Differential Mortality and High Viral Load in Naive Pacific Oyster Families Exposed to OsHV-1 Suggests Tolerance Rather than Resistance to Infection

 , , , and
, , , and 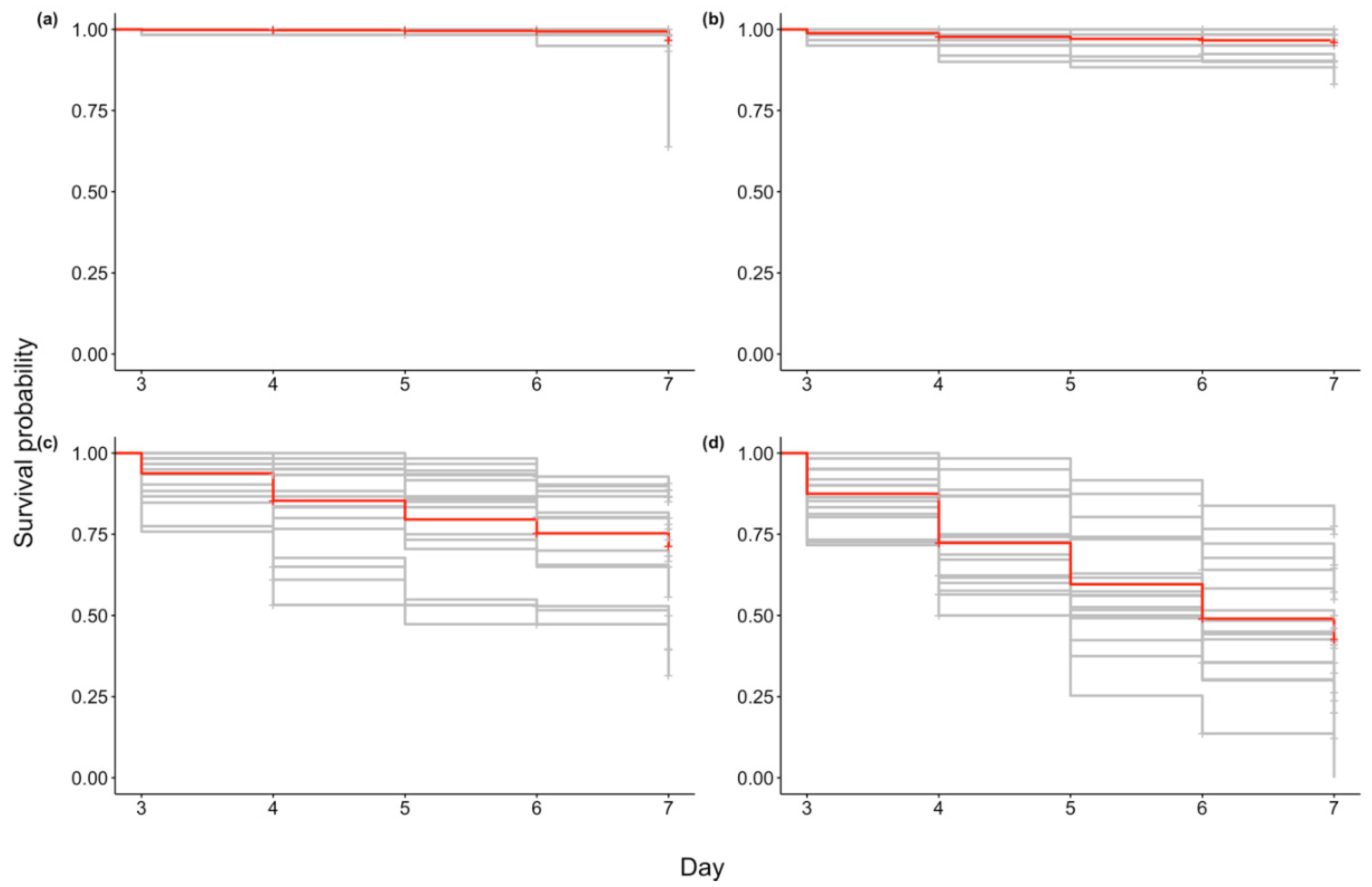{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Inoculum
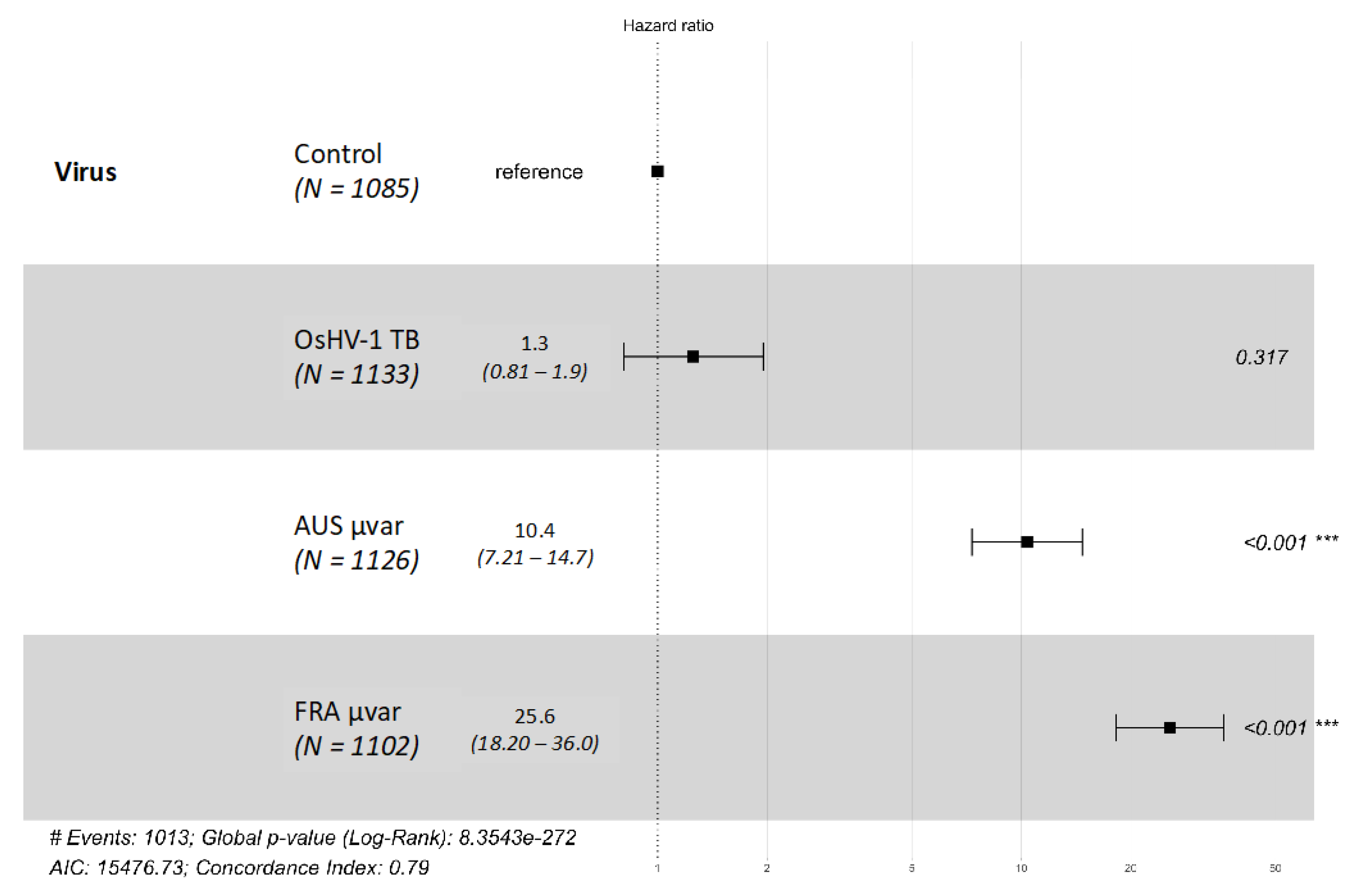
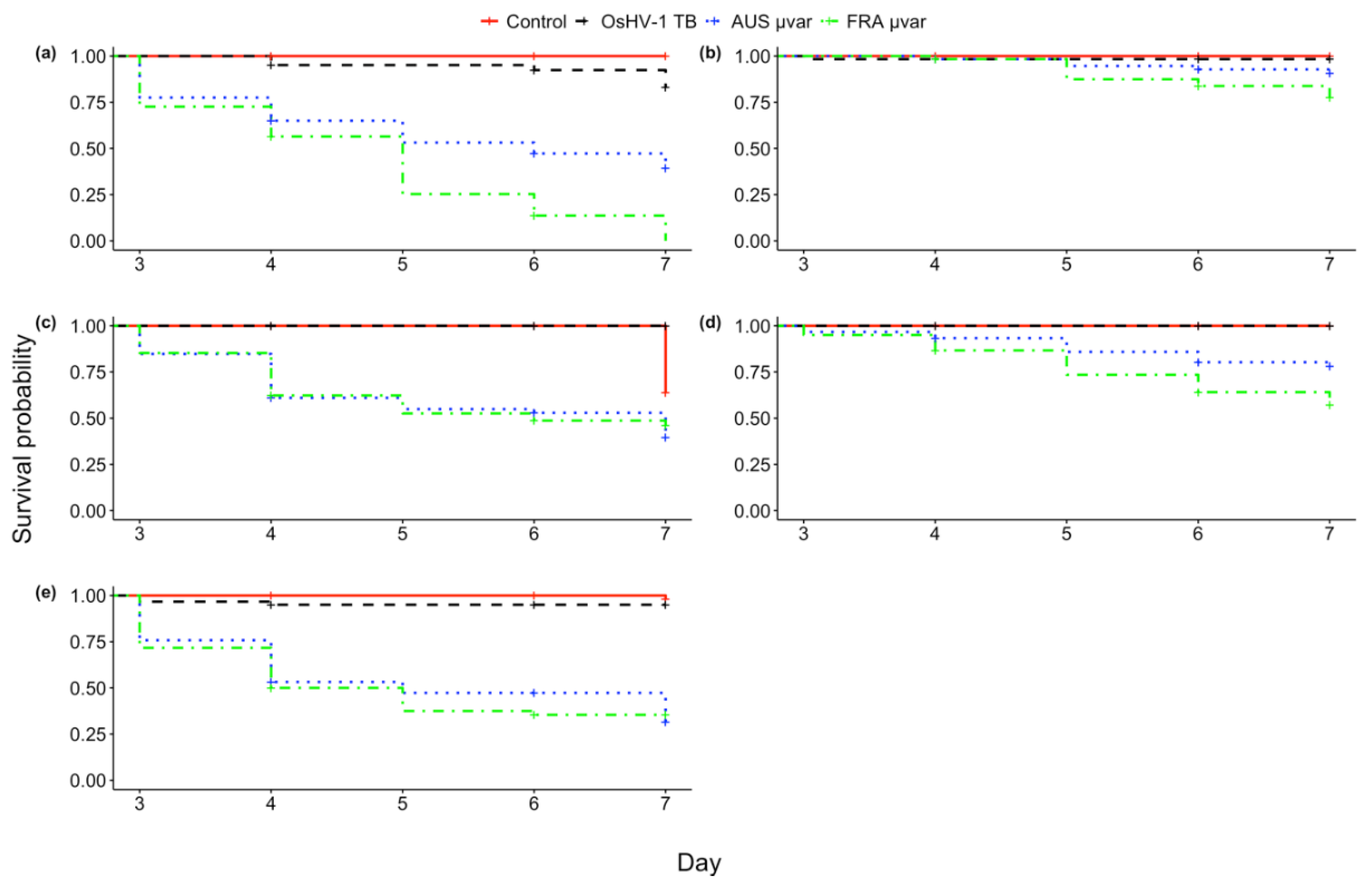
2.2. Mortality
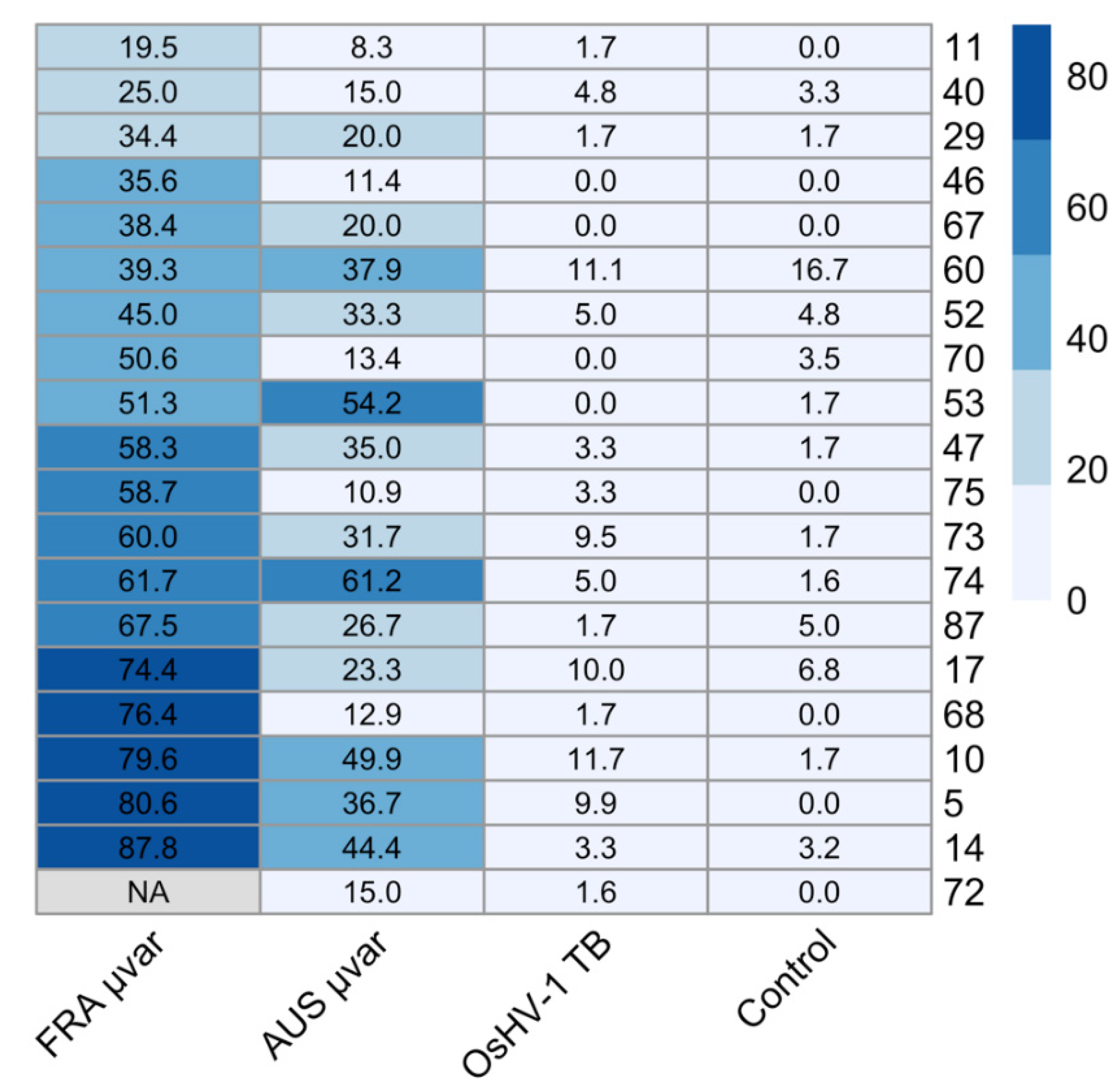
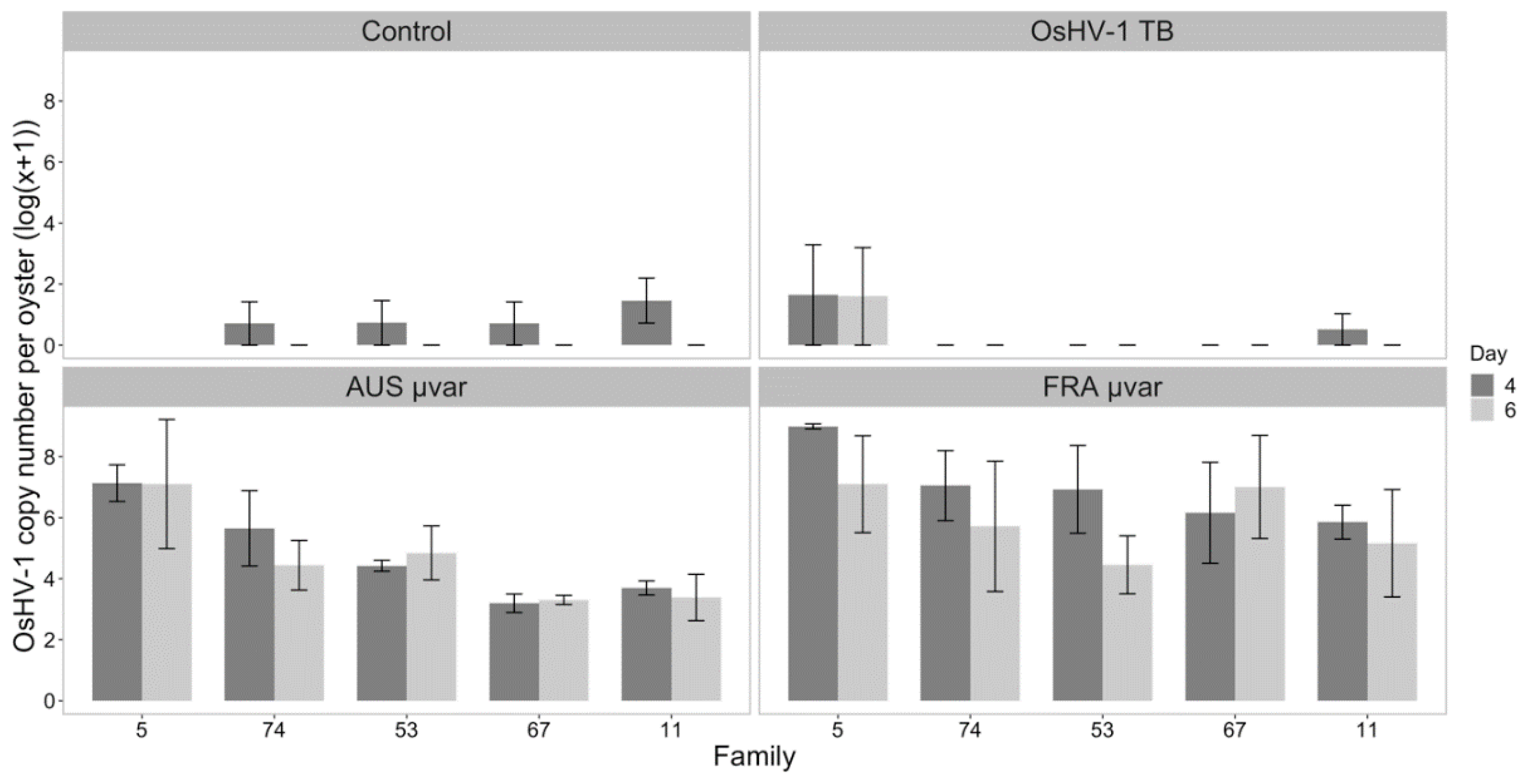
2.3. qPCR of Dead and Live Oysters
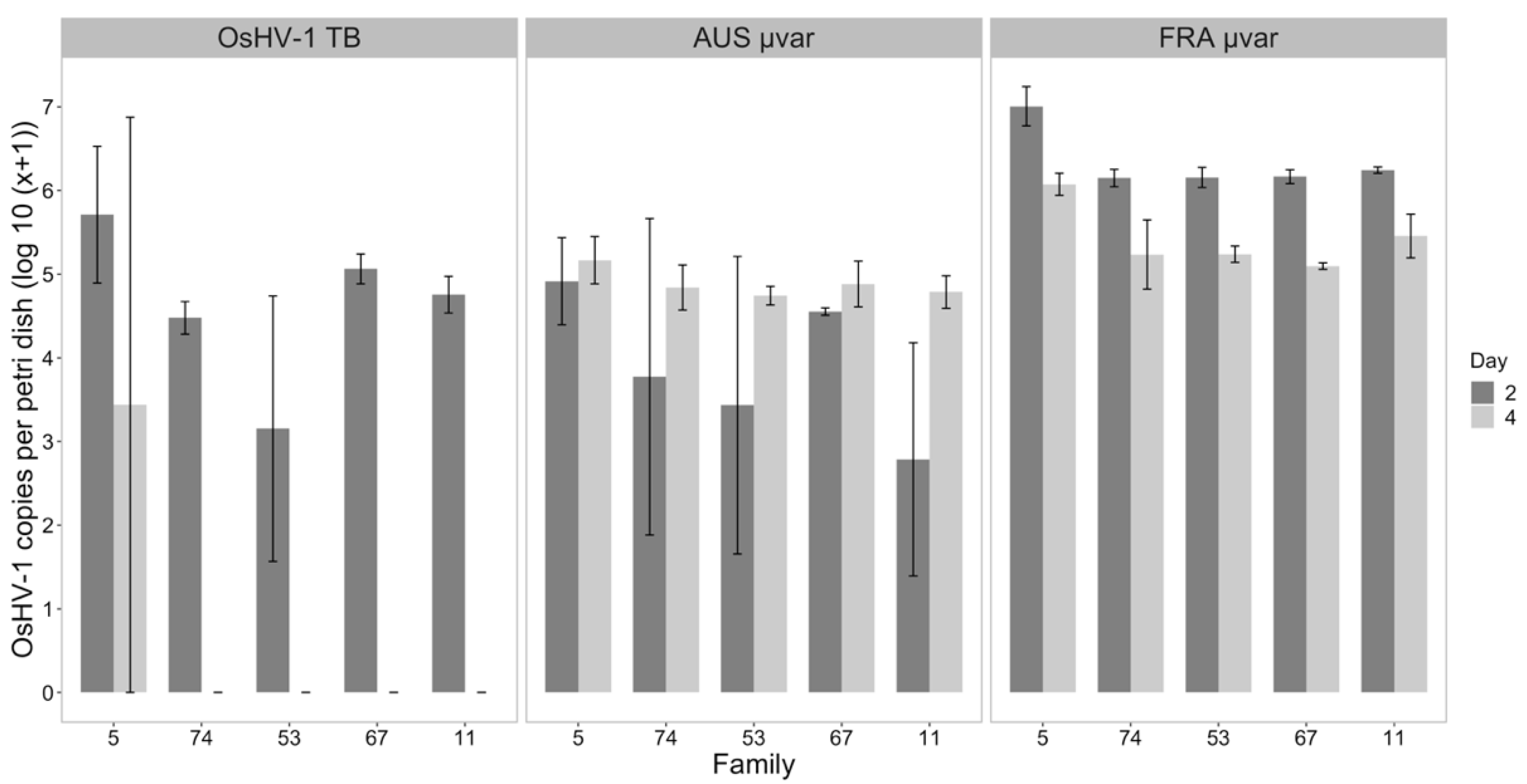
2.4. Water Samples
3. Discussion
4. Materials and Methods
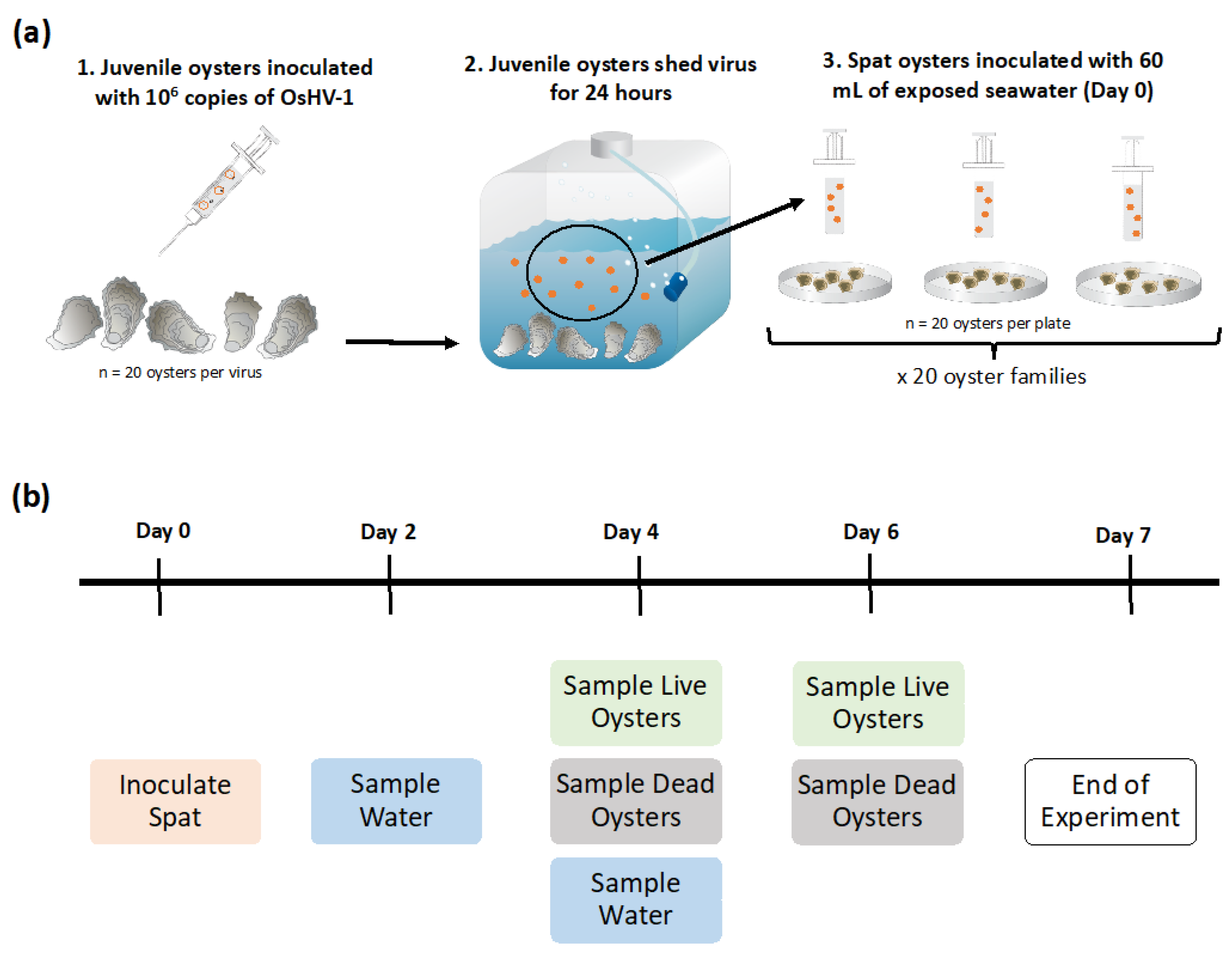
4.1. Oysters Challenged with OsHV-1
4.1.1. Oysters
4.1.2. Inoculum
4.1.3. Infection of Juvenile Oysters
4.2. Oyster and Water Sampling
4.2.1. Mortality Counts
4.2.2. Sampling of Water and Oysters
4.3. Sample Extraction and OsHV-1 Detection via qPCR
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Fishery Statistical Collections: Global Aquaculture Production (Online Query); FAO: Rome, Italy, 2019. [Google Scholar]
- Asche, F.; Anderson, J.L.; Botta, R.; Kumar, G.; Abrahamsen, E.B.; Nguyen, L.T.; Valderrama, D. The Economics of Shrimp Disease. J. Invertebr. Pathol. 2020. [Google Scholar] [CrossRef]
- Rimstad, E.; Markussen, T. Infectious salmon anaemia virus—Molecular biology and pathogenesis of the infection. J. Appl. Microbiol. 2019, 129, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Fernández Robledo, J.; Vasta, G.; Record, N. Protozoan parasites of bivalve molluscs: Literature follows culture. PLoS ONE 2014, 9, 100872. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.R.; Lafferty, K.D. The elusive baseline of marine disease: Are diseases in ocean ecosystems increasing? PLoS Biol. 2004, 2. [Google Scholar] [CrossRef] [Green Version]
- Tracy, A.M.; Pielmeier, M.L.; Yoshioka, R.M.; Heron, S.F.; Harvell, D.C. Increases and decreases in marine disease reports in an era of global change. Proc. R. Soc. B Biol. Sci. 2019, 286. [Google Scholar] [CrossRef] [PubMed]
- Burge, C.A.; Eakin, C.M.; Friedman, C.S.; Froelich, B.; Hershberger, P.K.; Hofmann, E.E.; Petes, L.E.; Prager, K.C.; Weil, E.; Willis, B.L.; et al. Climate change influences on marine infectious diseases: Implications for Management and Society. Ann. Rev. Mar. Sci. 2014, 6, 249–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate Warming and Disease Risks for Terrestrial and Marine Biota. Science 2002, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burge, C.A.; Hershberger, P.K. Climate change can drive marine disease. In Marine Disease Ecology; Behringer, D.C., Silliman, B.R., Lafferty, K.D., Eds.; Oxford University Press: New York, NY, USA, 2020; pp. 83–94. [Google Scholar]
- Lafferty, K.D.; Harvell, C.D.; Conrad, J.M.; Friedman, C.S.; Kent, M.L.; Kuris, A.M.; Powell, E.N.; Rondeau, D.; Saksida, S.M. Infectious Diseases Affect Marine Fisheries and Aquaculture Economics. Ann. Rev. Mar. Sci. 2015, 7, 471–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA 2018 Census of Aquaculture. 2019. Available online: Ahttps://www.nass.usda.gov/Publications/AgCensus/2017/Online_Resources/Aquaculture/index.php (accessed on 16 December 2020).
- Nicolas, J.L.; Comps, M.; Cochennec, N. Herpes-like virus infecting Pacific oyster larvae, Crassostrea gigas. Bull. Eur. Assoc. Fish Pathol. 1992, 12, 11–13. [Google Scholar]
- Renault, T.; Le Deuff, R.M.; Cochennec, N.; Maffart, P. Herpesviruses associated with mortalities among Pacifie oyster, Crassostrea gigas, in France—Comparative study. Rev. Méd. Vél. 1994, 145, 735–742. [Google Scholar]
- Hine, P.M.; Wesney, B.; Hay, B.E. Herpesvirus associated with mortalities among hatchery-reared larval Pacific oysters, Crassostrea gigas. Dis. Aquat. Organ. 1992, 12, 135–142. [Google Scholar] [CrossRef]
- Friedman, C.S.; Estes, R.M.; Stokes, N.A.; Burge, C.A.; Hargove, J.S.; Barber, B.J.; Elston, R.A.; Burreson, E.M.; Reece, K.S. Herpes virus in juvenile Pacific oysters Crassostrea gigas from Tomales Bay, California, coincides with summer mortality episodes. Dis. Aquat. Organ. 2005, 63, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burge, C.A.; Griffin, F.J.; Friedman, C.S. Mortality and herpesvirus infections of the Pacific oyster Crassostrea gigas in Tomales Bay, California, USA. Dis. Aquat. Organ. 2006, 72, 31–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burge, C.A.; Judah, L.R.; Conquest, L.L.; Griffin, F.J.; Cheney, D.P.; Suhrbier, A.; Vadopalas, B.; Olin, P.G.; Renault, T.; Friedman, C.S. Summer seed mortality of the Pacific oyster, Crassostrea gigas Thunberg grown in Tomales Bay, California, USA: The influence of oyster stock, planting time, pathogens, and environmental stressors. J. Shellfish Res. 2007, 26, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Segarra, A.; Pépin, J.F.; Arzul, I.; Morga, B.; Faury, N.; Renault, T. Detection and description of a particular Ostreid herpesvirus 1 genotype associated with massive mortality outbreaks of Pacific oysters, Crassostrea gigas, in France in 2008. Virus Res. 2010, 153, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, A.J.; Trus, B.L.; Cheng, N.; Steven, A.C.; Watson, M.S.; Cunningham, C.; Le Deuff, R.M.; Renault, T. A novel class of herpesvirus with bivalve hosts. J. Gen. Virol. 2005, 86, 41–53. [Google Scholar] [CrossRef]
- OIE. Infection with Ostreid herpesvirus 1 microvariants. In OIE—Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2019. [Google Scholar]
- Garcia, C.; Thébault, A.; Dégremont, L.; Arzul, I.; Miossec, L.; Robert, M.; Chollet, B.; François, C.; Joly, J.P.; Ferrand, S.; et al. Ostreid herpesvirus 1 detection and relationship with Crassostrea gigas spat mortality in France between 1998 and 2006. Vet. Res. 2011, 42. [Google Scholar] [CrossRef] [Green Version]
- Burge, C.A.; Shore-Maggio, A.; Rivlin, N.D. Ecology of emerging infectious diseases of invertebrates. In Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D.I., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 587–625. [Google Scholar]
- Shimahara, Y.; Kurita, J.; Kiryu, I.; Nishioka, T.; Yuasa, K.; Kawana, M.; Kamaishi, T.; Oseko, N. Surveillance of type 1 Ostreid herpesvirus (OsHV-1) variants in Japan. Fish Pathol. 2012, 47, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.Y.; Park, J.J.; Yu, H.J.; Hur, Y.B.; Arzul, I.; Couraleau, Y.; Park, M.A. Ostreid herpesvirus 1 infection in farmed Pacific oyster larvae Crassostrea gigas (Thunberg) in Korea. J. Fish Dis. 2013, 36, 969–972. [Google Scholar] [CrossRef] [Green Version]
- Bai, C.; Wang, C.; Xia, J.; Sun, H.; Zhang, S.; Huang, J. Emerging and endemic types of Ostreid herpesvirus 1 were detected in bivalves in China. J. Invertebr. Pathol. 2015, 124, 98–106. [Google Scholar] [CrossRef]
- Jenkins, C.; Hick, P.; Gabor, M.; Spiers, Z.; Fell, S.A.; Gu, X.; Read, A.; Go, J.; Dove, M.; O’Connor, W.; et al. Identification and characterisation of an ostreid herpesvirus-1 microvariant (OsHV-1 μ-var) in Crassostrea gigas (Pacific oysters) in Australia. Dis. Aquat. Organ. 2013, 105, 109–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bingham, P.; Brangenberg, N.; Williams, R.; van Andel, M. Investigation into the first diagnosis of ostreid herpesvirus type 1 in Pacific oysters. Surveillance 2013, 40, 20–24. [Google Scholar]
- Keeling, S.E.; Brosnahan, C.L.; Williams, R.; Gias, E.; Hannah, M.; Bueno, R.; McDonald, W.L.; Johnston, C. New Zealand juvenile oyster mortality associated with ostreid herpesvirus 1-an opportunistic longitudinal study. Dis. Aquat. Organ. 2014, 109, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, T.; Moreau, P.; Faury, N.; Pepin, J.; Segarra, A.; Webb, S. Analysis of Clinical Ostreid Herpesvirus 1 (Malacoherpesviridae) Specimens by Sequencing Amplified Fragments from Three Virus Genome Areas. J. Virol. 2012, 86, 5942–5947. [Google Scholar] [CrossRef] [Green Version]
- Burge, C.A.; Friedman, C.S.; Kachmar, M.L.; Humphrey, K.L.; Moore, J.D.; Elston, R.A. The first detection of a novel OsHV-1 microvariant in San Diego, California, USA. J. Invertebr. Pathol. 2020. under review. [Google Scholar]
- Peeler, E.J.; Reese, R.A.; Cheslett, D.L.; Geoghegan, F.; Power, A.; Thrush, M.A. Investigation of mortality in Pacific oysters associated with Ostreid herpesvirus-1μVar in the Republic of Ireland in 2009. Prev. Vet. Med. 2012, 105, 136–143. [Google Scholar] [CrossRef]
- Whittington, R.J.; Paul-Pont, I.; Evans, O.; Hick, P.; Dhand, N.K. Counting the dead to determine the source and transmission of the marine herpesvirus OsHV-1 in Crassostrea gigas. Vet. Res. 2018, 49. [Google Scholar] [CrossRef] [Green Version]
- Paul-Pont, I.; Evans, O.; Dhand, N.K.; Rubio, A.; Coad, P.; Whittington, R.J. Descriptive epidemiology of mass mortality due to Ostreid herpesvirus-1 (OsHV-1) in commercially farmed Pacific oysters (Crassostrea gigas) in the Hawkesbury River estuary, Australia. Aquaculture 2014, 422–423, 146–159. [Google Scholar] [CrossRef] [Green Version]
- Evans, O.; Paul-Pont, I.; Hick, P.; Whittington, R.J. A simple centrifugation method for improving the detection of Ostreid herpesvirus-1 (OsHV-1) in natural seawater samples with an assessment of the potential for particulate attachment. J. Virol. Methods 2014, 210, 59–66. [Google Scholar] [CrossRef]
- Dégremont, L.; Garcia, C.; Allen, S.K. Genetic improvement for disease resistance in oysters: A review. J. Invertebr. Pathol. 2015, 131, 226–241. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.D. Oyster diseases in Chesapeake Bay. Mar. Fish. Rev. 1979, 41, 45–53. [Google Scholar]
- Ragone Calvo, L.M.; Calvo, G.W.; Burreson, E.M. Dual disease resistance in a selectively bred Eastern oyster, Crassostrea virginica, strain tested in Chesapeake Bay. Aquaculture 2003, 220, 69–87. [Google Scholar] [CrossRef]
- Frank-Lawale, A.; Allen, S.K.; Dégremont, L. Breeding and Domestication of Eastern Oyster (Crassostrea virginica) Lines for Culture in the Mid-Atlantic, Usa: Line Development and Mass Selection for Disease Resistance. J. Shellfish Res. 2014, 33, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Dégremont, L. Evidence of herpesvirus (OsHV-1) resistance in juvenile Crassostrea gigas selected for high resistance to the summer mortality phenomenon. Aquaculture 2011, 317, 94–98. [Google Scholar] [CrossRef] [Green Version]
- Dégremont, L. Size and genotype affect resistance to mortality caused by OsHV-1 in Crassostrea gigas. Aquaculture 2013, 416–417, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Dégremont, L.; Nourry, M.; Maurouard, E. Mass selection for survival and resistance to OsHV-1 infection in Crassostrea gigas spat in field conditions: Response to selection after four generations. Aquaculture 2015, 446, 111–121. [Google Scholar] [CrossRef]
- Kube, P.; Dove, M.; Cunningham, M.; Kirkland, P.; O’Connor, W.; Elliott, N. Genetic variation in Pacific oysters (Crassostrea gigas) for resistance to Ostreid herpesvirus-1. Aquaculture 2017, 472, S119–S120. [Google Scholar]
- Gutierrez, A.P.; Symonds, J.; King, N.; Steiner, K.; Bean, T.P.; Houston, R.D. Potential of genomic selection for improvement of resistance to Ostreid Herpes virus in Pacific oyster (Crassostrea gigas). Anim. Genet. 2020, 51, 249–257. [Google Scholar] [CrossRef]
- Burge, C.A.; Reece, K.S.; Dhar, A.K.; Kirkland, P.; Morga, B.; Dégremont, L.; Faury, N.; Wippel, B.J.T.; MacIntyre, A.; Friedman, C.S. First comparison of French and Australian OsHV-1 µvars by bath exposure. Dis. Aquat. Organ. 2020, 138, 137–144. [Google Scholar] [CrossRef]
- Friedman, C.S.; Reece, K.S.; Wippel, B.J.T.; Agnew, M.V.; Dégremont, L.; Dhar, A.K.; Kirkland, P.; MacIntyre, A.; Morga, B.; Robison, C.; et al. Unraveling concordant and varying responses of oyster species to Ostreid Herpesvirus 1 variants. Sci. Total Environ. 2020, 739. [Google Scholar] [CrossRef]
- de Melo, C.M.R.; Durland, E.; Langdon, C. Improvements in desirable traits of the Pacific oyster, Crassostrea gigas, as a result of five generations of selection on the West Coast, USA. Aquaculture 2016, 460, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Divilov, K.; Schoolfield, B.; Morga, B.; Dégremont, L.; Burge, C.A.; Cortez, D.M.; Friedman, C.S.; Fleener, G.B.; Dumbauld, B.R.; Langdon, C. First evaluation of resistance to both a California OsHV-1 variant and a French OsHV-1 microvariant in Pacific oysters. BMC Genet. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Le Deuff, R.M.; Nicolas, L.; Renault, T.; Cochennec, N. Experimental transmission of a herpes-like virus to axenic larvae of pacific oyster, Crassostrea gigas. Bull. Eur. Assoc. Fish Pathol 1994, 14, 69. [Google Scholar]
- Arzul, I.; Renault, T.; Lipart, C. Experimental herpes-like viral infections in marine bivalves: Demonstration of interspecies transmission. Dis. Aquat. Organ. 2001, 46, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Burge, C.A.; Friedman, C.S. Quantifying Ostreid Herpesvirus (OsHV-1) Genome Copies and Expression during Transmission. Microb. Ecol. 2012, 63, 596–604. [Google Scholar] [CrossRef]
- Burge, C.A.; Agnew, M.V. Exposure of Juvenile Crassostrea gigas to OsHV-1 TB. Unpublished work. 2018. [Google Scholar]
- Boots, M.; Bowers, R.G. Three Mechanisms of Host Resistance to Microparasites-Avoidance, Recovery and Tolerance-Show Different Evolutionary Dynamics. J. Theor. Biol. 1999, 201, 13–23. [Google Scholar] [CrossRef]
- Friedman, C.S.; Wight, N.; Crosson, L.M.; VanBlaricom, G.R.; Lafferty, K.D. Reduced disease in black abalone following mass mortality: Phage therapy and natural selection. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Arzul, I.; Renault, T.; Thébault, A.; Gérard, A. Detection of oyster herpesvirus DNA and proteins in asymptomatic Crassostrea gigas adults. Virus Res. 2002, 84, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Oden, E.; Martenot, C.; Berthaux, M.; Travaillé, E.; Malas, J.P.; Houssin, M. Quantification of ostreid herpesvirus 1 (OsHV-1) in Crassostrea gigas by real-time PCR: Determination of a viral load threshold to prevent summer mortalities. Aquaculture 2011, 317, 27–31. [Google Scholar] [CrossRef]
- Le Deuff, R.M.; Renault, T.; Gérard, A. Effects of temperature on herpes-like virus detection among hatchery-reared larval Pacific oyster Crassostrea gigas. Dis. Aquat. Organ. 1996, 24, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Barbosa-Solomieu, V.; Dégremont, L.; Vázquez-Juárez, R.; Ascencio-Valle, F.; Boudry, P.; Renault, T. Ostreid Herpesvirus 1 (OsHV-1) detection among three successive generations of Pacific oysters (Crassostrea gigas). Virus Res. 2005, 107, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Kancharla, R.S.; Hanson, L.A. Production and shedding of channel catfish virus (CCV) and thymidine kinase negative CCV in immersion exposed channel catfish fingerlings. Dis. Aquat. Organ. 1996, 27, 25–34. [Google Scholar] [CrossRef]
- Yuasa, K.; Ito, T.; Sano, M. Effect of Water Temperature on Mortality and Virus Shedding in Carp Experimentally Infected with Koi Herpesvirus. Fish Pathol. 2008, 43, 83–85. [Google Scholar] [CrossRef] [Green Version]
- Faisal, M.; Purbayu, M.; Shavalier, M.A.; Marsh, T.L.; Loch, T.P. Shedding of the salmonid herpesvirus-3 by infected lake trout (Salvelinus namaycush). Viruses 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Petton, B.; Bruto, M.; James, A.; Labreuche, Y.; Alunno-Bruscia, M.; Le Roux, F. Crassostrea gigas mortality in France: The usual suspect, a herpes virus, may not be the killer in this polymicrobial opportunistic disease. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Ugalde, S.C.; Preston, J.; Ogier, E.; Crawford, C. Analysis of farm management strategies following herpesvirus (OsHV-1) disease outbreaks in Pacific oysters in Tasmania, Australia. Aquaculture 2018, 495, 179–186. [Google Scholar] [CrossRef]
- Pernet, F.; Barret, J.; Le Gall, P.; Corporeau, C.; Dégremont, L.; Lagarde, F.; Pépin, J.F.; Keck, N. Mass mortalities of Pacific oysters Crassostrea gigas reflect infectious diseases and vary with farming practices in the Mediterranean Thau lagoon, France. Aquac. Environ. Interact. 2012, 2, 215–237. [Google Scholar] [CrossRef] [Green Version]
- de Kantzow, M.; Hick, P.; Becker, J.A.; Whittington, R.J. Effect of water temperature on mortality of Pacific oysters Crassostrea gigas associated with microvariant ostreid herpesvirus 1 (OsHV-1 µVar). Aquac. Environ. Interact. 2016, 8, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Delisle, L.; Petton, B.; Burguin, J.F.; Morga, B.; Corporeau, C.; Pernet, F. Temperature modulate disease susceptibility of the Pacific oyster Crassostrea gigas and virulence of the Ostreid herpesvirus type 1. Fish Shellfish Immunol. 2018, 80, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Dégremont, L.; Morga, B.; Trancart, S.; Pépin, J.F. Resistance to OsHV-1 infection in Crassostrea gigas larvae. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Elston, R.A. Shellfish Health Laboratory Report. Unpublished work. 2018. [Google Scholar]
- Elston, R.A.; Aquatechnics Inc., Seqium, DC, USA; Burge, C.A.; UMBC, Baltimore, MD, USA. Personal communication, 2020.
- Sawyer, L.; Hog Island Oyster Company, Eurkea, California, USA. Personal communication, 2018.
- Kirkland, P.D.; Hick, P.; Gu, X. Development of a Laboratory Model for Infectious Challenge of Pacific Oysters (Crassostrea gigas) with Ostreid Herpesvirus Type-1; Elizabeth Macarthur Agriculture Institute: Sydney, Australia, 2015.
- Paul-Pont, I.; Evans, O.; Dhand, N.; Whittington, R. Experimental infections of Pacific oyster Crassostrea gigas using the Australian osterid herpesvirus-1 (OsHV-1) µVar strain. Dis. Aquat. Organ. 2015, 113, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 16 December 2020).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps. R package persion 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 16 December 2020).
- Therneau, T. A Package for Survival Analysis in R, R package version 3.2-3; 2020. Available online: https://CRAN.R-project.org/package=survival (accessed on 16 December 2020).
- Kassambara, L.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using “ggplot2”, R package version 0.4.6; 2019. Available online: https://CRAN.R-project.org/package=survminer (accessed on 16 December 2020).
- Bartoń, K. Package “MuMIn”—Multi-Model Inference. Available online: https://cran.r-project.org/web/packages/MuMIn/index.html (accessed on 16 December 2020).
- Kabacoff, R.I. Basic Statistics. In R in Action: Data Analysis and Graphics in R; Manning Publications Co.: Shelter Island, NY, USA, 2015. [Google Scholar]
- Agnew, M.V.; Friedman, C.S.; Langdon, C.; Divilov, K.; Schoolfield, B.; Morga, B.; Degrémont, L.; Dhar, A.K.; Kirkland, P.; Dumbauld, B.; et al. Data and statistical code associated with: Differential mortality and high viral load in naïve Pacific oyster families exposed to OsHV-1 suggests tolerance rather than resistance to infection. Figshare 2020. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agnew, M.V.; Friedman, C.S.; Langdon, C.; Divilov, K.; Schoolfield, B.; Morga, B.; Degremont, L.; Dhar, A.K.; Kirkland, P.; Dumbauld, B.; et al. Differential Mortality and High Viral Load in Naive Pacific Oyster Families Exposed to OsHV-1 Suggests Tolerance Rather than Resistance to Infection. Pathogens 2020, 9, 1057. https://doi.org/10.3390/pathogens9121057
Agnew MV, Friedman CS, Langdon C, Divilov K, Schoolfield B, Morga B, Degremont L, Dhar AK, Kirkland P, Dumbauld B, et al. Differential Mortality and High Viral Load in Naive Pacific Oyster Families Exposed to OsHV-1 Suggests Tolerance Rather than Resistance to Infection. Pathogens. 2020; 9(12):1057. https://doi.org/10.3390/pathogens9121057
Chicago/Turabian StyleAgnew, M. Victoria, Carolyn S. Friedman, Christopher Langdon, Konstantin Divilov, Blaine Schoolfield, Benjamin Morga, Lionel Degremont, Arun K. Dhar, Peter Kirkland, Brett Dumbauld, and et al. 2020. "Differential Mortality and High Viral Load in Naive Pacific Oyster Families Exposed to OsHV-1 Suggests Tolerance Rather than Resistance to Infection" Pathogens 9, no. 12: 1057. https://doi.org/10.3390/pathogens9121057
APA StyleAgnew, M. V., Friedman, C. S., Langdon, C., Divilov, K., Schoolfield, B., Morga, B., Degremont, L., Dhar, A. K., Kirkland, P., Dumbauld, B., & Burge, C. A. (2020). Differential Mortality and High Viral Load in Naive Pacific Oyster Families Exposed to OsHV-1 Suggests Tolerance Rather than Resistance to Infection. Pathogens, 9(12), 1057. https://doi.org/10.3390/pathogens9121057