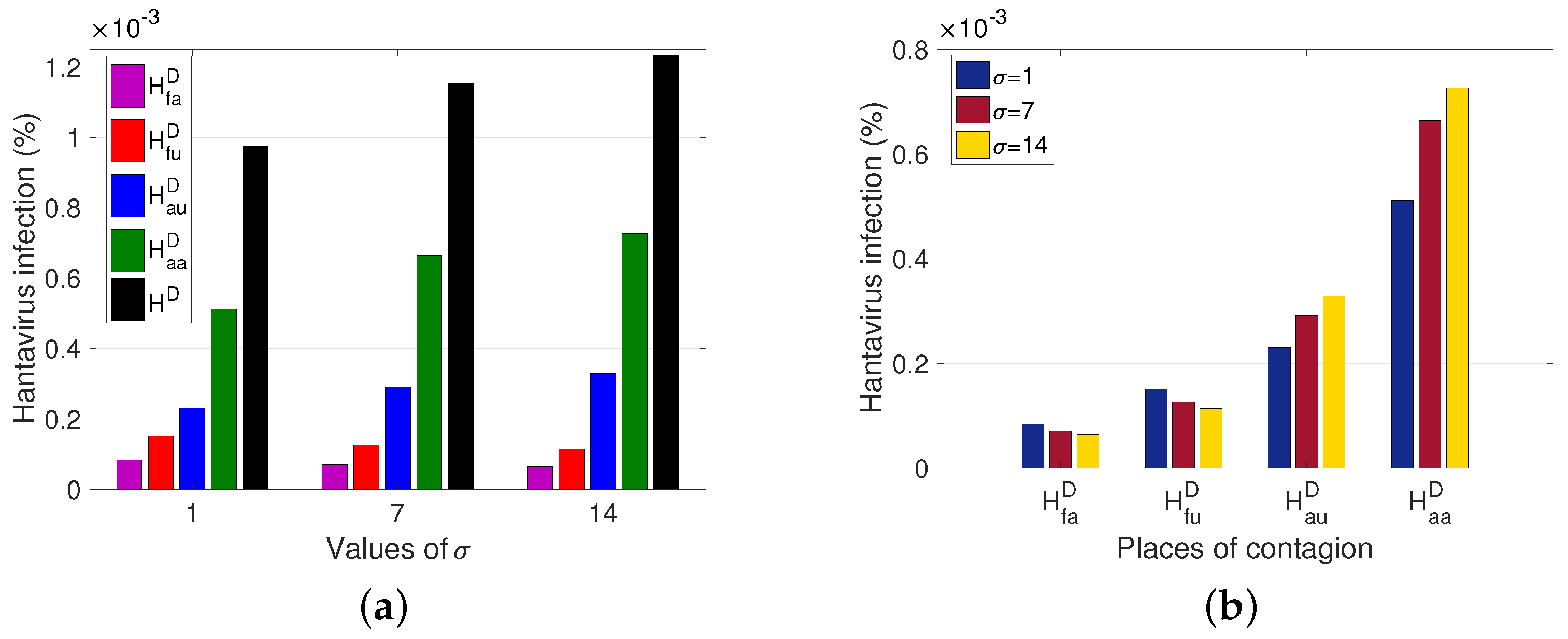
In what follows, we proceed to visualize the impact that mobility parameters have on the development of the disease.
The average time that a rodent from the non-populated rural sector remains in the populated rural sector (
) is a factor to consider. The longer the rodent stays as an outsider, the more HI cases increase for people living in (
) or traveling to the rural sector (
), while the cases associated with infection in the rural sector (
and
) decrease (see
Figure 7b). It is also observed that the total number of HI cases increases by approximately
of the population, after the increase of
between 1 and 14 days (see
Figure 7a).
Another relevant factor is the proportion of the rodent population living in the unpopulated rural sector that goes into the populated rural sector (
).
Figure 8 shows an effect similar to that of
Figure 7, since although there is a decrease in cases in the non-populated rural sector, the number of cases in the populated rural sector increases, and the total number of cases increases.
In addition to the dynamics of the reservoir’s movement, it is relevant to observe the effect that the movement of the human has in different scenarios by varying the average time spent as an outsider between the different sectors. Three cases are presented; the first corresponds to people who leave their sector and stay eight hours in another sector, that is,
(where
expresses that
), followed by the second case, where they stay 5 days (from Monday to Friday), and finally, the third case, which is 7 days (full week). From
Figure 9a, one can see the significant increase in the total number of HI cases in the population, after the variation of
, increased by approximately
of the population. Concerning the number of HI cases per sector, we explicitly observe how the number of cases is altered after the variation of
(see
Figure 9b).
4. Discussion and Conclusions
The dynamics of HI transmission were modeled by ordinary differential equations, incorporating the mobility of humans and rodents in three sectors: urban, populated rural, and non-populated rural, the latter two being the territories where infection occurs. To build the model, we have relied on previous compartmental models that study these dynamics without territorial mobility, the main novelty of our work. The data that feed our model were mainly from the cases reported in Chile and other studies previously carried out in different parts of the world. The scarcity of mobility information is one of the main limitations. However, the generality of our model can provide considerable qualitative results that, in a novel way, consider the mobility of humans and rodents, contributing to the literature and informing the guidelines in public health decision-making.
From the background consulted on the presence of the lethal HI disease in the Maule region and based on the existing literature, we mathematically modeled the territorial dynamics between humans and rodents, evidencing the impact on the spread of HI that can occur with increased human mobility.
Several variables are identified in the recent literature associated with the prevalence of HI and its infection through the rodent. For example, forest fires, high temperatures, and droughts are identified as precipitating factors for the increased prevalence of zoonoses [
39,
40]. A study conducted in a boreal forest in Sweden, which suffered a large-scale fire in 2006, determined a high risk of infection of Puumala orthohantavirus in areas close to fires due to the mobility and resistance of the rodent in the affected habitats; however, the risk was found to be even higher in non-burned forests [
39].
In the Maule region, Chile, since 2017, there have been a series of large forest fires that have mainly affected coastal areas or the central valley, with fewer reported cases of hantavirus infection. However, in the summer of 2020, a forest fire occurred in the Agua Fría sector in the municipality of Molina (Andes foothills sector), which affected about 13800 hectares, an event that could affect the displacement of rodents from their habitat in non-populated rural areas due to the possible absence of food to populated places, increasing the interaction between rodents and humans. However, due to the COVID-19 pandemic, people’s mobility has been reduced by the prevention measures imposed by the Chilean government (mainly quarantines and sanitary cordons), so there was no increase in HI cases [
32,
41,
42].
On the other hand, the change in land use from native forests to agricultural and forest lands as well as high temperatures in humid climates are identified as relevant factors in the interaction between agricultural workers and the rodent and, therefore, in the spread of hantavirus [
43]. A study in the Atlantic forest [
44] indicates that forest restoration could reduce the possibility of HI transmission by 45%. In [
45], they compared the exclusion of terrestrial mammalian predators and the degradation of the Interior Atlantic forest, showing that seasonality and landscape composition play a fundamental role in the prevalence of rodent reservoirs; in contrast, the exclusion of predators had little influence on the rodent population.
A study conducted in Chile [
46], during 19 years of sampling eleven rodent species, evaluated how ecology and geography influence host and viral dynamics in areas associated with HI cases in that country, finding that the main ANDV reservoir is
O. longicaudatus with an intraspecific seroprevalence of 6.5%. They also point out the need for research on rodents’ social and behavioral interactions, highlighting the integration of ecological understanding of the host and pathogens, spatial and temporal surveillance, epidemiology, and public health agencies, as fundamental to understanding the transmission of the virus to humans. Another study conducted in Patagonia, Argentina [
47] indicates that the high relative abundance of
O. longicaudatus in an unstable community associated with peridomestic environments favors intraspecific contact, leading to a higher probability of virus transmission. Thus, our study integrates, in the proposed mathematical model, ecology (rodent habitat), epidemiology (virus transmission), and territorial dynamics (mobility between urban, rural populated, and non-populated sectors) to contribute from this discipline to the description of virus transmission to humans.
A Bayesian analysis carried out with the expansion of sugar cane and the changes of temperature in Sao Paulo and the risk of hantavirus infection, using historical databases between 2000 and 2010 [
48], demonstrated that the presence of hantavirus cardiopulmonary syndrome was strengthened by the combination of the effects of climate change associated with the increase in temperatures and the transformation of the rodent’s natural habitat in sugar cane cultivations, evidencing similar conditions reported by another study carried out in China [
49] where high temperatures and oscillations of precipitation effected an increase in some types of vegetation that develop in humid areas, influencing the reproduction of the rodent and the mechanism of virus transmission. This, added to the mobility of workers in the rodent territory transformed into an agricultural or forestry crop, increases the risk of spreading hantavirus and the possibility of developing HCPS.
The long-tailed mouse lives less than 2000 m above sea level. In the Maule region, the vegetation and the climate in the Foothills of the Andes mountain range, where the sclerophyllous forest develops with humid zones and high temperatures in the summer, is a suitable habitat for the rodent. Moreover, the population of rodents increases in years that (due to the effect of the Niño current with southern oscillation) there is high pluviometry, which generates an abundance of vegetation and food [
48,
49]. In these years, population abundance promotes the competition for the territory and the displacement to populated sites. So, the displacement to places where humans live to seek sustenance increases, as does HI risk.
According to the analyzed results, the mobility of people going to non-populated sectors has increased, mainly to the realization of camping and excursions in places not predestined for this type of activity, which could increase the interaction between rodents and humans [
50].
Human activities affect natural systems, and global environmental changes put people’s health at risk [
51]. In addition, the mobility of people affects not only acute diseases but also chronic ones [
52]. On the other hand, it is known that if there is an increase in infected rodents, the possibility of infected humans increases [
53,
54], which can be attributed to the invasion of humans into the rodent’s habitat. Therefore, people’s mobility and behavior, such as, for example, the absence of self-care behaviors to avoid the spread of hantavirus, impacts the development of zoonotic diseases. From the results of this study, a significant increase in HI cases is exposed after the mobility of people towards sectors with the presence of rodents—transmitters of infection.
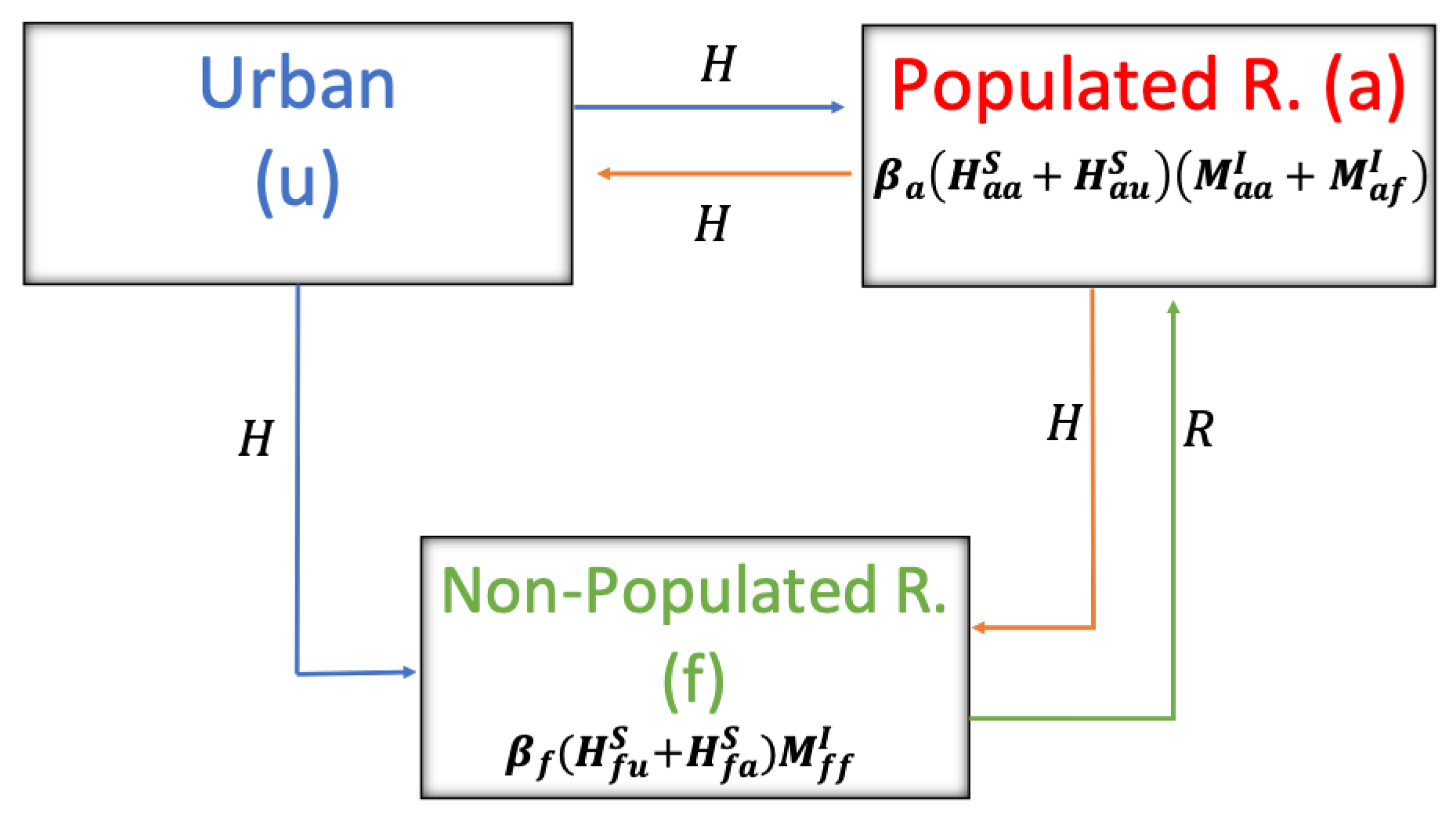
One of the main novelties presented in this work is the proposed HI transmission expression (℘), which explicitly shows how the rates associated with mobility between different sectors affect the contagion rate from rodents to humans.
From the results obtained, we observed the impact of human and rodent mobility on the spread of HI (
Figure 7,
Figure 8 and
Figure 9). The average time an individual stays in the unpopulated rural sector (
Figure 9) is more determinant in HI than the average time a rodent remains in the populated rural sector (
Figure 7) and the rodent mobility flow (
Figure 8). Therefore, there is a greater risk for the populated rural population due to the proximity of unpopulated rural areas and the type of work activities carried out, ratifying the results of other studies and records, which affirm that the most affected population are the inhabitants of the rural sector and agricultural and forestry workers over the occasional visitor. However, due to the increase in outdoor recreational activities, such as camping, excursions, and others, in non-populated areas, the interaction between rodents and people could increase, leading to a significant increase in cases. Therefore, if prevention and control measures are not established, epidemic outbreaks may occur, which can be fatal due to the high virus mortality rate.
The proposed model is a helpful tool that allows decision-makers, especially in the health and housing areas, to understand the evolution of the phenomenon under different scenarios; therefore, it is essential first to establish the epidemiological, social, and geographical characteristics underlying HI to apply the model and generate strategies. However, based on the data we collected from the Chilean territory we modeled, we recommend, in the first instance, active surveillance strategies for rodent control, public education and awareness, personal and household hygiene, safe food storage, prevention in open spaces, improvement of housing infrastructure, timely medical care, research and monitoring, and intersectoral work between the ministries of health, housing, environment, and other relevant entities to comprehensively address the prevention of HI [
55].
In addition to the above, it is essential to avoid constructing houses near nature reserves in risk areas where rodents live [
56,
57]. In this, it is vital to conduct comprehensive risk assessments that determine the presence of rodents, history of hantavirus cases, and other environmental and epidemiological factors that are sufficient causes of the disease; involve the local community in decision-making on urban development and the protection of natural areas [
58] that promote sustainable urban planning which considers the safety of the wild regions and the health of the population [
59]; and strengthen legislation and policies that support the prevention, regulation, and restriction of new housing construction in areas identified as high-risk for HI.
Finally, constant surveillance should also be carried out in the rodent’s territory, especially in natural disasters, forest fires, droughts, and adverse effects of climate change. Thus, by having this information and the effort of preventive measures, new cases of HI in humans would be avoided, and given its high lethality, lives would be saved in sectors where there were no cases of hantavirus.



 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
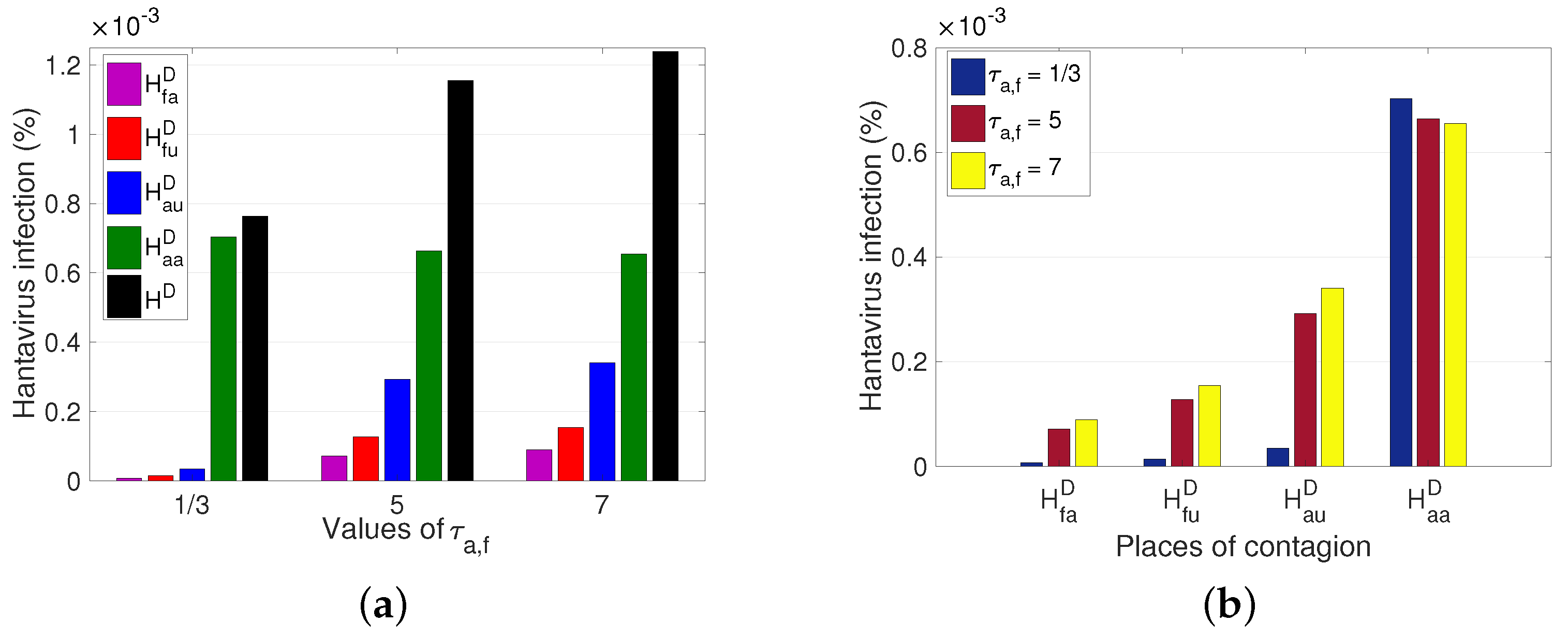{kind=link}