
Canine Schistosomiasis in the West Coast: Heterobilharzia americana in Two Natural Intermediate Hosts Found in the Colorado River, California

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
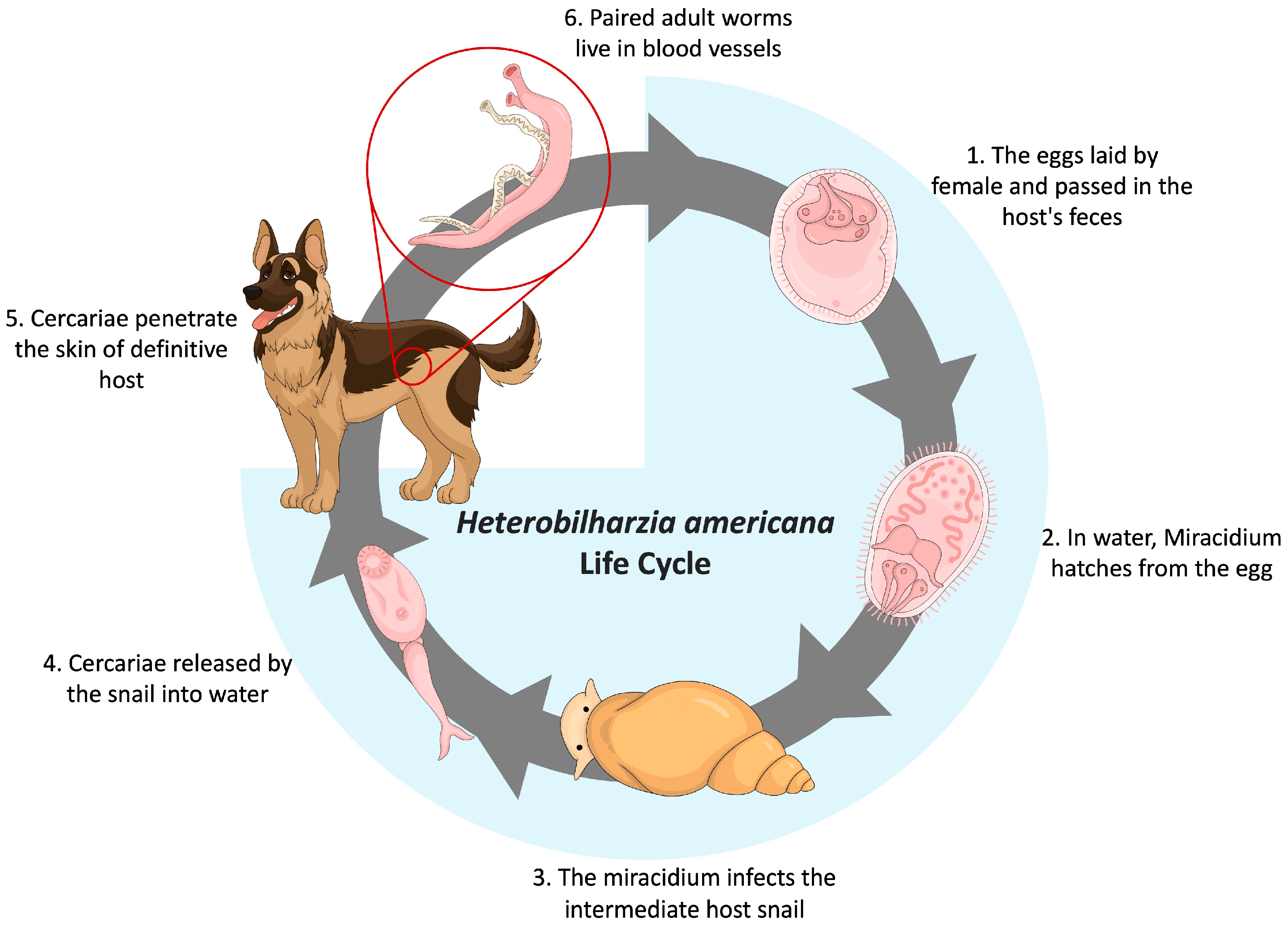
:1. Introduction
2. Materials and Methods
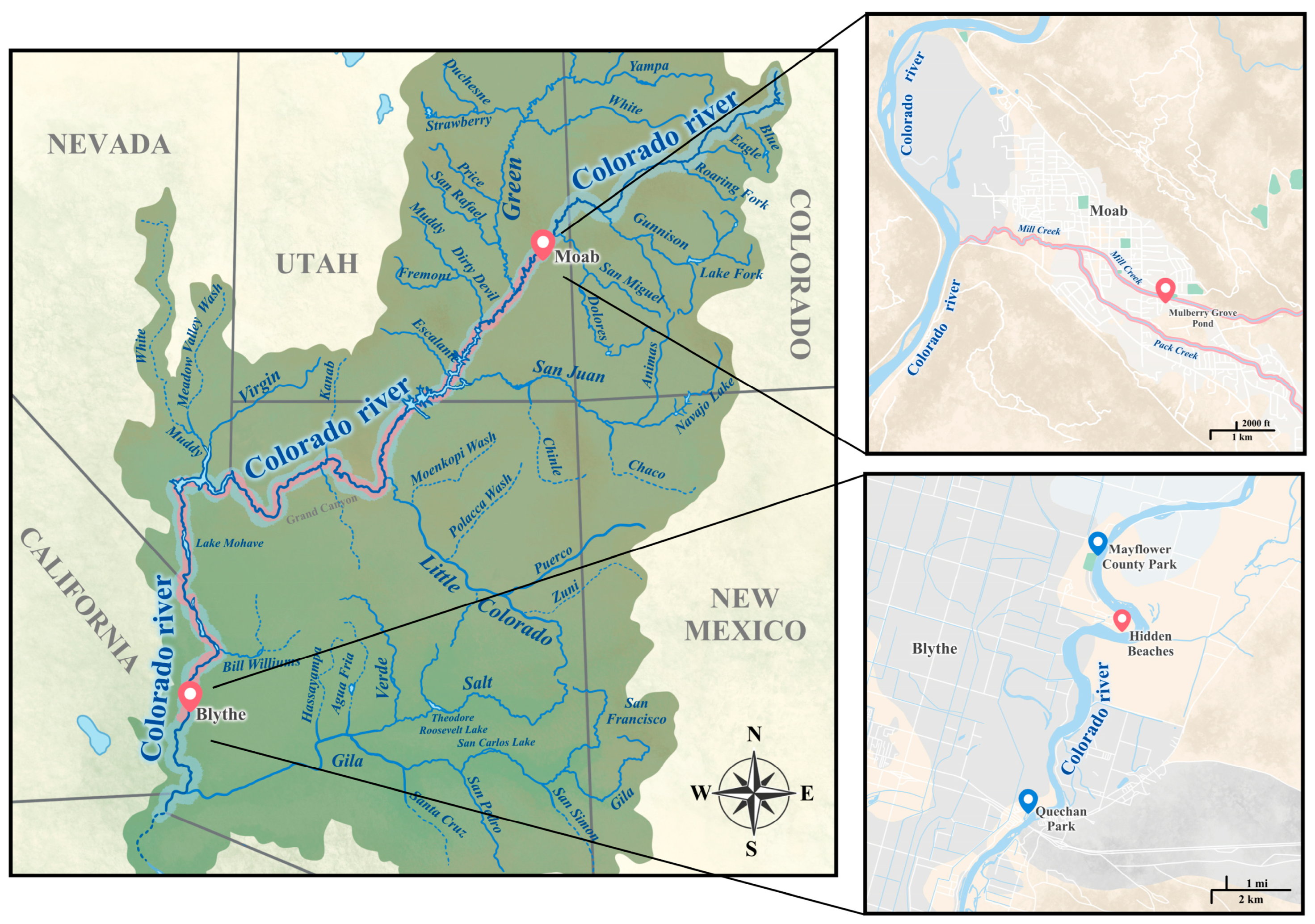
2.1. Collection and Screening of Snails
2.2. DNA Extraction for Molecular Identification of Snails and Cercariae
2.3. DNA Amplification and Sequencing
2.4. Identification and Phylogenetic Analysis
3. Results
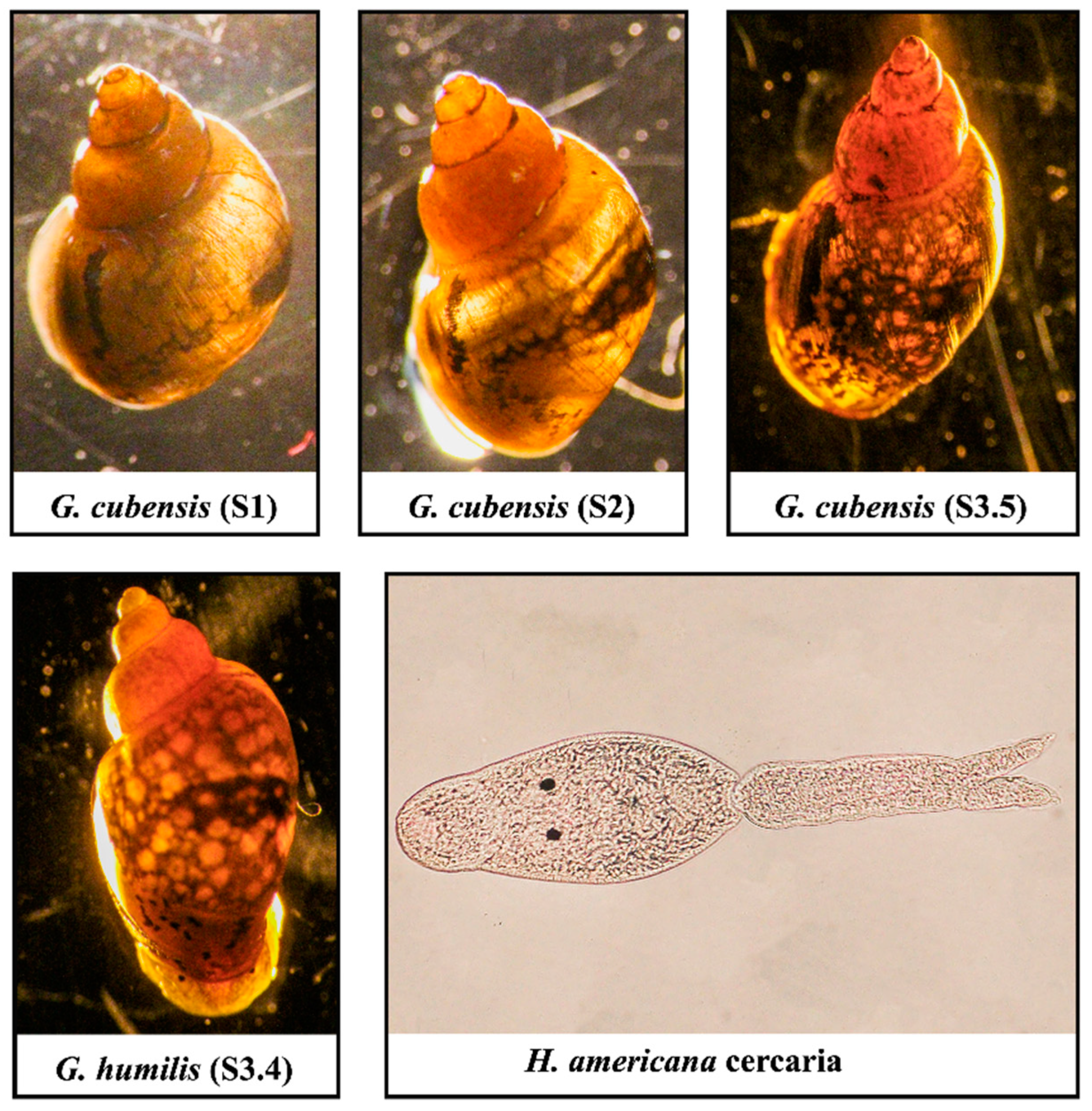
3.1. Survey Results
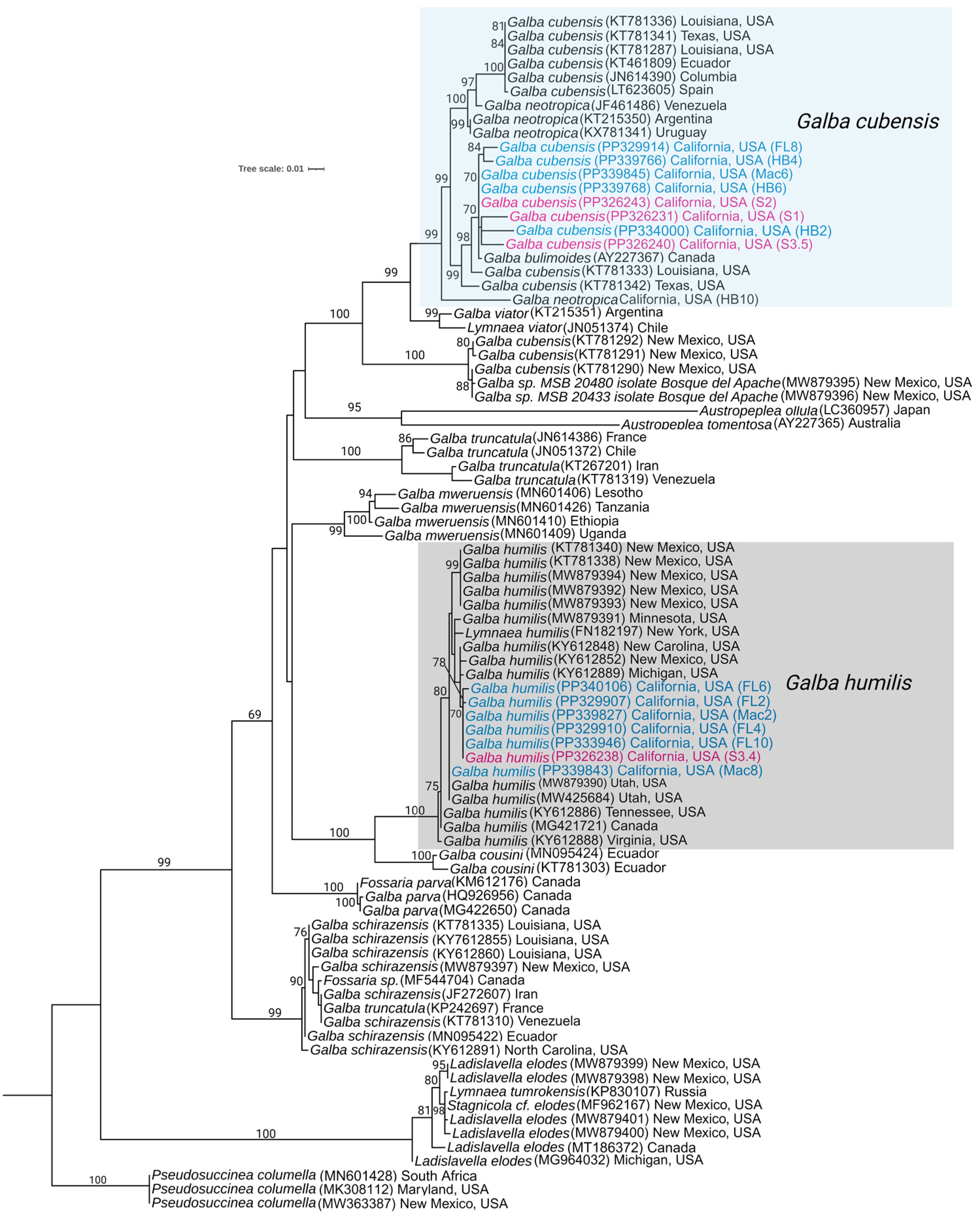
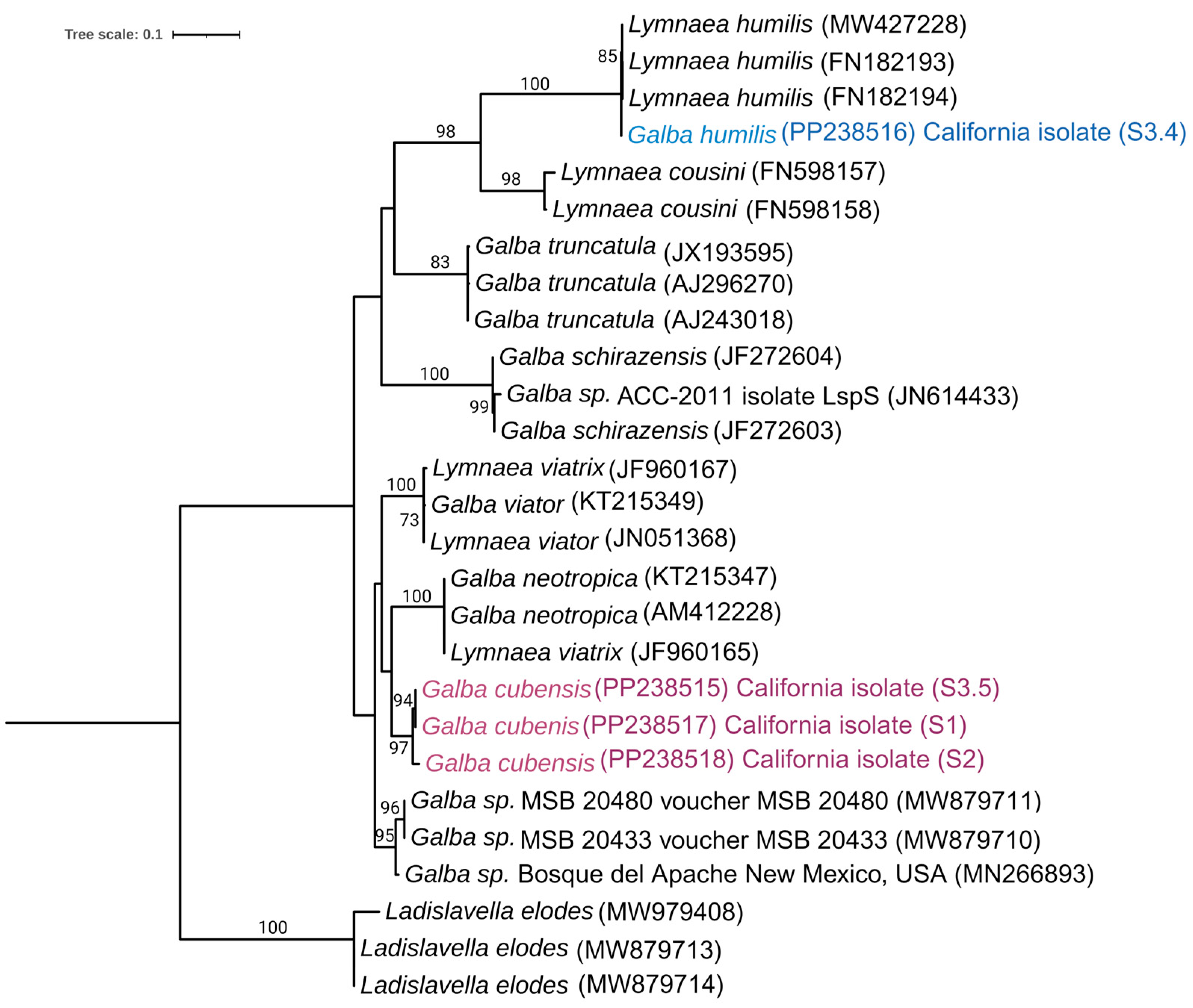
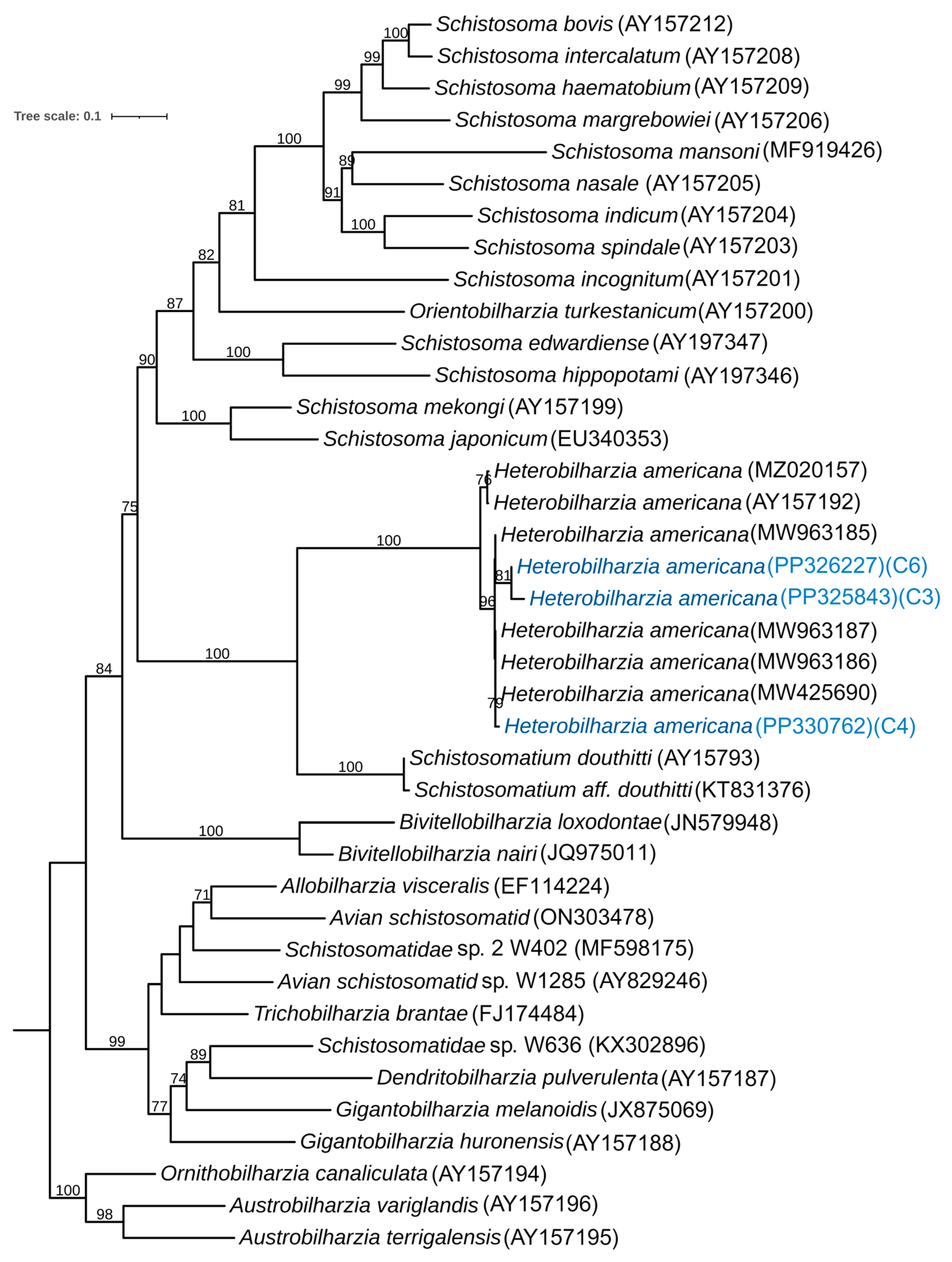
3.2. Phylogenetic Analysis of the Snail
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, T.; Murray, K.A.; Zambrana-Torrelio, C.; Morse, S.S.; Rondinini, C.; Di Marco, M.; Breit, N.; Olival, K.J.; Daszak, P. Global Hotspots and Correlates of Emerging Zoonotic Diseases. Nat. Commun. 2017, 8, 1124. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A Pneumonia Outbreak Associated with a New Coronavirus of Probable Bat Origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Gire, S.K.; Goba, A.; Andersen, K.G.; Sealfon, R.S.G.; Park, D.J.; Kanneh, L.; Jalloh, S.; Momoh, M.; Fullah, M.; Dudas, G.; et al. Genomic Surveillance Elucidates Ebola Virus Origin and Transmission during the 2014 Outbreak. Science 2014, 345, 1369–1372. [Google Scholar] [CrossRef]
- Hosie, M.J.; Jasim, S. The Case for Adopting a Combined Comparative Medicine and One Health Approach to Tackle Emerging Diseases. Vet. Rec. 2020, 187, 24. [Google Scholar] [CrossRef] [PubMed]
- Laing, G.; Vigilato, M.A.N.; Cleaveland, S.; Thumbi, S.M.; Blumberg, L.; Salahuddin, N.; Abdela-Ridder, B.; Harrison, W. One Health for Neglected Tropical Diseases. R. Soc. Trop. Med. Hyg. 2021, 115, 182–184. [Google Scholar] [CrossRef]
- Loker, E.S.; Dolginow, S.Z.; Pape, S.; Topper, C.D.; Alda, P.; Pointier, J.P.; Ebbs, E.T.; Sanchez, M.C.; Verocai, G.G.; DeJong, R.J.; et al. An Outbreak of Canine Schistosomiasis in Utah: Acquisition of a New Snail Host (Galba humilis) by Heterobilharzia americana, a Pathogenic Parasite on the Move. One Health 2021, 13, 100280. [Google Scholar] [CrossRef]
- Brinkerhoff, R.J.; Folsom-O’Keefe, C.M.; Tsao, K.; Diuk-Wasser, M.A. Do Birds Affect Lyme Disease Risk? Range Expansion of the Vector-Borne Pathogen Borrelia burgdorferi. Front. Ecol. Environ. 2011, 9, 103–110. [Google Scholar] [CrossRef]
- Wasel, S.M.; Samuel, W.M.; Crichton, V. Distribution and Ecology of Meningeal Worm, Parelaphostrongylus tenuis (Nematoda), in Northcentral North America. J. Wildl. Dis. 2003, 39, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [PubMed]
- Muehlenbachs, A.; Bhatnagar, J.; Agudelo, C.A.; Hidron, A.; Eberhard, M.L.; Mathison, B.A.; Frace, M.A.; Ito, A.; Metcalfe, M.G.; Rollin, D.C.; et al. Malignant Transformation of Hymenolepis nana in a Human Host. N. Eng. J. Med. 2015, 373, 1845–1852. [Google Scholar] [CrossRef]
- Hossain, M.E.; Kennedy, K.J.; Wilson, H.L.; Spratt, D.; Koehler, A.; Gasser, R.B.; Šlapeta, J.; Hawkins, C.A.; Bandi, H.P.; Senanayake, S.N. Human Neural Larva Migrans Caused by Ophidascaris robertsi Ascarid. Emerg. Infect. Dis. 2023, 29, 1900. [Google Scholar] [CrossRef]
- Malek, E.A. Experimental Infection of Several Lymnaeid Snails with Heterobilharzia americana. J. Parasitol. 1967, 53, 700–702. [Google Scholar] [CrossRef]
- Rodriguez, J.Y.; Lewis, B.C.; Snowden, K.F. Distribution and Characterization of Heterobilharzia americana in Dogs in Texas. Vet. Parasitol. 2014, 203, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.Y.; Camp, J.W.; Lenz, S.D.; Kazacos, K.R.; Snowden, K.F. Identification of Heterobilharzia americana Infection in a Dog Residing in Indiana with No History of Travel. J. Am. Vet. Med. Assoc. 2016, 248, 827–830. [Google Scholar] [CrossRef]
- Johnson, E. Canine Schistosomiasis in North America: An Underdiagnosed Disease. Compen. Cont. Edu. Veter. 2010, E1–E4. [Google Scholar]
- Duncan, K.T.; Koons, N.R.; Litherland, M.A.; Little, S.E.; Nagamori, Y. Prevalence of Intestinal Parasites in Fecal Samples and Estimation of Parasite Contamination from Dog Parks in Central Oklahoma. Vet. Parasitol. Reg. Stud. Rep. 2020, 19, 100362. [Google Scholar] [CrossRef]
- McKown, R.D.; Veatch, J.K.; Fox, L.B. New Locality Record for Heterobilharzia americana. J. Wildl. Dis. 1991, 27, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.F. Life History of Heterobilharzia americana Price 1929, a Schistosome of the Raccoon and Other Mammals in Southeastern United States. J. Parasitol. 1962, 48, 728–739. [Google Scholar] [CrossRef]
- Nation, C.S.; Da’dara, A.A.; Marchant, J.K.; Skelly, P.J. Schistosome Migration in the Definitive Host. PLoS Negl. Trop. Dis. 2020, 14, e0007951. [Google Scholar] [CrossRef] [PubMed]
- Alda, P.; Lounnas, M.; Vázquez, A.A.; Ayaqui, R.; Calvopiña, M.; Celi-Erazo, M.; Dillon, R.T.; González Ramírez, L.C.; Loker, E.S.; Muzzio-Aroca, J.; et al. Systematics and Geographical Distribution of Galba Species, a Group of Cryptic and Worldwide Freshwater Snails. Mol. Phylogenet. Evol. 2021, 157, 107035. [Google Scholar] [CrossRef]
- Laidemitt, M.R.; Loker, E.S. Vector Compatibility of New Mexico Galba Species with the Canine Schistosome Heterobilharzia americana, Including the First Report of Galba schirazensis as a Compatible Host. J. Parasitol. 2023, 109, 633–637. [Google Scholar] [CrossRef]
- Malek, E.A.; Armstrong, J.C. Infection with Heterobilharzia americana in Primates. Am. J. Trop. Med. Hyg. 1967, 16, 708–714. [Google Scholar] [CrossRef]
- Madsen, H.; Frandsen, F. The Spread of Freshwater Snails Including Those of Medical and Veterinary Importance. Acta Trop. 1989, 46, 139–146. [Google Scholar] [CrossRef]
- Johnson, P.D.; Bogan, A.E.; Brown, K.M.; Burkhead, N.M.; Cordeiro, J.R.; Garner, J.T.; Hartfield, P.D.; Lepitzki, D.A.W.; Mackie, G.L.; Pip, E.; et al. Conservation Status of Freshwater Gastropods of Canada and the United States. Fisheries 2013, 38, 247–282. [Google Scholar] [CrossRef]
- Lounnas, M.; Vázquez, A.A.; Alda, P.; Sartori, K.; Pointier, J.P.; David, P.; Hurtrez-Boussès, S. Isolation, Characterization and Population-Genetic Analysis of Microsatellite Loci in the Freshwater Snail Galba cubensis (Lymnaeidae). J. Moll. Stud. 2017, 83, 63–68. [Google Scholar] [CrossRef]
- Vázquez, A.A.; de Vargas, M.; Alba, A.; Sánchez, J.; Alda, P.; Sabourin, E.; Vittecoq, M.; Alarcón-Elbal, P.M.; Pointier, J.P.; Hurtrez-Boussès, S. Reviewing Fasciola hepatica Transmission in the West Indies and Novel Perceptions from Experimental Infections of Sympatric vs. Allopatric Snail/Fluke Combinations. Vet. Parasitol. 2019, 275, 108955. [Google Scholar] [CrossRef] [PubMed]
- Saito, T. First Record of the Non-Indigenous Freshwater Snail Galba humilis (Say, 1822) (Mollusca: Hygrophila: Lymnaeidae) in Japan. Bioinvasions Rec. 2022, 11, 428–439. [Google Scholar] [CrossRef]
- Thompson, F.G. The Freshwater Snails of Florida. J. Moll. Biol. 1999, 10, 1–84. [Google Scholar]
- Jokinen, E.H. The Freshwater Snails (Mollusca: Gastropoda) of New York State; Biol. Survey; University of the State of New York, State Education Department, New York State Museum: Albany, NY, USA, 1992. [Google Scholar]
- Clarke, A.H. The Freshwater Molluscs of Canada; Clarke, A.H., Ed.; National Museum of Natural Sciences, National Museums of Canada: Ottawa, ON, Canada, 1981. [Google Scholar]
- Fabrick, C.; Bugbee, A.; Fosgate, G. Clinical Features and Outcome of Heterobilharzia americana Infection in Dogs. J. Vet. Intern. Med. 2010, 24, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Flowers, J.; Hammerberg, B.; Wood, S.; Malarkey, D.; van Dam, G.J.; Levy, M.; McLawhorn, L. Heterobilharzia americana Infection in a Dog. Am. Veter. Med. Assoc. 2002, 220, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Fradkin, J.M.; Braniecki, A.M.; Craig, T.M.; Ramiro-Ibanez, F.; Rogers, K.S.; Zoran, D.L. Elevated Parathyroid Hormone-Related Protein and Hypercalcemia in Two Dogs with Schistosomiasis. J. Am. Anim. Hosp. Assoc. 2001, 37, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Rohrer, C.R.; Phillips, L.A.; Ford, S.L.; Ginn, P.E. Hypercalcemia in a Dog: A Challenging Case. J. Am. Anim. Hosp. Assoc. 2000, 36, 20–25. [Google Scholar] [CrossRef]
- Bartsch, R.C.; Ward, B.C. Visceral Lesions in Raccoons Naturally Infected with Heterobilharzia americana. Vet. Pathol. 1976, 13, 241–249. [Google Scholar] [CrossRef]
- Corapi, W.V.; Snowden, K.F.; Rodrigues, A.; Porter, B.F.; Buote, M.A.; Birch, S.M.; Jackson, N.D.; Eden, K.B.; Whitley, D.B.; Mansell, J.; et al. Natural Heterobilharzia americana Infection in Horses in Texas. Vet. Pathol. 2012, 49, 552–556. [Google Scholar] [CrossRef]
- Veterinary Public Health; Los Angeles County Department of Public Health. Animal Health Advisory: Heterobilharzia americana (Canine Schistosomiasis) Confirmed in 11 Dogs in Three Southern California Counties (LA, Orange, and Riverside). J. Parasitol. 2023, 109, 633–637. [Google Scholar]
- Corapi, W.V.; Eden, K.B.; Edwards, J.F.; Snowden, K.F. Heterobilharzia americana Infection and Congestive Heart Failure in a Llama (Lama glama). Vet. Pathol. 2015, 52, 562–565. [Google Scholar] [CrossRef]
- Almeyda-Artigas, R.J.; Bargues, M.D.; MasComa, S. ITS-2 RDNA Sequencing of Gnathostoma Species (Nematoda) and Elucidation of the Species Causing Human Gnathostomiasis in the Americas. J. Parasitol. 2000, 86, 537–544. [Google Scholar] [CrossRef]
- Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Bio Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Lockyer, A.E.; Olson, P.D.; Østergaard, P.; Rollinson, D.; Johnston, D.A.; Attwood, S.W.; Southgate, V.R.; Horak, P.; Snyder, S.D.; Le, T.H.; et al. The Phylogeny of the Schistosomatidae Based on Three Genes with Emphasis on the Interrelationships of Schistosoma Weinland, 1858. Parasitology 2003, 126, 203–224. [Google Scholar] [CrossRef]
- Brant, S.V.; Loker, E.S. Molecular Systematics of the Avian Schistosome Genus Trichobilharzia (Trematoda: Schistosomatidae) in North America. J. Parasitol. 2009, 95, 941–963. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.A.T.; DeJong, R.J.; Kazibwe, F.; Mkoji, G.M.; Loker, E.S. A Newly-Identified Lineage of Schistosoma. Int. J. Parasitol. 2003, 33, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Asimenos, G.; Toh, H. Multiple Alignment of DNA Sequences with MAFFT. Met. Mol. Biol. 2009, 537, 39–64. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Meunier, C.; Tirard, C.; Hurtrez-Boussès, S.; Durand, P.; Bargues, M.D.; Mas-Coma, S.; Pointier, P.; Jourdane, J.; Renaud, F. Lack of Molluscan Host Diversity and the Transmission of an Emerging Parasitic Disease in Bolivia. Mol. Ecol. 2001, 10, 1333–1340. [Google Scholar] [CrossRef]
- Lounnas, M.; Correa, A.C.; Alda, P.; David, P.; Dubois, M.P.; Calvopiña, M.; Caron, Y.; Celi-Erazo, M.; Dung, B.T.; Jarne, P.; et al. Population Structure and Genetic Diversity in the Invasive Freshwater Snail Galba schirazensis (Lymnaeidae). Can. J. Zool. 2018, 96, 425–435. [Google Scholar] [CrossRef]
- Correa, A.C.; Escobar, J.S.; Durand, P.; Renaud, F.; David, P.; Jarne, P.; Pointier, J.P.; Hurtrez-Boussès, S. Bridging Gaps in the Molecular Phylogeny of the Lymnaeidae (Gastropoda: Pulmonata), Vectors of Fascioliasis. BMC Evol. Biol. 2010, 10, 381. [Google Scholar] [CrossRef]
- Correa, A.C.; Escobar, J.S.; Noya, O.; Velásquez, L.E.; González-Ramírez, C.; Hurtrez-Boussès, S.; Pointier, J.P. Morphological and Molecular Characterization of Neotropic lymnaeidae (Gastropoda: Lymnaeoidea), Vectors of Fasciolosis. Infect. Genet. Evol. 2011, 11, 1978–1988. [Google Scholar] [CrossRef]
- Bargues, M.D.; Artigas, P.; Khoubbane, M.; Flores, R.; Glöer, P.; Rojas-García, R.; Ashrafi, K.; Falkner, G.; Mas-Coma, S. Lymnaea schirazensis, an Overlooked Snail Distorting Fascioliasis Data: Genotype, Phenotype, Ecology, Worldwide Spread, Susceptibility, Applicability. PLoS ONE 2011, 6, e24567. [Google Scholar] [CrossRef]
- Meunier, C.; Hurtrez-Boussès, S.; Jabbour-Zahab, R.; Durand, P.; Rondelaud, D.; Renaud, F. Field and Experimental Evidence of Preferential Selfing in the Freshwater Mollusc Lymnaea truncatula (Gastropoda, Pulmonata). Heredity 2004, 92, 316–322. [Google Scholar] [CrossRef]
- Alda, P.; Lounnas, M.; Vázquez, A.A.; Ayaqui, R.; Calvopiña, M.; Celi-Erazo, M.; Dillon, R.T.; Jarne, P.; Loker, E.S.; Muñiz Pareja, F.C.; et al. A New Multiplex PCR Assay to Distinguish among Three Cryptic Galba Species, Intermediate Hosts of Fasciola hepatica. Vet. Parasitol. 2018, 251, 101–105. [Google Scholar] [CrossRef]
- Bogan, M.A.; Finley, R.B.; Fitzgerald, J.P.; Meaney, C.A.; Armstrong, D.M. Mammals of Colorado. J. Mammal. 1996, 77, 901. [Google Scholar] [CrossRef]
- Kelley, S.W. Heterobilharziasis (Trematoda: Schistosomatidae) in Raccoons (Procyon lotor) of North-Central Texas. Tex. J. Sci. 2009, 62, 41–48. [Google Scholar]
- Savioli, L.; Renganathan, E.; Montresor, A.; Davis, A.; Behbehani, K. Control of Schistosomiasis-A Global Picture. Parasitol. Today 1997, 13, 444–448. [Google Scholar] [CrossRef]
- De Gentile, L.; Picot, H.; Bourdeau, P.; Bardet, R.; Kerjan, A.; Piriou, M.; Le Guennic, A.; Bayssade-Dufour, C.; Chabasse, D.; Mott, K.E. Cercarial Dermatitis in Europe: A New Public Health Problem? Bull. World Health Organ. 1996, 74, 159. [Google Scholar]
- Platt, T.R.; Brooks, D.R. Evolution of the Schistosomes (Digenea: Schistosomatoidea): The Origin of Dioecy and Colonization of the Venous System. J. Parasitol. 1997, 83, 1035–1044. [Google Scholar] [CrossRef]
- Snyder, S.D.; Loker, E.S. Evolutionary Relationships Among the Schistosomatidae (Platyhelminthes: Digenea) and an Asian Origin for Schistosoma. J. Parasitol. 2000, 86, 283–288. [Google Scholar] [CrossRef]
- Malek, E.A. Further Studies on Mammalian Susceptibility to Experimental Infection with Heterobilharzia americana. J. Parasitol. 1970, 56, 64–66. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baniya, A.; Goldy, C.J.; Ardpairin, J.; Achi, P.; Chang, Y.W.; Adrianza, R.C.; Vitta, A.; Dillman, A.R. Canine Schistosomiasis in the West Coast: Heterobilharzia americana in Two Natural Intermediate Hosts Found in the Colorado River, California. Pathogens 2024, 13, 245. https://doi.org/10.3390/pathogens13030245
Baniya A, Goldy CJ, Ardpairin J, Achi P, Chang YW, Adrianza RC, Vitta A, Dillman AR. Canine Schistosomiasis in the West Coast: Heterobilharzia americana in Two Natural Intermediate Hosts Found in the Colorado River, California. Pathogens. 2024; 13(3):245. https://doi.org/10.3390/pathogens13030245
Chicago/Turabian StyleBaniya, Anil, Connor J. Goldy, Jiranun Ardpairin, Perla Achi, Yu Wei Chang, Rose C. Adrianza, Apichat Vitta, and Adler R. Dillman. 2024. "Canine Schistosomiasis in the West Coast: Heterobilharzia americana in Two Natural Intermediate Hosts Found in the Colorado River, California" Pathogens 13, no. 3: 245. https://doi.org/10.3390/pathogens13030245
APA StyleBaniya, A., Goldy, C. J., Ardpairin, J., Achi, P., Chang, Y. W., Adrianza, R. C., Vitta, A., & Dillman, A. R. (2024). Canine Schistosomiasis in the West Coast: Heterobilharzia americana in Two Natural Intermediate Hosts Found in the Colorado River, California. Pathogens, 13(3), 245. https://doi.org/10.3390/pathogens13030245