Immunosuppressive Compounds Affect the Fungal Growth and Viability of Defined Aspergillus Species


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
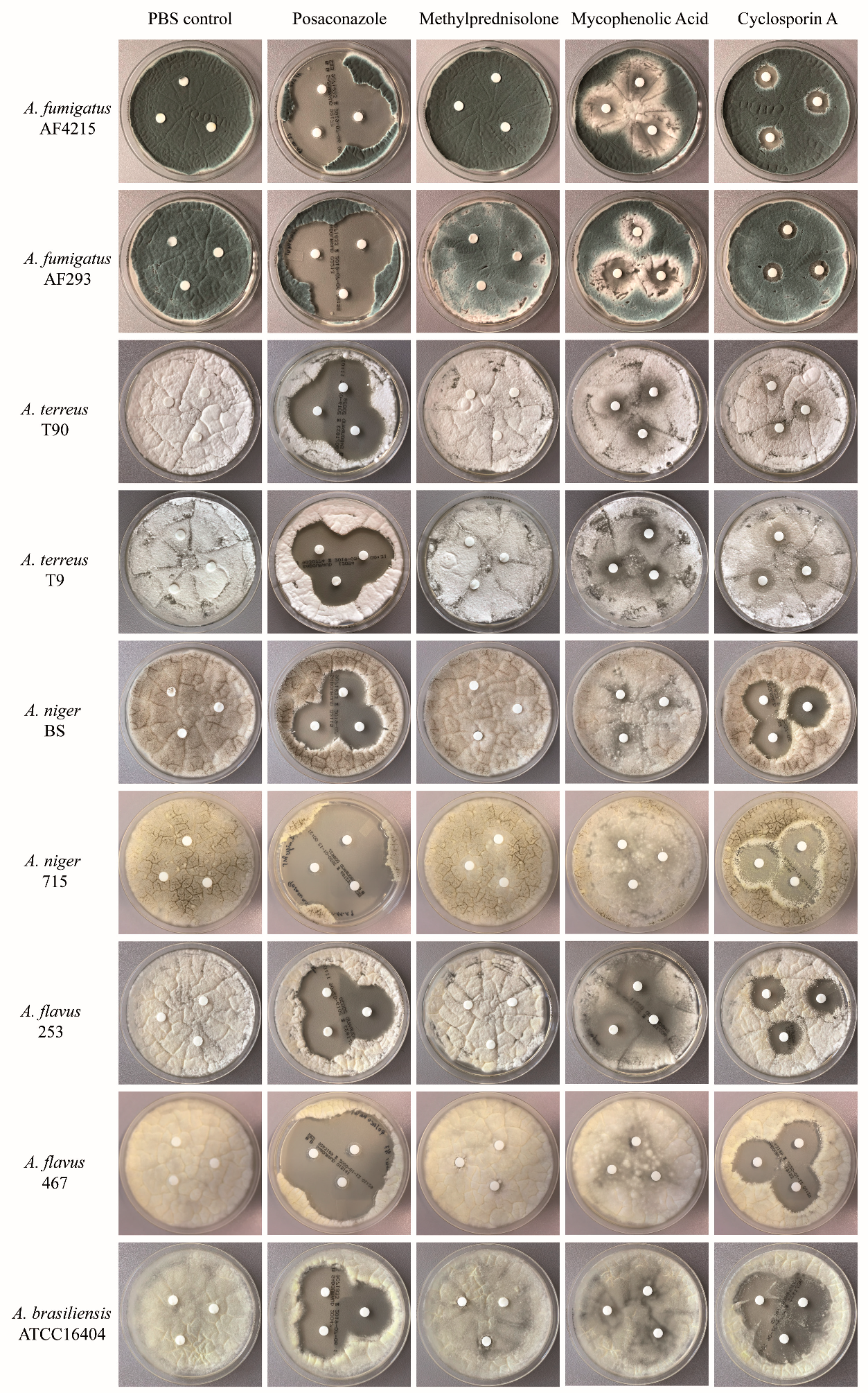
2.1. Immunosuppressive Agents Inhibit the Fungal Growth of Aspergillus Species—Disc Diffusion Assay for Screening
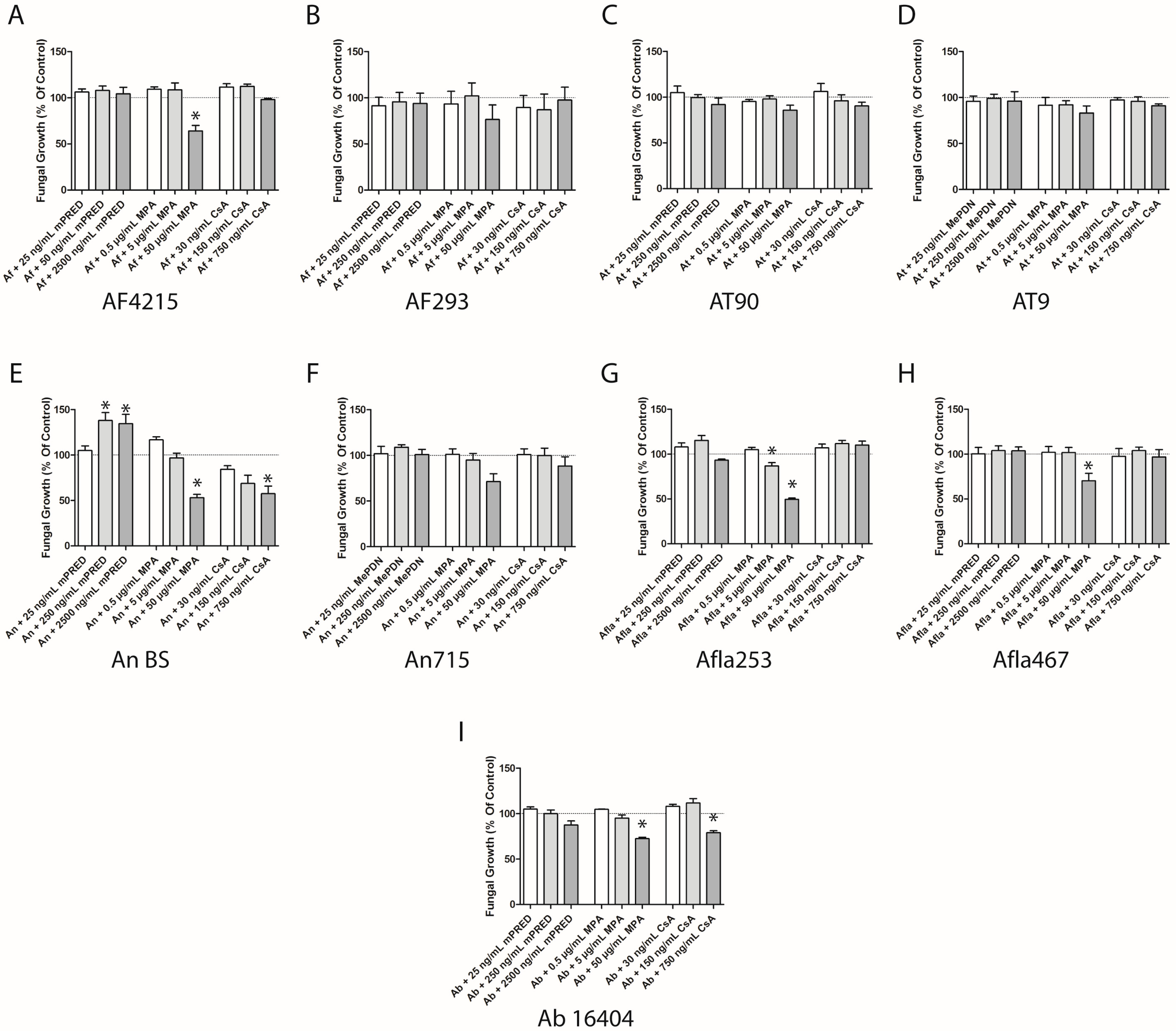
2.2. Immunosuppressive Agents Inhibit the Fungal Growth of Aspergillus Species—Quantitative Assessment of Fungal Cell Density
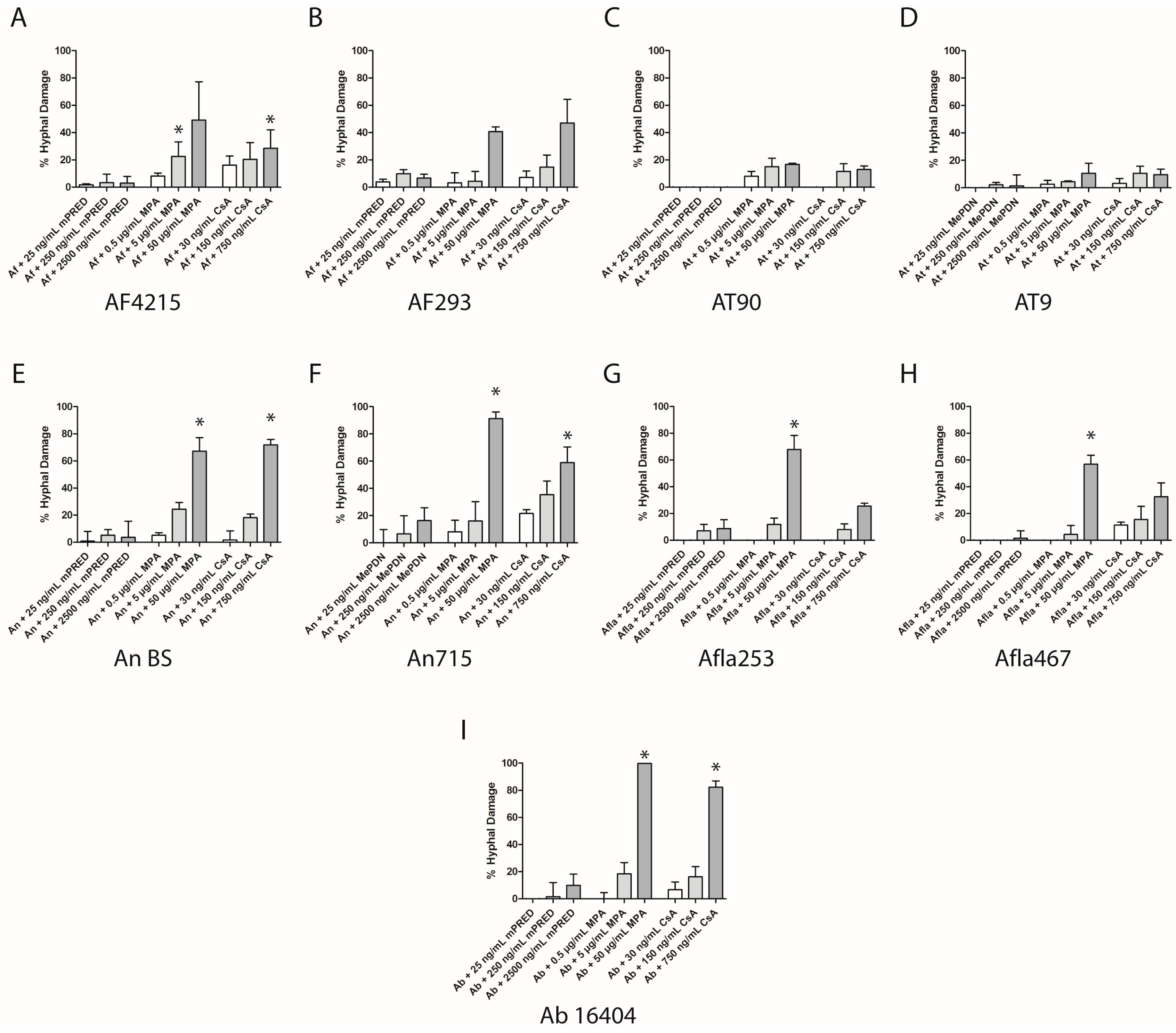
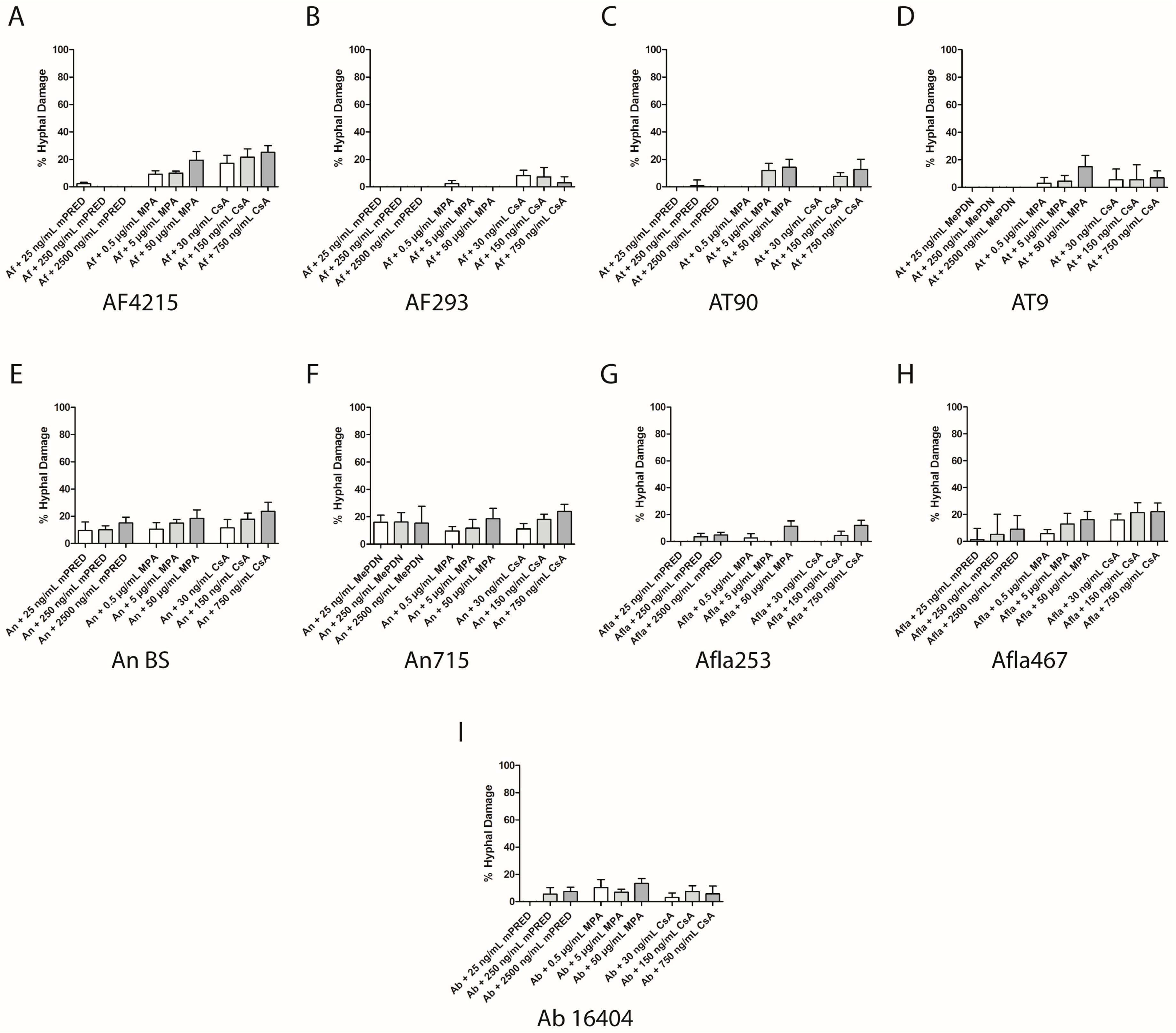
2.3. Immunosuppressive Agents Damage Aspergillus Species
3. Discussion
4. Materials and Methods
4.1. Preparation of Fungi
4.2. Immunosuppressive Agents
4.3. Assessment of the Antifungal Activity of Immunosuppressive Agents
4.3.1. Disc Diffusion Assay
4.3.2. XTT Assay
4.3.3. Assessment of Fungal Cell Density
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lehrnbecher, T.; Foster, C.; Vázquez, N.; Mackall, C.L.; Chanock, S.J. Therapy-induced alterations in host defense in children receiving therapy for cancer. J. Pediatr. Hematol. Oncol. 1997, 19, 399–417. [Google Scholar] [CrossRef]
- Vandevyver, S.; Dejager, L.; Tuckermann, J.; Libert, C. New insights into the anti-inflammatory mechanisms of glucocorticoids: An emerging role for glucocorticoid-receptor-mediated transactivation. Endocrinology 2013, 154, 993–1007. [Google Scholar] [CrossRef]
- Tramsen, L.; Schmidt, S.; Roeger, F.; Schubert, R.; Salzmann-Manrique, E.; Latge, J.-P.; Klingebiel, T.; Lehrnbecher, T. Immunosuppressive Compounds Exhibit Particular Effects on Functional Properties of Human Anti-Aspergillus TH1 Cells. Infect. Immun. 2014, 82, 2649–2656. [Google Scholar] [CrossRef]
- Allison, A.C.; Eugui, E.M. Mycophenolate mofetil and its mechanisms of action. Immunopharmacology 2000, 47, 85–118. [Google Scholar] [CrossRef]
- Allison, A.C. Mechanisms of action of mycophenolate mofetil. Lupus 2005, 14, s2–s8. [Google Scholar] [CrossRef]
- Flores, C.; Fouquet, G.; Moura, I.C.; Maciel, T.T.; Hermine, O. Lessons to Learn From Low-Dose Cyclosporin-A: A New Approach for Unexpected Clinical Applications. Front. Immunol. 2019, 10, 588. [Google Scholar] [CrossRef]
- Miyakoshi, S.; Kusumi, E.; Matsumura, T.; Hori, A.; Murashige, N.; Hamaki, T.; Yuji, K.; Uchida, N.; Masuoka, K.; Wake, A.; et al. Invasive Fungal Infection Following Reduced-Intensity Cord Blood Transplantation for Adult Patients with Hematologic Diseases. Biol. Blood Marrow Transplant. 2007, 13, 771–777. [Google Scholar] [CrossRef]
- Fisher, B.T.; Robinson, P.D.; Lehrnbecher, T.; Steinbach, W.J.; Zaoutis, T.E.; Phillips, B.; Sung, L. Risk Factors for Invasive Fungal Disease in Pediatric Cancer and Hematopoietic Stem Cell Transplantation: A Systematic Review. J. Pediatric Infect. Dis. Soc. 2018, 7, 191–198. [Google Scholar] [CrossRef]
- Steinbach, W.J.; Schell, W.A.; Blankenship, J.R.; Onyewu, C.; Heitman, J.; Perfect, J.R. In Vitro interactions between antifungals and immunosuppressants against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2004, 48, 1664–1669. [Google Scholar] [CrossRef]
- Li, Y.; Sun, S.; Guo, Q.; Ma, L.; Shi, C.; Su, L.; Li, H. In Vitro interaction between azoles and cyclosporin A against clinical isolates of Candida albicans determined by the chequerboard method and time-kill curves. J. Antimicrob. Chemother. 2008, 61, 577–585. [Google Scholar] [CrossRef]
- Kontoyiannis, D.P.; Marr, K.A.; Park, B.J.; Alexander, B.D.; Anaissie, E.J.; Walsh, T.J.; Ito, J.; Andes, D.R.; Baddley, J.W.; Brown, J.M.; et al. Prospective Surveillance for Invasive Fungal Infections in Hematopoietic Stem Cell Transplant Recipients, 2001–2006: Overview of the Transplant-Associated Infection Surveillance Network (TRANSNET) Database. Clin. Infect. Dis. 2010, 50, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Baddley, J.W.; Marr, K.A.; Andes, D.R.; Walsh, T.J.; Kauffman, C.A.; Kontoyiannis, D.P.; Ito, J.I.; Balajee, S.A.; Pappas, P.G.; Moser, S.A. Patterns of Susceptibility of Aspergillus Isolates Recovered from Patients Enrolled in the Transplant-Associated Infection Surveillance Network. J. Clin. Microbiol. 2009, 47, 3271–3275. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Singh, N.; Miller, J.L.; Benjamin, D.K.; Schell, W.A.; Heitman, J.; Perfect, J.R. In Vitro interactions between antifungals and immunosuppressants against Aspergillus fumigatus isolates from transplant and nontransplant patients. Antimicrob. Agents Chemother. 2004, 48, 4922–4925. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.C.; Fox, D.S.; Heitman, J. Calcineurin is required for hyphal elongation during mating and haploid fruiting in Cryptococcus neoformans. EMBO J. 2001, 20, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Prokisch, H.; Yarden, O.; Dieminger, M.; Tropschug, M.; Barthelmess, I.B. Impairment of calcineurin function in Neurospora crassa reveals its essential role in hyphal growth, morphology and maintenance of the apical Ca2+ gradient. Mol. Gen. Genet. 1997, 256, 104–114. [Google Scholar] [CrossRef]
- Wong, S.S.W.; Rasid, O.; Laskaris, P.; Fekkar, A.; Cavaillon, J.M.; Steinbach, W.J.; Ibrahim-Granet, O. Treatment of Cyclosporin A retains host defense against invasive pulmonary aspergillosis in a non-immunosuppressive murine model by preserving the myeloid cell population. Virulence 2017, 8, 1744–1752. [Google Scholar] [CrossRef]
- Punnett, A.; Sung, L.; Price, V.; Das, P.; Diezi, M.; Doyle, J.; Dupuis, L.L. Achievement of target cyclosporine concentrations as a predictor of severe acute graft versus host disease in children undergoing hematopoietic stem cell transplantation and receiving cyclosporine and methotrexate prophylaxis. Ther. Drug Monit. 2007, 29, 750–757. [Google Scholar] [CrossRef]
- Juvvadi, P.R.; Lee, S.C.; Heitman, J.; Steinbach, W.J. Calcineurin in fungal virulence and drug resistance: Prospects for harnessing targeted inhibition of calcineurin for an antifungal therapeutic approach. Virulence 2017, 8, 186–197. [Google Scholar] [CrossRef]
- Oz, H.S.; Hughes, W.T. Novel anti-Pneumocystis carinii effects of the immunosuppressant mycophenolate mofetil in contrast to provocative effects of tacrolimus, sirolimus, and dexamethasone. J. Infect. Dis. 1997, 175, 901–904. [Google Scholar] [CrossRef]
- Husain, S.; Singh, N. The impact of novel immunosuppressive agents on infections in organ transplant recipients and the interactions of these agents with antimicrobials. Clin. Infect. Dis. 2002, 35, 53–61. [Google Scholar] [CrossRef]
- Ritter, M.L.; Pirofski, L. Mycophenolate mofetil: Effects on cellular immune subsets, infectious complications, and antimicrobial activity. Transpl. Infect. Dis. 2009, 11, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.T.C.; Robson, G.D.; Denning, D.W. Hydrocortisone-enhanced growth of Aspergillus spp.: Implications for pathogenesis. Microbiology 1994, 140, 2475–2479. [Google Scholar] [CrossRef] [PubMed]
- Loose, D.S.; Stevens, D.A.; Schurman, D.J.; Feldman, D. Distribution of a corticosteroid-binding protein in Candida and other fungal genera. J. Gen. Microbiol. 1983, 129, 2379–2385. [Google Scholar] [CrossRef] [PubMed]
- Bellanger, A.; Minetos, Y.; Albert, N.; Shirazi, F.; Walsh, T.; Kontoyiannis, D. Glucocorticosteroids do not impact directly growth rate and biomass of Rhizopus arrhizus (syn. R. oryzae) In Vitro. Virulence 2015, 6, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kriesche, H.U.; Friedman, G.; Jacobs, M.; Mulgaonkar, S.; Vaghela, M.; Kaplan, B. Infectious complications in geriatric renal transplant patients: Comparison of two immunosuppressive protocols. Transplantation 1999, 68, 1496–1502. [Google Scholar] [CrossRef] [PubMed]
- Tharayil John, G.; Shankar, V.; Talaulikar, G.; Mathews, M.S.; Abraham Abraham, M.; Punnakuzhathil Thomas, P.; Korula Jacob, C. Epidemiology of systemic mycoses among renal-transplant recipients in India. Transplantation 2003, 75, 1544–1551. [Google Scholar] [CrossRef]
- Narreddy, S.; Manavathu, E.; Chandrasekar, P.H.; Alangaden, G.J.; Revankar, S.G. In Vitro interaction of posaconazole with calcineurin inhibitors and sirolimus against zygomycetes. J. Antimicrob. Chemother. 2010, 65, 701–703. [Google Scholar] [CrossRef]
- Schwarz, P.; Schwarz, P.V.; Felske-Zech, H.; Dannaoui, E. In vitro interactions between isavuconazole and tacrolimus, cyclosporin A or sirolimus against Mucorales. J. Antimicrob. Chemother. 2019, 74, 1921–1927. [Google Scholar] [CrossRef]
- Rohatagi, S.; Barth, J.; Möllmann, H.; Hochhaus, G.; Soldner, A.; Möllmann, C.; Derendorf, H. Pharmacokinetics of methylprednisolone and prednisolone after single and multiple oral administration. J. Clin. Pharmacol. 1997, 37, 916–925. [Google Scholar] [CrossRef]
- Osunkwo, I.; Bessmertny, O.; Harrison, L.; Cheung, Y.K.; Van De Ven, C.; del Toro, G.; Garvin, J.; George, D.; Bradley, M.B.; Wolownik, K.; et al. A pilot study of tacrolimus and mycophenolate mofetil graft-versus-host disease prophylaxis in childhood and adolescent allogeneic stem cell transplant recipients. Biol. Blood Marrow Transplant. 2004, 10, 246–258. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Hanisch, M.; Latgé, J.-P.; Huenecke, S.; Koehl, U.; Lehrnbecher, T. Human natural killer cells exhibit direct activity against Aspergillus fumigatus hyphae, but not against resting conidia. J. Infect. Dis. 2011, 203, 430–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, B.J.; Kim, Y.; Nandakumar, M.P.; Marten, M.R. Quantifying metabolic activity of filamentous fungi using a colorimetric XTT assay. Biotechnol. Prog. 2008, 24, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Loures, F.; Levitz, S. XTT Assay of Antifungal Activity. Bio-Protocol 2015, 5, e1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antachopoulos, C.; Demchok, J.P.; Roilides, E.; Walsh, T.J. Fungal biomass is a key factor affecting polymorphonuclear leucocyte-induced hyphal damage of filamentous fungi. Mycoses 2009, 53, 321–328. [Google Scholar] [CrossRef]
- Antachopoulos, C.; Meletiadis, J.; Roilides, E.; Sein, T.; Sutton, D.A.; Wickes, B.L.; Rinaldi, M.G.; Merz, W.G.; Shea, Y.R.; Walsh, T.J. Relationship between metabolism and biomass of medically important zygomycetes. Med. Mycol. 2006, 44, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Dannaoui, E.; Persat, F.; Monier, M.-F.; Borel, E.; Piens, M.-A.; Picot, S. In-vitro susceptibility of Aspergillus spp. isolates to amphotericin B and itraconazole. J. Antimicrob. Chemother. 1999, 44, 553–555. [Google Scholar] [CrossRef] [Green Version]
- Meletiadis, J.; Stergiopoulou, T.; O’Shaughnessy, E.M.; Peter, J.; Walsh, T.J. Concentration-dependent synergy and antagonism within a triple antifungal drug combination against Aspergillus species: Analysis by a new response surface model. Antimicrob. Agents Chemother. 2007, 51, 2053–2064. [Google Scholar] [CrossRef] [Green Version]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, S.; Hogardt, M.; Demir, A.; Röger, F.; Lehrnbecher, T. Immunosuppressive Compounds Affect the Fungal Growth and Viability of Defined Aspergillus Species. Pathogens 2019, 8, 273. https://doi.org/10.3390/pathogens8040273
Schmidt S, Hogardt M, Demir A, Röger F, Lehrnbecher T. Immunosuppressive Compounds Affect the Fungal Growth and Viability of Defined Aspergillus Species. Pathogens. 2019; 8(4):273. https://doi.org/10.3390/pathogens8040273
Chicago/Turabian StyleSchmidt, Stanislaw, Michael Hogardt, Asuman Demir, Frauke Röger, and Thomas Lehrnbecher. 2019. "Immunosuppressive Compounds Affect the Fungal Growth and Viability of Defined Aspergillus Species" Pathogens 8, no. 4: 273. https://doi.org/10.3390/pathogens8040273
APA StyleSchmidt, S., Hogardt, M., Demir, A., Röger, F., & Lehrnbecher, T. (2019). Immunosuppressive Compounds Affect the Fungal Growth and Viability of Defined Aspergillus Species. Pathogens, 8(4), 273. https://doi.org/10.3390/pathogens8040273