Performance Indices in Wheat Chlorophyll a Fluorescence and Protein Quality Influenced by FHB

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
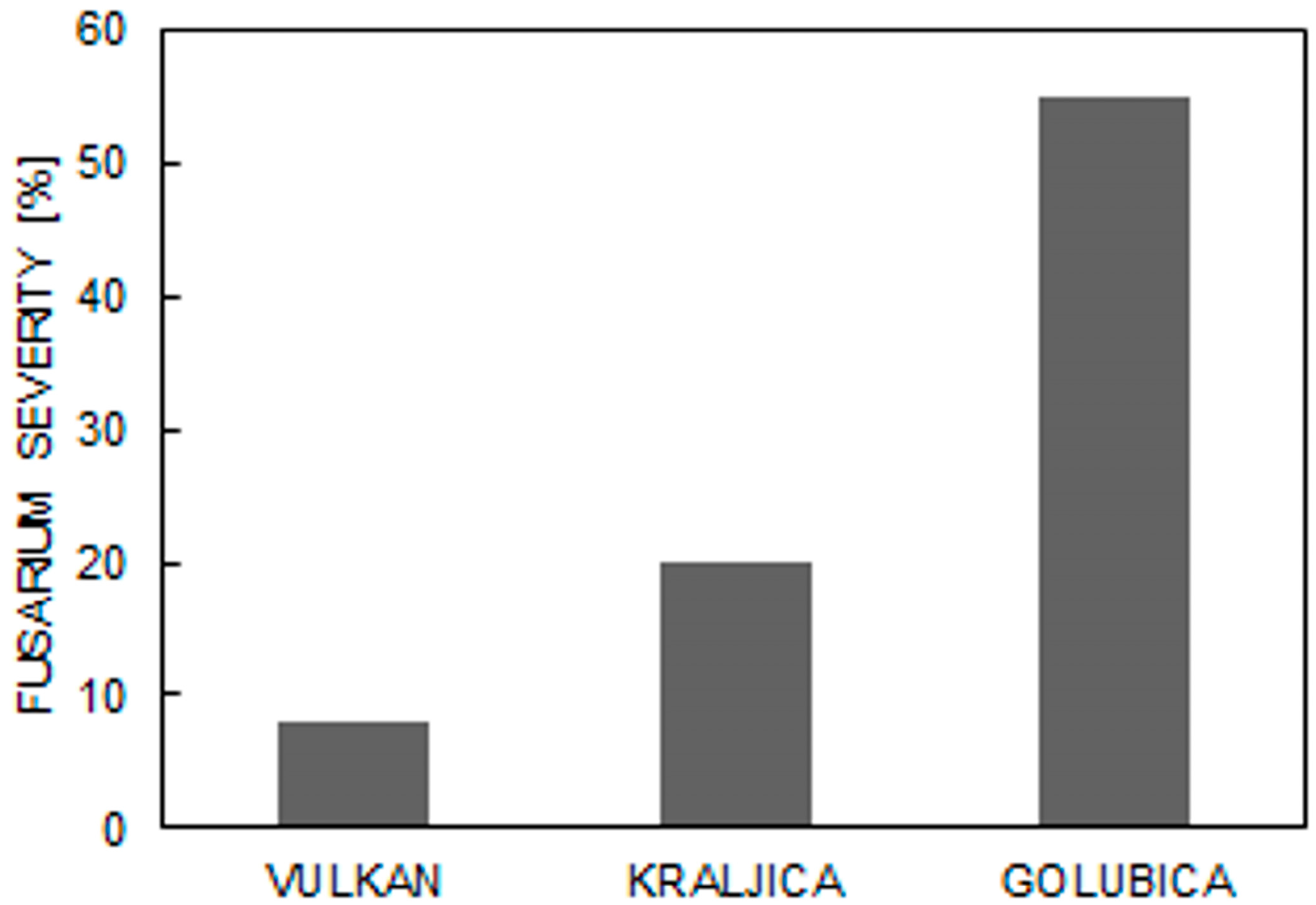
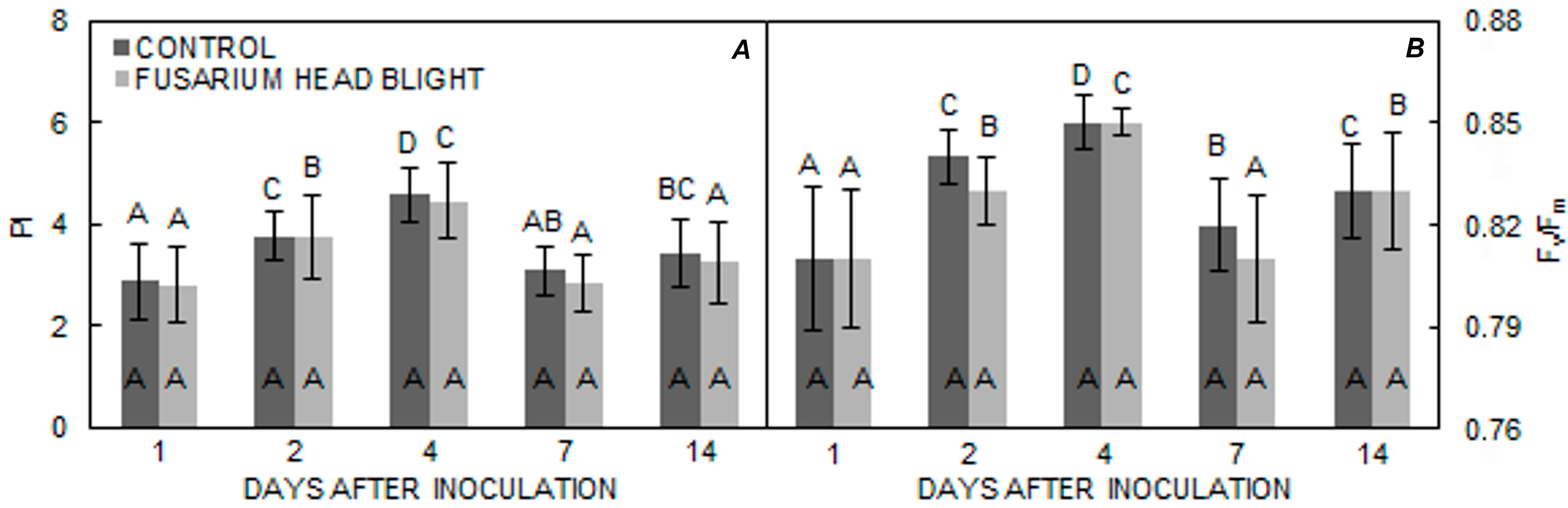
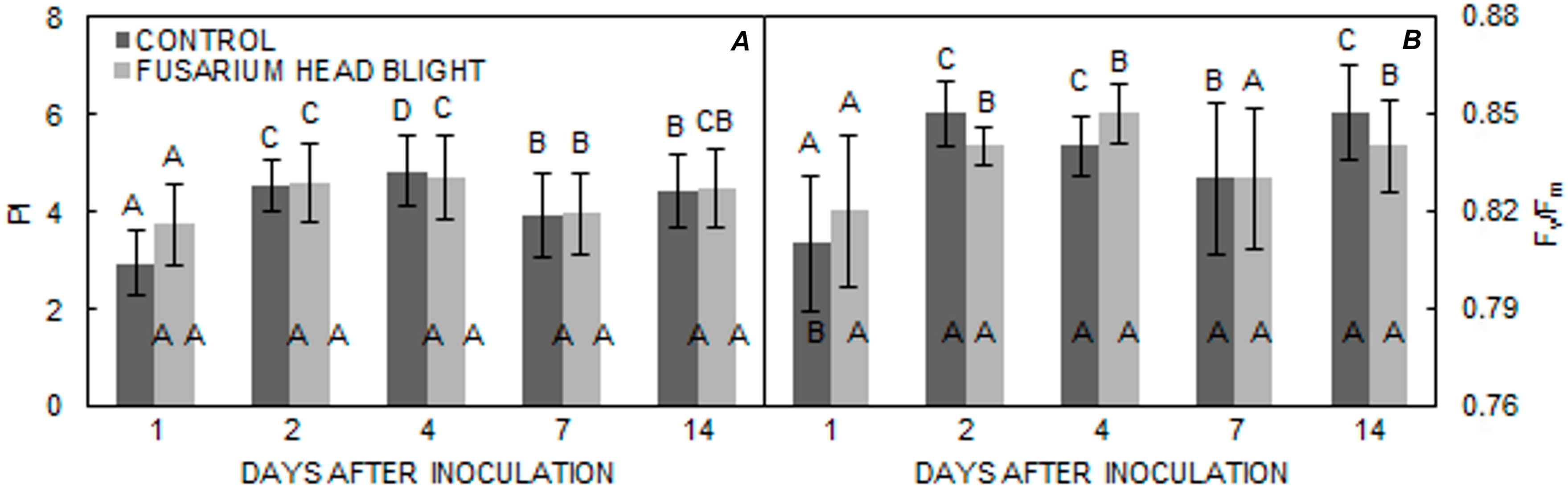
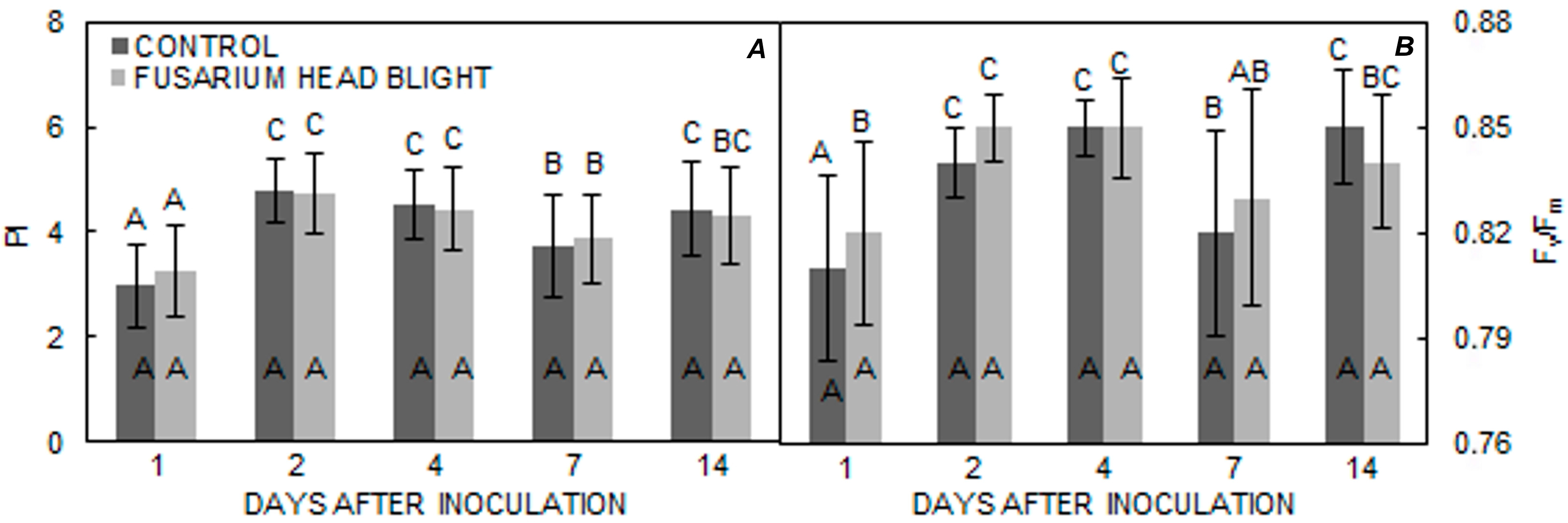
2. Results
3. Discussion
4. Materials and Methods
4.1. Inoculum Production
4.2. Field Trial
4.3. Chlorophyll a Fluorescence
4.4. Proteins Characterization
4.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Bai, G.H.; Guo, P.; Kolb, F.L. Genetics Relationships among Head Blight Resistance Cultivars of Wheat Assessed on the Basis of Molecular Markers. Crop Sci. 2003, 43, 498–507. [Google Scholar] [CrossRef]
- Bai, G.H.; Shaner, G.E. Scab of wheat: Prospects for control. Plant Dis. 1994, 78, 760–766. [Google Scholar] [CrossRef]
- World Health Organization. Deoxynivalenol. In WHO Food Additives Series 47; World Health Organization: Geneva, Switzerland, 2001; Volume 74, pp. 419–556. [Google Scholar]
- Blandino, M.; Pilati, A.; Reyneri, A. Effect of foliar treatments to durum wheat on flag leaf senescence, grain yield, quality and deoxynivalenol contamination in North Italy. Field Crops Res. 2009, 114, 214–222. [Google Scholar] [CrossRef]
- Gil, D.H.; Bonfil, D.J.; Svoray, T. Multi scale analysis of the factors influencing wheat quality as determined by Gluten Index. Field Crops Res. 2011, 123, 1–9. [Google Scholar] [CrossRef]
- Czaban, J.; Wróblewska, B.; Sułek, A.; Mikos, M.; Boguszewska, E.; Podolska, G.; Nieróbca, A. Colonisation of winter wheat grain by Fusarium. spp. and mycotoxin content as dependent on a wheat variety, crop rotation, a crop management system and weather conditions. Food Addit. Contam. 2015, 32, 874–910. [Google Scholar] [CrossRef] [PubMed]
- Debona, D.; Rodrigues, F.A.; Rios, J.A.; Martins, S.C.V.; Pereira, L.F.; DaMatta, F.M. Limitations to Photosynthesis in Leaves of Wheat Plants Infected by Pyricularia oryzae. Phytopathology 2014, 104, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Živčák, M.; Brestič, M.; Olšovská, K.; Slamka, P. Performance index as a sensitive indicator of water stressin Triticum. aestivum L. Plant Soil Environ. 2008, 54, 133–139. [Google Scholar]
- Yang, S.M.; Li, X.; Chen, W.Q.; Liu, T.G.; Zhong, S.F.; Ma, L.X.; Zhang, M.; Zhang, H.Y.; Yu, D.L.; Luo, P.G. Wheat Resistance to Fusarium Head Blight is Associated With Changes in Photosynthetic Parameters. Plant Dis. 2016, 100, 847–852. [Google Scholar] [CrossRef]
- Wagner, A.; Michalek, V.; Jamiolkowska, A. Chlorophyll fluorescence measurements as indicators of fusariosis severity in tomato plants. Agron. Res. 2006, 4, 461–464. [Google Scholar]
- Sylvester-Bradley, R.; Scott, R.K.; Wright, C.E. Physiology in the production and improvement of cereals. In Home-Grown Cereals Authority Research Review; HGCA: London, UK, 1990; Volume 18. [Google Scholar]
- Gregersen, P.L.; Holm, P.B.; Krupinska, K. Leaf senescence and nutrient remobilisation in barley and wheat. Plant Biol. 2008, 10, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Luo, P.; Ren, Z.; Zhang, H.Y. Controlling fusarium head blight of wheat (Triticum. aestivum L.) with genetic. Adv. Biosci. Biotechnol. 2011, 2, 263–270. [Google Scholar] [CrossRef]
- Eggert, K.; Wieser, H.; Pawelzik, E. The influence of Fusarium infection and growing location on the quantitative protein composition of (part I) emmer (Triticum. dicoccum). Eur. Food Res. Technol. 2010, 230, 837–847. [Google Scholar] [CrossRef]
- Eggert, K.; Rawel, H.M.; Pawelzik, E. In vitro degradation of wheat gluten fractions by Fusarium graminearum proteases. Eur. Food Res. Technol. 2011, 233, 697–705. [Google Scholar] [CrossRef]
- Horvat, D.; Španić, V.; Dvojković, K.; Šimić, G.; Magdić, D.; Nevistić, A. The Influence of Fusarium Infection on Wheat (Triticum. aestivum L.) Proteins Distribution and Baking Quality. Cereal Res. Commun. 2015, 43, 61–71. [Google Scholar] [CrossRef]
- Ali, M.; Saddiqui, M.F.; Ullah, I.; Bakht, J. Analysis of HMW-GS of historical set of Pakistani bread wheat varieties using SDS-PAGE. Afr. J. Biotechnol. 2010, 9, 4286–4289. [Google Scholar]
- Mastilović, J.; Horvat, D.; Živančev, D.; Torbica, T.; Kevrešan, Ž.; Đukić, N.; Magdić, D.; Šimić, G. Analysis of interrelations between wheat protein fractions composition and its technological quality by combined multivariate and univariate statistics. Hem. Ind. 2014, 68, 321–329. [Google Scholar] [CrossRef]
- Murata, Y. Dependence of the potential productivity and efficiency in solar energy utilization on leaf photosynthetic capacity in crop species. Jpn. J. Crop Sci. 1981, 50, 223–232. [Google Scholar] [CrossRef]
- Evans, L.T.; Rawson, H.M. Photosynthesis and respiration by the flag leaf and components of the ear during grain development in wheat. Aust. J. Biol. Sci. 1970, 23, 245–254. [Google Scholar] [CrossRef]
- Abbad, H.; El Jaafari, S.; Bort, J.; Araus, J.L. Comparison of flag leaf and ear photosynthesis with biomass and grain yield of durum wheat under various water conditions and genotypes. Agronomie 2004, 24, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Kibite, S.; Evans, L.E. Causes of negative correlations between grain yield and grain protein concentration in common wheat. Euphytica 1984, 33, 801–810. [Google Scholar] [CrossRef]
- Drezner, G.; Dvojkovic, K.; Horvat, D.; Novoselovic, D.; Lalic, A.; Babic, D.; Kovacevic, J. Grain yield and quality of winter wheat genotypes in different environments. Cereal Res. Commun. 2006, 34, 457–460. [Google Scholar] [CrossRef]
- Papoušková, L.; Capouchová, I.; Kostelanská, M.; Škeříková, A.; Prokinová, E.; Hajšlová, J.; Salava, J.; Faměra, O. Changes in baking quality of winter wheat with different intensity of Fusarium spp. contamination detected by means of new rheological system Mixolab. Czech J. Food Sci. 2011, 29, 420–429. [Google Scholar]
- Sanchez-Bragado, R.; Molero, G.; Reynolds, M.P.; Araus, J.L. Photosynthetic contribution of the ear to grain filling in wheat: A comparison of different methodologies for evaluation. J. Exp. Bot. 2016, 67, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Slafer, G.A.; Savin, R. Post-anthesis green area duration in a semi-dwarf and a standard-height wheat cultivar as affected by sink strength. Aust. J. Agric. 1994, 45, 1337–1346. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A.; Nelson, R.; Long, S.P. Testing the ‘source–sink’ hypothesis of downregulation of photosynthesis in elevated (CO2) in the field with single gene substitutions in Glycine max. Agric. For. Meteorol. 2004, 122, 85–94. [Google Scholar] [CrossRef]
- Hörtensteiner, S.; Feller, U. Nitrogen metabolism and remobilization during senescence. J. Exp. Bot. 2002, 53, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Christen, D.; Schönmann, S.; Jermini, M.; Strasser, R.J.; Dèfago, G. Characterization and early detection of grapevine (Vitis. vinifera) stress responses to esca disease by in situ chlorophyll fluorescence and comparison with drought stress. Environ. Exp. Bot. 2007, 60, 504–514. [Google Scholar] [CrossRef]
- Snijders, C.H.A.; Van Eeuwijk, F.A. Genotype X strain interactions for resistance to Fusarium head blight caused by Fusarium. culmorum in winter wheat. Theor. Appl. Genet. 1991, 81, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, M.; Haim, K.; Lew, H.; Ruckenbauer, P. The effect of nitrogen fertilization on Fusarium head blight development and deoxynivalenol contamination in wheat. Phytopathology 2004, 152, 1–8. [Google Scholar] [CrossRef]
- Lemmens, M.; Buerstmayr, H.; Ruckenabauer, P. Variation in Fusarium head blight susceptibility of international and Austrian wheat breeding material. Die Bodenkultur 1993, 44, 65–78. [Google Scholar]
- Spanic, V.; Drezner, G. Influence of Fusarium artificial infection on agronomic traits. Glasnik Zaštite Bilja 2011, 4, 60–64. [Google Scholar]
- Spanic, V.; Lemmens, M.; Drezner, G. Variability of components of fusarium head blight resistance among wheat genotypes. Cereal Res. Commun. 2013, 41, 420–430. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzac, F.C. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Stirbet, A.; Govindjee. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Wieser, H.; Antes, S.; Selmeier, W. Quantitative determination of gluten protein types in wheat flour by reverse-phase high preformance liquid chromatography. Cereal Chem. 1998, 75, 644–650. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Df | Mean Square | ||||||
|---|---|---|---|---|---|---|---|---|
| TW | TKW | AG | GLI | GLU | HMW | LMW | ||
| Variety | 2 | 102.6 *** | 14.7 ** | 2.810 * | 43.40 *** | 28.87 *** | 18.11 *** | 30.29 *** |
| Treatment | 1 | 13.44 ** | 11.21 * | 0.090ns | 21.12 *** | 18.45 *** | 2.98 *** | 9.11 *** |
| Variety * Treatment | 2 | 73.86 *** | 21.05 * | 6.741 *** | 24.94 *** | 32.41 *** | 2.93 *** | 13.13 *** |
| Error | 6 | 1.17 | 1.39 | 0.364 | 0.3 | 0.32 | 0.09 | 0.3 |
| Source of Variation | Df | Mean Square | |
|---|---|---|---|
| PI | Fv/Fm | ||
| Variety | 2 | 27.096 *** | 0.0048 ** |
| Measurement time | 4 | 44.415 *** | 0.0246 *** |
| Treatment | 1 | 0.052 | 0.0001 |
| Variety × Measurement time | 8 | 2.894 *** | 0.0005 |
| Variety × Treatment | 2 | 0.931 | 0.0005 |
| Measurement time × Treatment | 4 | 0.368 | 0.0006 |
| Variety × Measurement time × Treatment | 8 | 0.157 | 0.0002 |
| Error | 561 | 0.583 | 0.0003 |
| PI | Fv/Fm | |||
|---|---|---|---|---|
| Con | FHB | Con | FHB | |
| Vulkan | 3.5330 b | 3.4145 b | 0.8275 b | 0.8250 b |
| Kraljica | 4.0642 a | 4.2181 a | 0.8331 a | 0.8366 a |
| Golubica | 4.0890 a | 4.1230 a | 0.8344 a | 0.8357 a |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spanic, V.; Viljevac Vuletic, M.; Drezner, G.; Zdunic, Z.; Horvat, D. Performance Indices in Wheat Chlorophyll a Fluorescence and Protein Quality Influenced by FHB. Pathogens 2017, 6, 59. https://doi.org/10.3390/pathogens6040059
Spanic V, Viljevac Vuletic M, Drezner G, Zdunic Z, Horvat D. Performance Indices in Wheat Chlorophyll a Fluorescence and Protein Quality Influenced by FHB. Pathogens. 2017; 6(4):59. https://doi.org/10.3390/pathogens6040059
Chicago/Turabian StyleSpanic, Valentina, Marija Viljevac Vuletic, Georg Drezner, Zvonimir Zdunic, and Daniela Horvat. 2017. "Performance Indices in Wheat Chlorophyll a Fluorescence and Protein Quality Influenced by FHB" Pathogens 6, no. 4: 59. https://doi.org/10.3390/pathogens6040059
APA StyleSpanic, V., Viljevac Vuletic, M., Drezner, G., Zdunic, Z., & Horvat, D. (2017). Performance Indices in Wheat Chlorophyll a Fluorescence and Protein Quality Influenced by FHB. Pathogens, 6(4), 59. https://doi.org/10.3390/pathogens6040059