Current Developments in the Epidemiology and Control of Enzootic Bovine Leukosis as Caused by Bovine Leukemia Virus

 , , , and
, , , and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Increasing Prevalence
3. Transmission
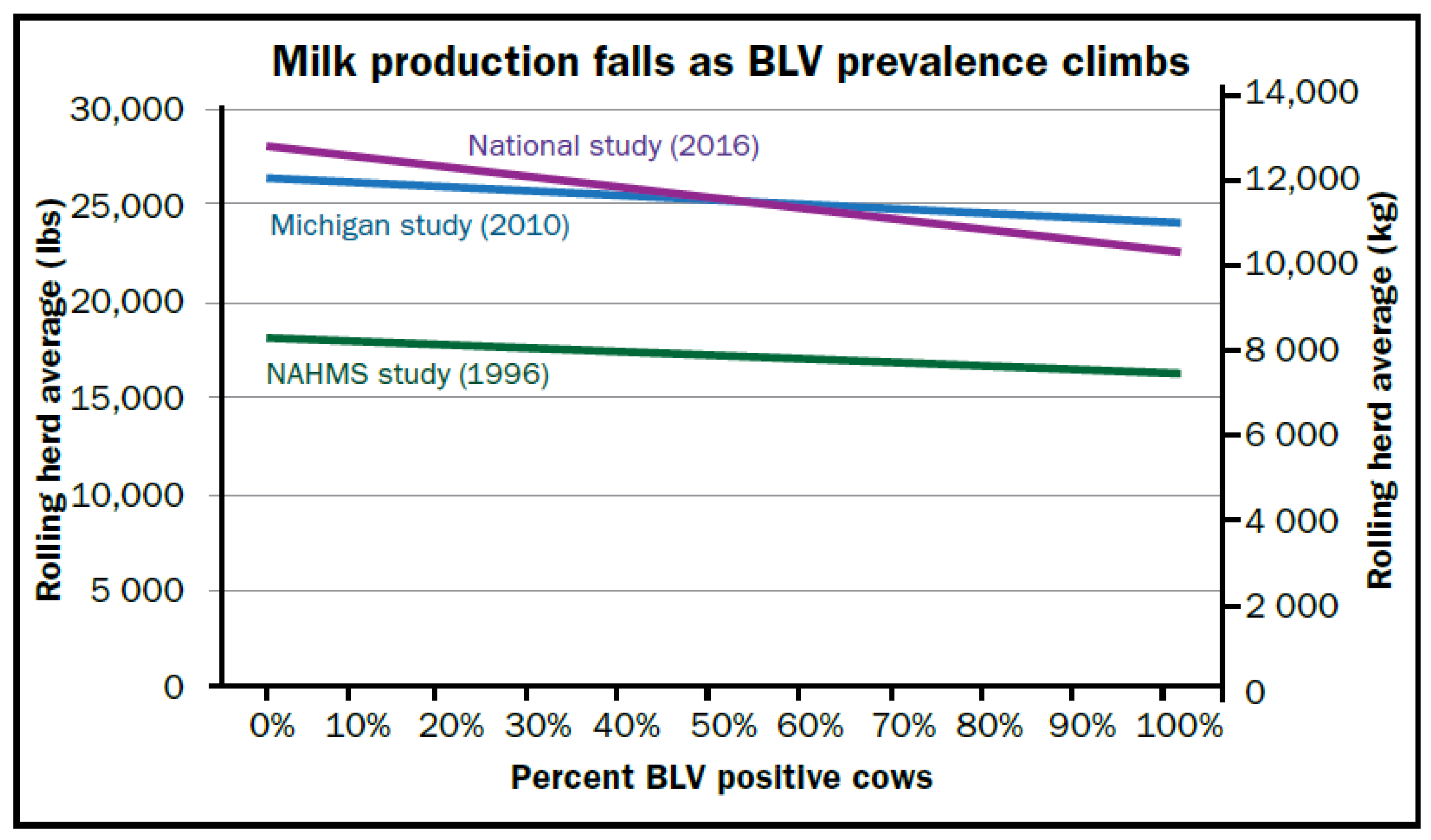
4. Economic Impact in Dairy Cattle
5. Altered Immunity
6. Proviral Load (PVL)
7. BLV in Beef Cattle
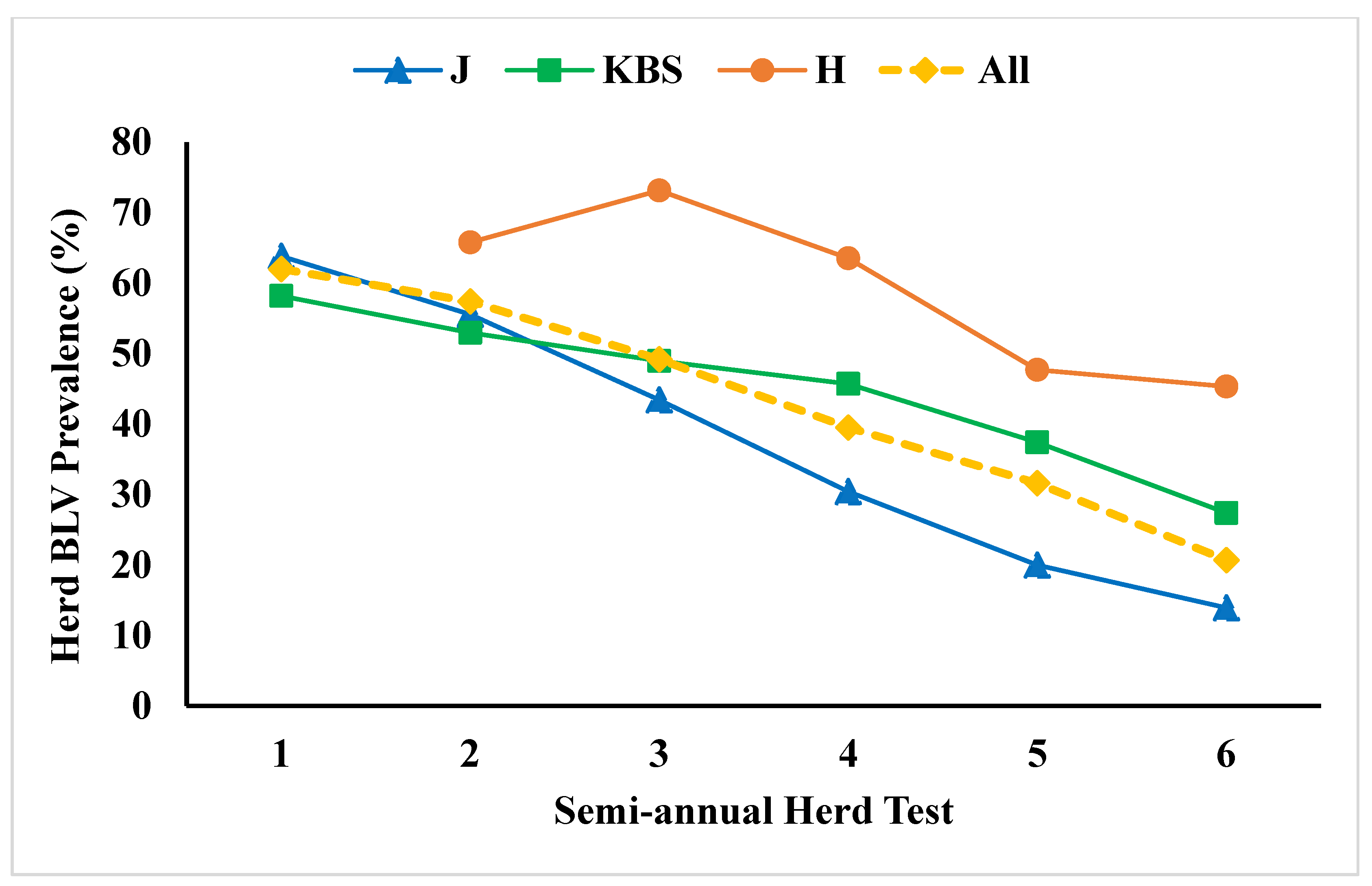
8. Controlling BLV
9. Host Genetics
10. The BLV Viral Genome
11. Human Health
12. Vaccines
13. Issues for Future Inquiry
- What causes a minority of cattle to develop extremely high proviral load and to what extent is this determined by their genetics or the time at which they become infected (in utero or very early in life via colostrum)?
- How important are host genetics, whereby certain alleles predispose an animal to a high or low proviral load, antibody titer and/or severity of pathology?
- Why do first lactation animals seemingly avoid the milk production loss and increased risk of culling that are associated with older ELISA-positive cows?
- The severe inbreeding of Holstein dairy cattle is such that the effective population size is <50 animals, and they would be considered critically endangered if they were a wild animal species [94]. This raises the question of how long a provirus-infected lymphocyte could survive and propagate upon being introduced into an almost genetically identical cow compared to a genetically distinct cow.
- Attribution studies are needed to determine the percentage of new BLV infections that come from direct contact, biting fly transmission, shared hypodermic needles, etc.
- The role of free BLV RNA in disease transmission should be definitively established.
- Controlled studies are required to measure the extent to which BLV infection contributes to the causation of the major dairy diseases such as mastitis, lameness, infertility, etc.
- Longitudinal studies should determine how commonly animals are infected with BLV as calves, but the virus becomes sequestered until the stress of birth and lactation. Genetic methods should be developed to determine if newly recognized infections in cows are usually caused by the same strain found in their dams over two years ago, or the same strains found in their contemporaries.
- The role of BLV in the induction and pathogenesis of human breast cancer and other human diseases should be definitively established.
- What role does microRNA play in the pathogenesis of BLV, and can it be used for diagnosis or prognosis?
- A safe and efficacious BLV vaccine should be developed that allows differentiation between vaccinates and natural infection.
- The likely return on investment for BLV control and eradication should be determined for test and removal using ELISA and by using ELISA screening with proviral load follow-up testing.
14. BLV and the Future of Animal Agriculture
15. Conclusions
Funding
Conflicts of Interest
References
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benavides, B.B.; Quevedo, D.A.C.; de La Cruz, M.F.S. Epidemiological study of bovine leukemia virus in dairy cows in six herds in the municipality of Pasto, Nariño. Rev. Lasallista Investig. 2013, 10, 18–23. [Google Scholar]
- Úsuga-Monroy, C.; Echeverri, J.; López-Herrera, H. Molecular diagnosis of bovine Leukemia Virus in a population of Holstein cows, Colombia. Arch. Zootec. 2015, 64, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, C.; Bonilla, J.A.; Dolz, G.; Rodriguez, L.R.; Herrero, L.; Bolaños, E.; Cortéz, M.R.; Moreno, E. Bovine Leukaemia-virus Infection in Costa Rica. J. Veter. Med. Ser. B 1995, 42, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Haghparast, A.; Mohammadi, G.; Tabatabaeizadeh, S.-E. Prevalence of bovine leukemia virus (BLV) infection in the northeast of Iran. Veter. Res. Forum Int. Q. J. 2014, 5, 135–139. [Google Scholar]
- Nekoei, S.; Hafshejani, T.T.; Doosti, A.; Khamesipour, F. Molecular detection of bovine leukemia virus in peripheral blood of Iranian cattle, camel and sheep. Pol. J. Veter. Sci. 2015, 18, 703–707. [Google Scholar] [CrossRef] [Green Version]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.-I.; Tsutsui, T. Nationwide Survey of Bovine Leukemia Virus Infection among Dairy and Beef Breeding Cattle in Japan from 2009–2011. J. Veter. Med. Sci. 2013, 75, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Ochirkhuu, N.; Konnai, S.; Odbileg, R.; Nishimori, A.; Okagawa, T.; Murata, S.; Ohashi, K. Detection of bovine leukemia virus and identification of its genotype in Mongolian cattle. Arch. Virol. 2015, 161, 985–991. [Google Scholar] [CrossRef]
- Polat, M.; Ohno, A.; Takeshima, S.-N.; Kim, J.; Kikuya, M.; Matsumoto, Y.; Mingala, C.N.; Onuma, M.; Aida, Y. Detection and molecular characterization of bovine leukemia virus in Philippine cattle. Arch. Virol. 2014, 160, 285–296. [Google Scholar] [CrossRef]
- Ndou, R.V.; Sejesho, F.; Dzoma, B.M.; Motsei, L.; Nyirenda, M.; Bakunzi, F.R. A Serosurvey of the Prevalence of Enzootic Bovine Leukosis in the Mafikeng Area of the North West Province of South Africa. J. Hum. Ecol. 2011, 36, 53–55. [Google Scholar] [CrossRef]
- Şevik, M.; Avcı, O.; Ince, Ö.B. An 8-year longitudinal sero-epidemiological study of bovine leukaemia virus (BLV) infection in dairy cattle in Turkey and analysis of risk factors associated with BLV seropositivity. Trop. Anim. Health Prod. 2015, 47, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Walrand, F.; Fumoux, F.; Roelants, G.; Parodi, A.-L.; Levy, D. Incidence of bovine leukemia virus-specific antibodies in west african cattle. Int. J. Cancer 1986, 37, 619–621. [Google Scholar] [CrossRef]
- Monti, G.; Schrijver, R.; Beier, D. Genetic diversity and spread of Bovine leukaemia virus isolates in Argentine dairy cattle. Arch. Virol. 2004, 150, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Nekouei, O.; VanLeeuwen, J.; Sanchez, J.; Kelton, D.; Tiwari, A.; Keefe, G. Herd-level risk factors for infection with bovine leukemia virus in Canadian dairy herds. Prev. Veter. Med. 2015, 119, 105–113. [Google Scholar] [CrossRef] [PubMed]
- USDA Bovine Leukosis Virus (BLV) on U.S. Dairy Operations 2007. Available online: https://web.archive.org/web/20190205034903/https://www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy07/Dairy07_is_BLV.pdf (accessed on 4 February 2019).
- LaDronka, R.M.; Ainsworth, S.; Wilkins, M.J.; Norby, B.; Byrem, T.M.; Bartlett, P.C. Prevalence of Bovine Leukemia Virus Antibodies in US Dairy Cattle. Veter. Med. Int. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Commission. Commission Implementing Decision 2016/168. Off. J. Eur. Union 2016, 32, 153–157. [Google Scholar]
- White, T.L.; Moore, D.A. Reasons for whole carcass condemnations of cattle in the United States and implications for producer education and veterinary intervention. J. Am. Veter. Med. Assoc. 2009, 235, 937–941. [Google Scholar] [CrossRef]
- Emanuelson, U.; Scherling, K.; Pettersson, H. Relationships between herd bovine leukemia virus infection status and reproduction, disease incidence, and productivity in Swedish dairy herds. Prev. Veter. Med. 1992, 12, 121–131. [Google Scholar] [CrossRef]
- Brenner, J.; Van-Haam, M.; Savir, D.; Trainin, Z. The implication of BLV infection in the productivity, reproductive capacity and survival rate of a dairy cow. Veter. Immunol. Immunopathol. 1989, 22, 299–305. [Google Scholar] [CrossRef]
- Trainin, Z.; Brenner, J.; Meirom, R.; Ungar-Waron, H. Detrimental effect of bovine leukemia virus (BLV) on the immunological state of cattle. Veter. Immunol. Immunopathol. 1996, 54, 293–302. [Google Scholar] [CrossRef]
- Haredasht, S.A.; Vidal, G.; Edmondson, A.; Moore, D.; Silva-Del-Río, N.; Martínez-López, B. Characterization of the Temporal Trends in the Rate of Cattle Carcass Condemnations in the US and Dynamic Modeling of the Condemnation Reasons in California With a Seasonal Component. Front. Veter. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Pollari, F.L.; Wangsuphachart, V.L.; Digiacomo, R.F.; Evermann, J.F. Effects of bovine leukemia virus infection on production and reproduction in dairy cattle. Can. J. Veter. Res. 1992, 56, 289–295. [Google Scholar]
- Huber, N.L.; Digiacomo, R.F.; Evermann, J.F.; Studer, E. Bovine leukemia virus infection in a large Holstein herd: Prospective comparison of production and reproductive performance in antibody-negative and antibody-positive cows. Am. J. Veter. Res. 1981, 42, 1477–1481. [Google Scholar]
- Sorge, U.S.; Lissemore, K.; Cantin, R.; Kelton, D.F. Short communication: Milk ELISA status for bovine leukosis virus infection is not associated with milk production in dairy cows. J. Dairy Sci. 2011, 94, 5062–5064. [Google Scholar] [CrossRef] [Green Version]
- Nekouei, O.; VanLeeuwen, J.; Stryhn, H.; Kelton, D.; Keefe, G. Lifetime effects of infection with bovine leukemia virus on longevity and milk production of dairy cows. Prev. Veter. Med. 2016, 133, 1–9. [Google Scholar] [CrossRef]
- Norby, B.; Bartlett, P.C.; Byrem, T.M.; Erskine, R.J. Effect of infection with bovine leukemia virus on milk production in Michigan dairy cows. J. Dairy Sci. 2016, 99, 2043–2052. [Google Scholar] [CrossRef]
- Ott, S.L.; Johnson, R.; Wells, S.J. Association between bovine-leukosis virus seroprevalence and herd-level productivity on US dairy farms. Prev. Veter. Med. 2003, 61, 249–262. [Google Scholar] [CrossRef]
- Erskine, R.J.; Bartlett, P.C.; Byrem, T.M.; Render, C.L.; Febvay, C.; Houseman, J.T. Association between bovine leukemia virus, production, and population age in Michigan dairy herds. J. Dairy Sci. 2012, 95, 727–734. [Google Scholar] [CrossRef]
- Wu, D.; Murakami, K.; Morooka, A.; Jin, H.; Inoshima, Y.; Sentsui, H. In vivo transcription of bovine leukemia virus and bovine immunodeficiency-like virus. Virus Res. 2003, 97, 81–87. [Google Scholar] [CrossRef]
- Bartlett, P.C.; Norby, B.; Byrem, T.M.; Parmelee, A.; Ledergerber, J.T.; Erskine, R.J. Bovine leukemia virus and cow longevity in Michigan dairy herds. J. Dairy Sci. 2013, 96, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.K.; Pelzer, K.D.; Johnson, Y.J. Economic implications of bovine leukemia virus infection in mid-Atlantic dairy herds. J. Am. Veter. Med. Assoc. 2003, 223, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Frie, M.C.; Coussens, P.M. Bovine leukemia virus: A major silent threat to proper immune responses in cattle. Veter. Immunol. Immunopathol. 2015, 163, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Erskine, R.J.; Bartlett, P.C.; Sabo, K.M.; Sordillo, L.M. Bovine Leukemia Virus Infection in Dairy Cattle: Effect on Serological Response to Immunization against J5Escherichia coliBacterin. Veter. Med. Int. 2011, 2011, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frie, M.C.; Sporer, K.R.; Wallace, J.C.; Maes, R.K.; Sordillo, L.M.; Bartlett, P.C.; Coussens, P.M. Reduced humoral immunity and atypical cell-mediated immunity in response to vaccination in cows naturally infected with bovine leukemia virus. Veter. Immunol. Immunopathol. 2016, 182, 125–135. [Google Scholar] [CrossRef]
- Frie, M.C.; Sporer, K.R.B.; Benitez, O.J.; Wallace, J.C.; Droscha, C.J.; Bartlett, P.C.; Coussens, P.M. Dairy Cows Naturally Infected with Bovine Leukemia Virus Exhibit Abnormal B- and T-Cell Phenotypes after Primary and Secondary Exposures to Keyhole Limpet Hemocyanin. Front. Veter. Sci. 2017, 4, 112. [Google Scholar] [CrossRef]
- Puentes, R.; De Brun, L.; Algorta, A.; Da Silva, V.; Mansilla, F.; Sacco, G.; Llambí, S.; Capozzo, A.V. Evaluation of serological response to foot-and-mouth disease vaccination in BLV infected cows. BMC Veter. Res. 2016, 12, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Blagitz, M.G.; Souza, F.N.; Batista, C.F.; Azevedo, L.F.F.; Sanchez, E.M.R.; Diniz, S.A.; Silva, M.X.; Haddad, J.P.; Della Libera, A.M.M.P. Immunological implications of bovine leukemia virus infection. Res. Veter. Sci. 2017, 114, 109–116. [Google Scholar] [CrossRef]
- Della Libera, A.M.M.P.; Souza, F.N.; Batista, C.F.; Santos, B.P.; De Azevedo, L.F.F.; Ramos-Sanchez, E.M.; Diniz, S.A.; Silva, M.X.; Haddad, J.P.; Blagitz, M.G. Effects of bovine leukemia virus infection on milk neutrophil function and the milk lymphocyte profile. Veter. Res. 2015, 46, 2. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Kitamura-Muramatsu, Y.; Saito, S.; Ishizaki, H.; Nakano, M.; Haga, S.; Matoba, K.; Ohno, A.; Murakami, H.; Takeshima, S.-N.; et al. Detection of the BLV provirus from nasal secretion and saliva samples using BLV-CoCoMo-qPCR-2: Comparison with blood samples from the same cattle. Virus Res. 2015, 210, 248–254. [Google Scholar] [CrossRef]
- Ruggiero, V.J. Field Studies on the Control of Bovine Leukemia Virus in Dairy Cows. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, 2019. [Google Scholar]
- Ruggiero, V.J.; Dziuba, M.; Bartlett, P.C.; Coussens, P.M. Untitled BLV-Immunoglobulin Paper. Unpublished manuscript in preparation. 2020. [Google Scholar]
- Onuma, M.; Sagata, N.; Okada, K.; Ogawa, Y.; Ikawa, Y.; Oshima, K. Integration of Bovine Leukemia Virus DNA in the Genomes of Bovine Lymphosarcoma Cells. Microbiol. Immunol. 1982, 26, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Juliarena, M.A.; Gutierrez, S.E.; Ceriani, C. Determination of proviral load in bovine leukemia virus–infected cattle with and without lymphocytosis. Am. J. Veter. Res. 2007, 68, 1220–1225. [Google Scholar] [CrossRef]
- Fechner, H.; Kurg, A.; Geue, L.; Blankenstein, P.; Mewes, G.; Ebner, D.; Beier, D. Evaluation of Polymerase Chain Reaction (PCR) Application in Diagnosis of Bovine Leukaemia Virus (BLV) Infection in Naturally Infected Cattle. J. Veter. Med. Ser. B 1996, 43, 621–630. [Google Scholar] [CrossRef]
- Klintevall, K.; Ballagi-Pordány, A.; Näslund, K.; Belák, S. Bovine leukaemia virus: Rapid detection of proviral DNA by nested PCR in blood and organs of experimentally infected calves. Veter. Microbiol. 1994, 42, 191–204. [Google Scholar] [CrossRef]
- Kaplan, J.E.; Khabbaz, R.F.; Murphy, E.L.; Hermansen, S.; Roberts, C.; Lal, R.; Heneine, W.; Wright, D.; Matijas, L.; Thomson, R.; et al. Male-to-Female Transmission of Human T-Cell Lymphotropic Virus Types I and II: Association with Viral Load. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996, 12, 193–201. [Google Scholar] [CrossRef]
- Percher, F.; Jeannin, P.; Martin-Latil, S.; Gessain, A.; Afonso, P.V.; Vidy, A.; Ceccaldi, P.-E. Mother-to-Child Transmission of HTLV-1 Epidemiological Aspects, Mechanisms and Determinants of Mother-to-Child Transmission. Viruses 2016, 8, 40. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, G.; Álvarez, I.; Politzki, R.; Lomónaco, M.; Santos, M.J.D.; Rondelli, F.; Fondevila, N.; Trono, K. Natural progression of Bovine Leukemia Virus infection in Argentinean dairy cattle. Veter. Microbiol. 2011, 151, 255–263. [Google Scholar] [CrossRef]
- Rodríguez, S.M.; Florins, A.; Gillet, N.; De Brogniez, A.; Sánchez-Alcaraz, M.T.; Boxus, M.; Boulanger, F.; Gutiérrez, G.; Trono, K.; Alvarez, I.; et al. Preventive and Therapeutic Strategies for Bovine Leukemia Virus: Lessons for HTLV. Viruses 2011, 3, 1210–1248. [Google Scholar] [CrossRef]
- Ohno, A.; Takeshima, S.-N.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus Res. 2015, 210, 283–290. [Google Scholar] [CrossRef]
- Buxton, B.A.; Schultz, R.D. Factors affecting the infectivity of lymphocytes from cattle with bovine leukosis virus. Can. J. Comp. Med. Rev. Can. Med. Comp. 1984, 48, 365–369. [Google Scholar]
- Mammerickx, M.; Portetelle, D.; De Clercq, K.K.; Burny, A. Experimental transmission of enzootic bovine leukosis to cattle, sheep and goats: Infectious doses of blood and incubation period of the disease. Leuk. Res. 1987, 11, 353–358. [Google Scholar] [CrossRef]
- Molloy, J.B.; Dimmock, C.K.; Eaves, F.W.; Bruyeres, A.G.; Cowley, J.A.; Ward, W.H. Control of bovine leukaemia virus transmission by selective culling of infected cattle on the basis of viral antigen expression in lymphocyte cultures. Veter. Microbiol. 1994, 39, 323–333. [Google Scholar] [CrossRef]
- Gutiérrez, G.; Alvarez, I.; Merlini, R.; Rondelli, F.; Trono, K. Dynamics of perinatal bovine leukemia virus infection. BMC Veter. Res. 2014, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Juliarena, M.A.; Barrios, C.N.; Ceriani, M.C.; Esteban, E.N. Hot topic: Bovine leukemia virus (BLV)-infected cows with low proviral load are not a source of infection for BLV-free cattle. J. Dairy Sci. 2016, 99, 4586–4589. [Google Scholar] [CrossRef] [Green Version]
- Mekata, H.; Sekiguchi, S.; Konnai, S.; Kirino, Y.; Horii, Y.; Norimine, J. Horizontal transmission and phylogenetic analysis of bovine leukemia virus in two districts of Miyazaki, Japan. J. Veter. Med. Sci. 2015, 77, 1115–1120. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, H.C. Transmission and Progression of Bovine Leukemia Virus. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, 2020. [Google Scholar]
- Benitez, O.J.; Norby, B.; Bartlett, P.C.; Maeroff, J.E.; Grooms, D.L. Impact of bovine leukemia virus infection on beef cow longevity. Prev. Veter. Med. 2020, 181, 105055. [Google Scholar] [CrossRef]
- Benitez, O.J.; Roberts, J.N.; Norby, B.; Bartlett, P.C.; Takeshima, S.-N.; Watanuki, S.; Aida, Y.; Grooms, D.L. Breeding bulls as a potential source of bovine leukemia virus transmission in beef herds. J. Am. Veter. Med. Assoc. 2019, 254, 1335–1340. [Google Scholar] [CrossRef]
- Bendixen, H.J. Preventive measures in cattle leukemia: Leukosis enzootica bovis*. Ann. N. Y. Acad. Sci. 2006, 108, 1241–1267. [Google Scholar] [CrossRef]
- Ruggiero, V.J.; Bartlett, P.C. Single-use hypodermic needles and obstetric sleeves failed to reduce bovine leukemia virus transmission in three dairy herds. Bov. Pract. 2019, 53, 128–133. [Google Scholar] [CrossRef]
- Ruggiero, V.; Norby, B.; Benitez, O.; Hutchinson, H.; Sporer, K.; Droscha, C.; Swenson, C.; Bartlett, P. Controlling bovine leukemia virus in dairy herds by identifying and removing cows with the highest proviral load and lymphocyte counts. J. Dairy Sci. 2019, 102, 9165–9175. [Google Scholar] [CrossRef] [Green Version]
- Taxis, T.M.; Sporer, K.R.B.; Droscha, C.J.; Niles, D.; DeJong, T.; Swenson, C.L.; Bartlett, P.C. Reducing bovine leukemia virus prevalence on a large midwestern dairy farm by using lymphocyte counts, ELISA antibody testing, and proviral load. Bov. Pract. 2020. under review. [Google Scholar]
- Hayashi, T.; Mekata, H.; Sekiguchi, S.; Kirino, Y.; Mitoma, S.; Honkawa, K.; Horii, Y.; Norimine, J. Cattle with the BoLA class II DRB3*0902 allele have significantly lower bovine leukemia proviral loads. J. Veter. Med. Sci. 2017, 79, 1552–1555. [Google Scholar] [CrossRef] [Green Version]
- Miyasaka, T.; Takeshima, S.-N.; Jimba, M.; Matsumoto, Y.; Kobayashi, N.; Matsuhashi, T.; Sentsui, H.; Aida, Y. Identification of bovine leukocyte antigen class II haplotypes associated with variations in bovine leukemia virus proviral load in Japanese Black cattle. Tissue Antigens 2013, 81, 72–82. [Google Scholar] [CrossRef]
- Lo, C.-W.; Borjigin, L.; Saito, S.; Fukunaga, K.; Saitou, E.; Okazaki, K.; Mizutani, T.; Wada, S.; Takeshima, S.-N.; Aida, Y. BoLA-DRB3 Polymorphism is Associated with Differential Susceptibility to Bovine Leukemia Virus-Induced Lymphoma and Proviral Load. Viruses 2020, 12, 352. [Google Scholar] [CrossRef] [Green Version]
- Carignano, H.; Beribe, M.J.; Caffaro, M.E.; Amadio, A.; Nani, J.P.; Gutierrez, G.; Alvarez, I.; Trono, K.; Miretti, M.M.; Poli, M. BOLA-DRB3gene polymorphisms influence bovine leukaemia virus infection levels in Holstein and Holstein × Jersey crossbreed dairy cattle. Anim. Genet. 2017, 48, 420–430. [Google Scholar] [CrossRef]
- Maccari, G.; Robinson, J.; Ballingall, K.; Guethlein, L.A.; Grimholt, U.; Kaufman, J.; Ho, C.-S.; De Groot, N.G.; Flicek, P.; Bontrop, R.E.; et al. IPD-MHC 2.0: An improved inter-species database for the study of the major histocompatibility complex. Nucleic Acids Res. 2017, 45, D860–D864. [Google Scholar] [CrossRef]
- Abt, D.A.; Marshak, R.R.; Kulp, H.W.; Pollock, R.J. Studies on the Relationship Between Lymphocytosis and Bovine Leukosis. In Proceedings of the Comparative Leukemia Research 1969, 4th Inernational Symposium, Cherry Hill, NJ, USA, September 1969; Dutcher, R., Ed.; Karger Publishers: New York, NY, USA, 1970; pp. 527–536. [Google Scholar]
- Lewin, H.A.; Bernoco, D. Evidence for BoLA-linked resistance and susceptibility to subclinical progression of bovine leukaemia virus infection. Anim. Genet. 1986, 17, 197–207. [Google Scholar] [CrossRef]
- Xu, A.; van Eijk, M.J.; Park, C.; Lewin, H.A. Polymorphism in BoLA-DRB3 exon 2 correlates with resistance to persistent lymphocytosis caused by bovine leukemia virus. J. Immunol. 1993, 151, 6977–6985. [Google Scholar]
- Udina, I.G.; Karamysheva, E.E.; Turkova, S.O.; Orlova, A.R.; Sulimova, G.E. Genetic mechanisms of resistance and susceptibility to leukemia in Ayrshire and black pied cattle breeds determined by allelic distribution of gene Bola-DRB3. Генетика 2003, 39, 306–317. [Google Scholar] [CrossRef]
- Juliarena, M.A.; Poli, M.; Sala, L.; Ceriani, C.; Gutierrez, S.; Dolcini, G.L.; Rodrguez, E.M.; Mario, B.; Rodrguez-Dubra, C.; Esteban, E.; et al. Association of BLV infection profiles with alleles of theBoLA-DRB3.2gene. Anim. Genet. 2008, 39, 432–438. [Google Scholar] [CrossRef]
- Takeshima, S.-N.; Ohno, A.; Aida, Y. Bovine leukemia virus proviral load is more strongly associated with bovine major histocompatibility complex class II DRB3 polymorphism than with DQA1 polymorphism in Holstein cow in Japan. Retrovirology 2019, 16, 1–6. [Google Scholar] [CrossRef]
- Pluta, A.; Willems, L.; Douville, R.N.; Kuźmak, J. Effects of Naturally Occurring Mutations in Bovine Leukemia Virus 5′-LTR and Tax Gene on Viral Transcriptional Activity. Pathogens 2020, 9, 836. [Google Scholar] [CrossRef] [PubMed]
- Zyrianova, I.M.; Kovalchuk, S. Bovine leukemia virus tax gene/Tax protein polymorphism and its relation to Enzootic Bovine Leukosis. Virulence 2020, 11, 80–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frie, M.C.; Droscha, C.J.; Greenlick, A.E.; Coussens, P.M. MicroRNAs Encoded by Bovine Leukemia Virus (BLV) Are Associated with Reduced Expression of B Cell Transcriptional Regulators in Dairy Cattle Naturally Infected with BLV. Front. Veter. Sci. 2018, 4, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safari, R.; Jacques, J.-R.; Brostaux, Y.; Willems, L. Ablation of non-coding RNAs affects bovine leukemia virus B lymphocyte proliferation and abrogates oncogenesis. PLOS Pathog. 2020, 16, e1008502. [Google Scholar] [CrossRef]
- Cuesta, L.M.; Lendez, P.A.; Farias, M.V.N.; Dolcini, G.L.; Ceriani, M.C. Can Bovine Leukemia Virus Be Related to Human Breast Cancer? A Review of the Evidence. J. Mammary Gland. Biol. Neoplasia 2018, 23, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Buehring, G.C.; Philpott, S.M.; Choi, K.Y. Humans Have Antibodies Reactive with Bovine Leukemia Virus. AIDS Res. Hum. Retrovir. 2003, 19, 1105–1113. [Google Scholar] [CrossRef]
- Buehring, G.C.; Delaney, A.; Shen, H.; Chu, D.L.; Razavian, N.; Schwartz, D.A.; Demkovich, Z.R.; Bates, M.N. Bovine leukemia virus discovered in human blood. BMC Infect. Dis. 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Khalilian, M.; Hosseini, S.M.; Madadgar, O. Bovine leukemia virus detected in the breast tissue and blood of Iranian women. Microb. Pathog. 2019, 135, 103566. [Google Scholar] [CrossRef]
- Burridge, M.J. The zoonotic potential of bovine leukemia virus. Veter. Res. Commun. 1981, 5, 117–126. [Google Scholar] [CrossRef]
- Gillet, N.A.; Willems, L. Whole genome sequencing of 51 breast cancers reveals that tumors are devoid of bovine leukemia virus DNA. Retrovirology 2016, 13, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Khoury, J.D.; Tannir, N.M.; Williams, M.D.; Chen, Y.; Yao, H.; Zhang, J.; Thompson, E.J.; Meric-Bernstam, F.; Medeiros, L.J.; Weinstein, J.N.; et al. Landscape of DNA Virus Associations across Human Malignant Cancers: Analysis of 3775 Cases Using RNA-Seq. J. Virol. 2013, 87, 8916–8926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.-W.; Alaei-Mahabadi, B.; Samuelsson, T.; Lindh, M.; Larsson, E. The landscape of viral expression and host gene fusion and adaptation in human cancer. Nat. Commun. 2013, 4, 2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltzell, K.; Shen, H.M.; Krishnamurthy, S.; Sison, J.D.; Nuovo, G.J.; Buehring, G.C. Bovine leukemia virus linked to breast cancer but not coinfection with human papillomavirus: Case-control study of women in Texas. Cancer 2017, 124, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Buehring, G.C.; Shen, H.; Schwartz, D.A.; Lawson, J.S. Bovine leukemia virus linked to breast cancer in Australian women and identified before breast cancer development. PLoS ONE 2017, 12, e0179367. [Google Scholar] [CrossRef] [Green Version]
- Schwingel, D.; Andreolla, A.P.; Erpen, L.M.S.; Frandoloso, R.; Kreutz, L.C. Bovine leukemia virus DNA associated with breast cancer in women from South Brazil. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef]
- Buehring, G.C.; Shen, H.M.; Jensen, H.M.; Jin, D.L.; Hudes, M.; Block, G. Exposure to Bovine Leukemia Virus Is Associated with Breast Cancer: A Case-Control Study. PLoS ONE 2015, 10, e0134304. [Google Scholar] [CrossRef] [Green Version]
- Olaya-Galán, N.N.; Corredor-Figueroa, A.P.; Guzmán-Garzón, T.C.; Ríos-Hernandez, K.S.; Salas-Cárdenas, S.P.; Patarroyo, M.A.; Gutierrez, M.F. Bovine leukaemia virus DNA in fresh milk and raw beef for human consumption. Epidemiol. Infect. 2017, 145, 3125–3130. [Google Scholar] [CrossRef] [Green Version]
- Abdala, A.; Alvarez, I.; Brossel, H.; Calvinho, L.; Carignano, H.; Franco, L.; Gazon, H.; Gillissen, C.; Hamaidia, M.; Hoyos, C.; et al. BLV: Lessons on vaccine development. Retrovirology 2019, 16, 1–6. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, M.; Undark. From Two Bulls, Nine Million Dairy Cows. Sci. Am. 2019. Available online: https://www.scientificamerican.com/article/from-two-bulls-nine-million-dairy-cows/ (accessed on 17 December 2020).
- RethinkX Food and Agriculture Report—RethinkX. Available online: https://www.rethinkx.com/food-and-agriculture (accessed on 29 July 2020).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartlett, P.C.; Ruggiero, V.J.; Hutchinson, H.C.; Droscha, C.J.; Norby, B.; Sporer, K.R.B.; Taxis, T.M. Current Developments in the Epidemiology and Control of Enzootic Bovine Leukosis as Caused by Bovine Leukemia Virus. Pathogens 2020, 9, 1058. https://doi.org/10.3390/pathogens9121058
Bartlett PC, Ruggiero VJ, Hutchinson HC, Droscha CJ, Norby B, Sporer KRB, Taxis TM. Current Developments in the Epidemiology and Control of Enzootic Bovine Leukosis as Caused by Bovine Leukemia Virus. Pathogens. 2020; 9(12):1058. https://doi.org/10.3390/pathogens9121058
Chicago/Turabian StyleBartlett, Paul C., Vickie J. Ruggiero, Holden C. Hutchinson, Casey J. Droscha, Bo Norby, Kelly R. B. Sporer, and Tasia M. Taxis. 2020. "Current Developments in the Epidemiology and Control of Enzootic Bovine Leukosis as Caused by Bovine Leukemia Virus" Pathogens 9, no. 12: 1058. https://doi.org/10.3390/pathogens9121058
APA StyleBartlett, P. C., Ruggiero, V. J., Hutchinson, H. C., Droscha, C. J., Norby, B., Sporer, K. R. B., & Taxis, T. M. (2020). Current Developments in the Epidemiology and Control of Enzootic Bovine Leukosis as Caused by Bovine Leukemia Virus. Pathogens, 9(12), 1058. https://doi.org/10.3390/pathogens9121058