The Examination of Viral Characteristics of HIV-1 CRF07_BC and Its Potential Interaction with Extracellular Galectin-3

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. CRF07_BC Was Predominant in HIV-1(+) Injecting Drug Users
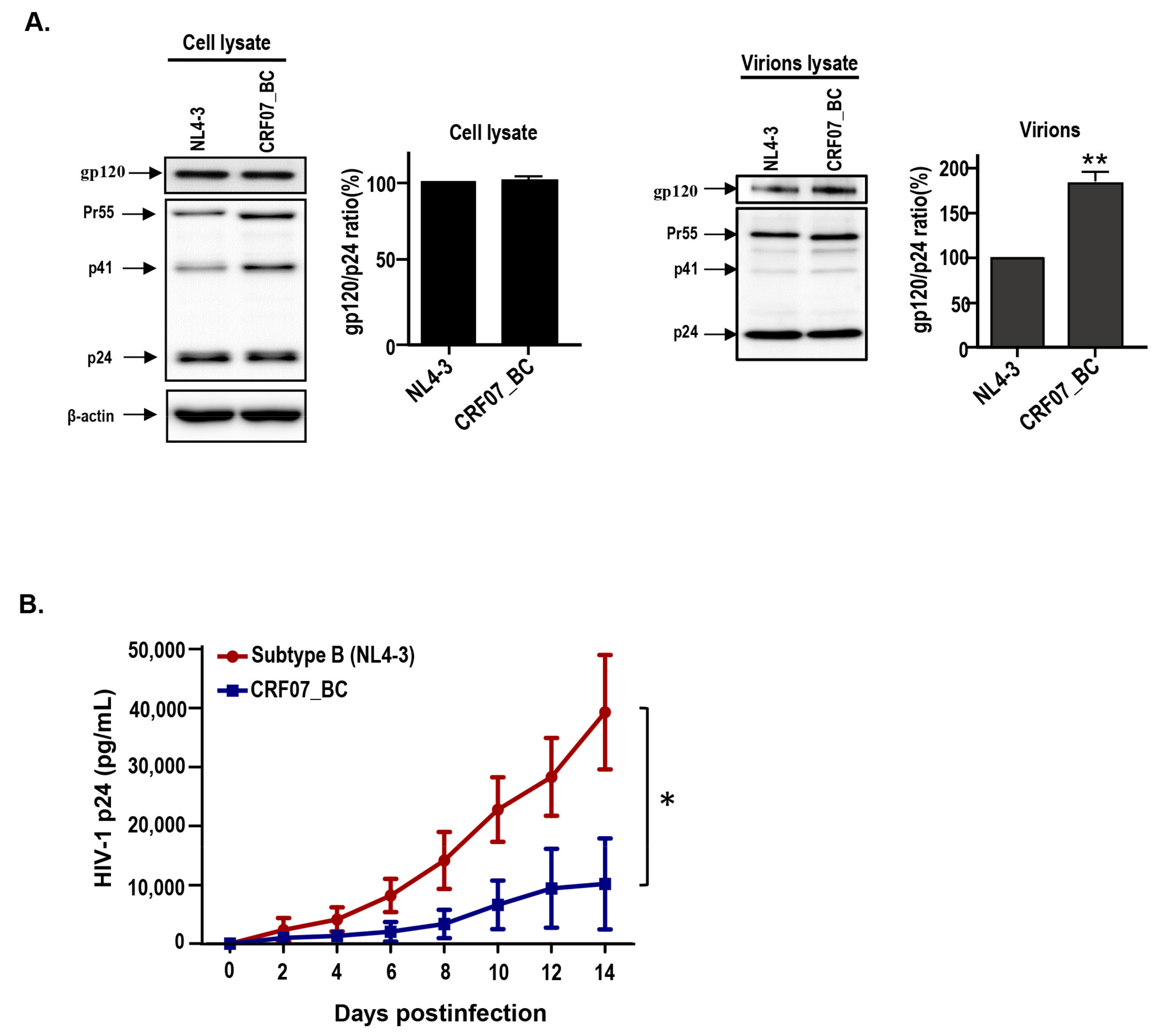
2.2. Viral Characteristics of HIV-1 CRF07_BC
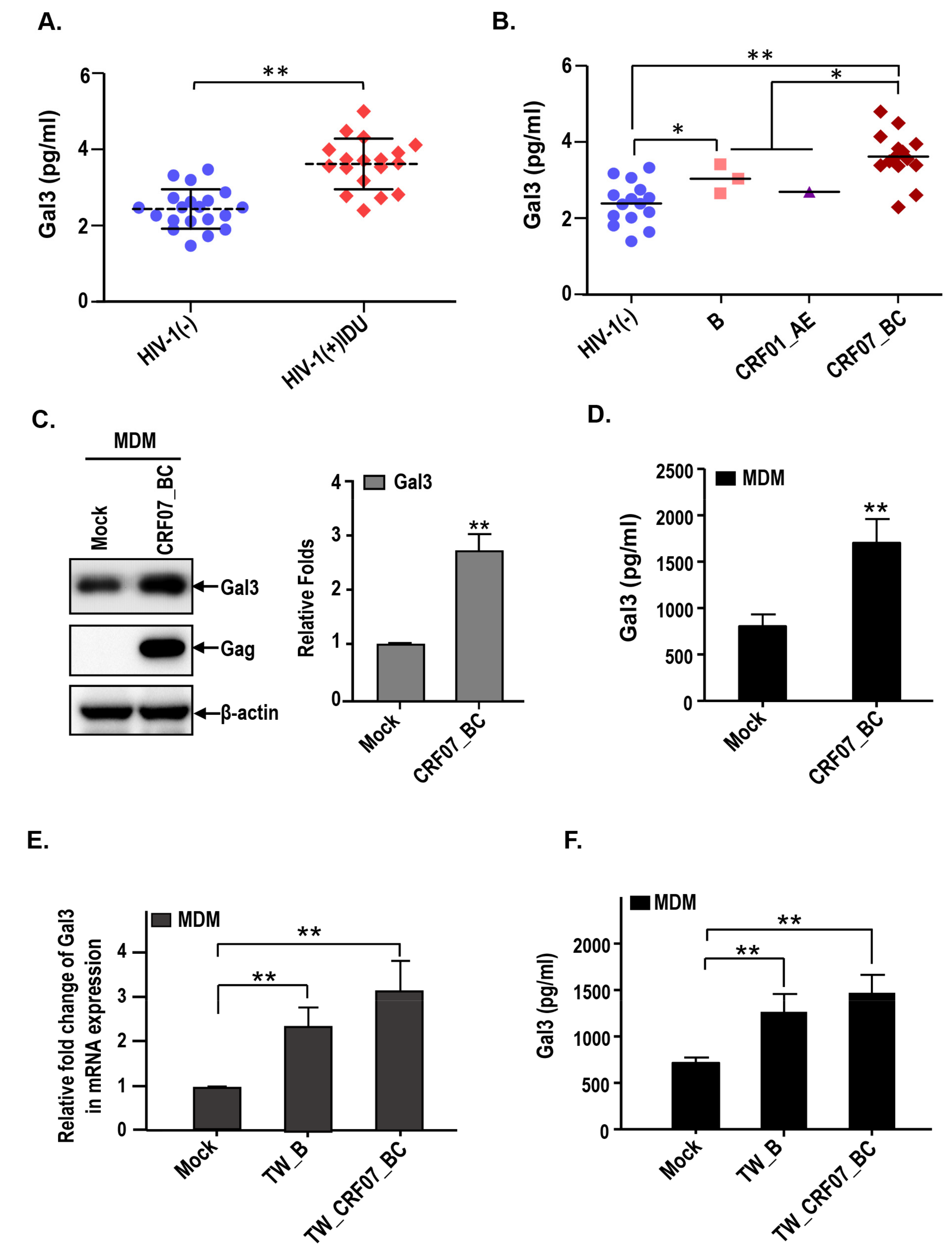
2.3. HIV-1 CRF07_BC Infection Induced Galectin-3 Expression and Secretion
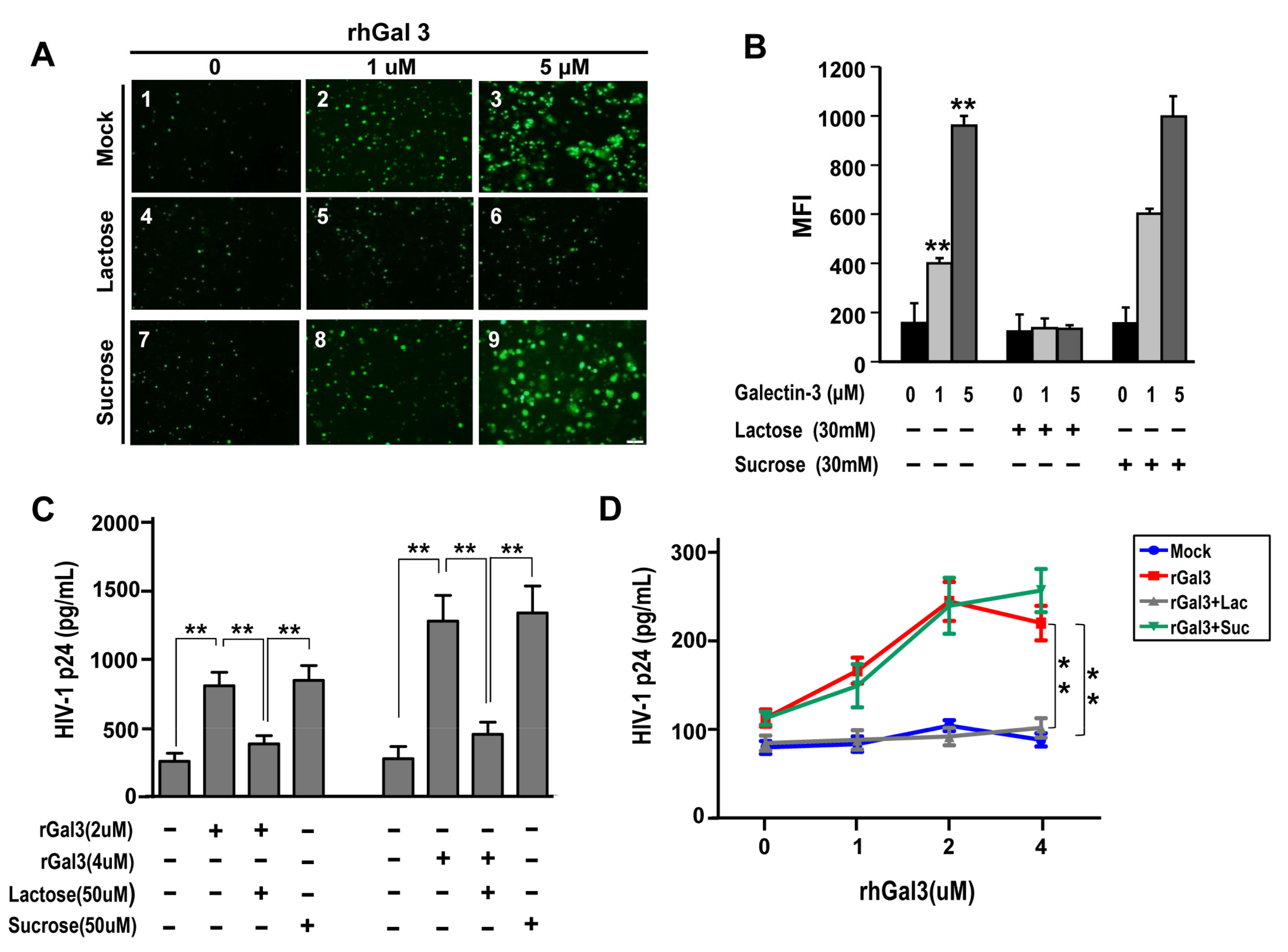
2.4. Galetcin-3 Promotes HIV-1 CRF07_BC Attachment and Internalization
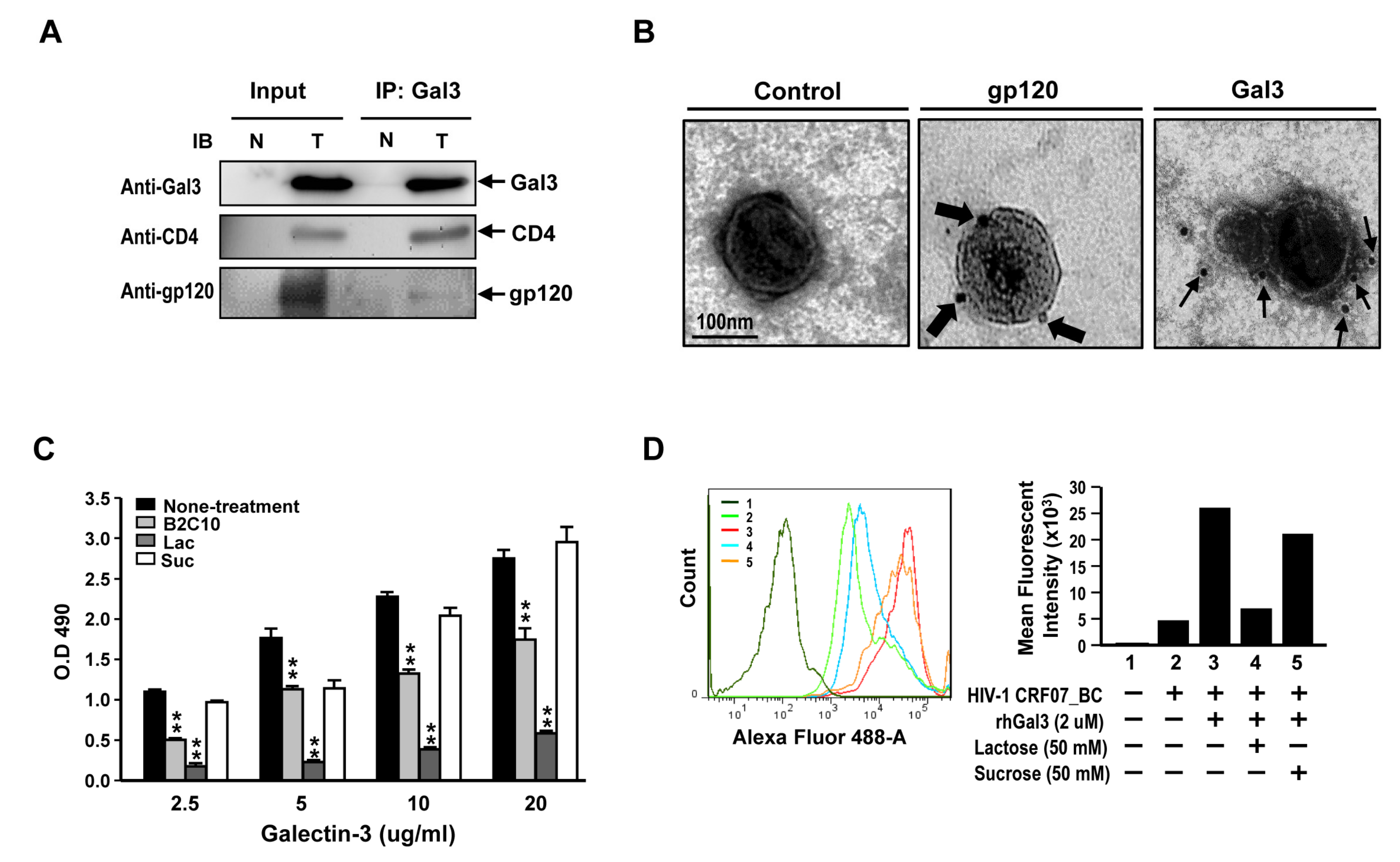
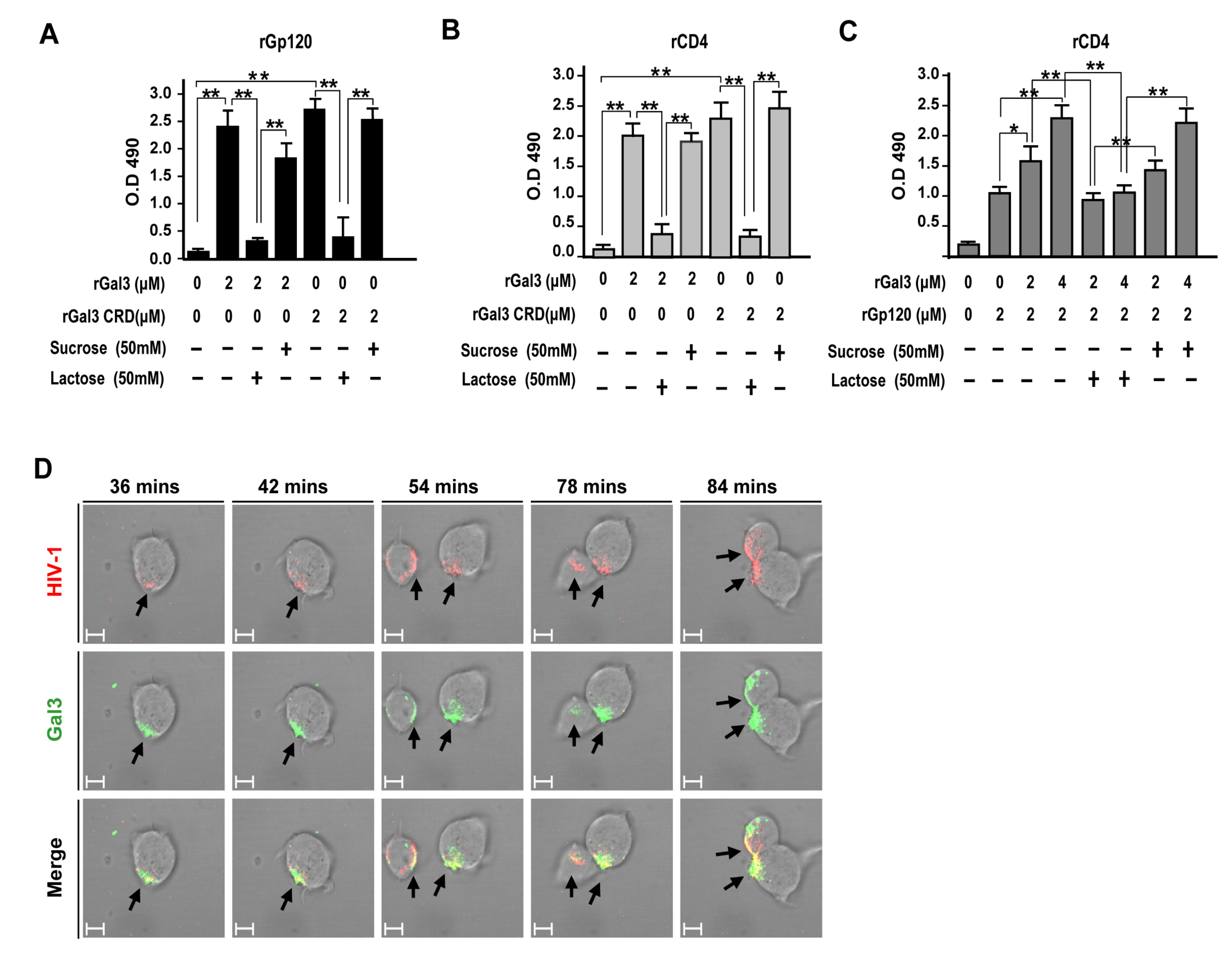
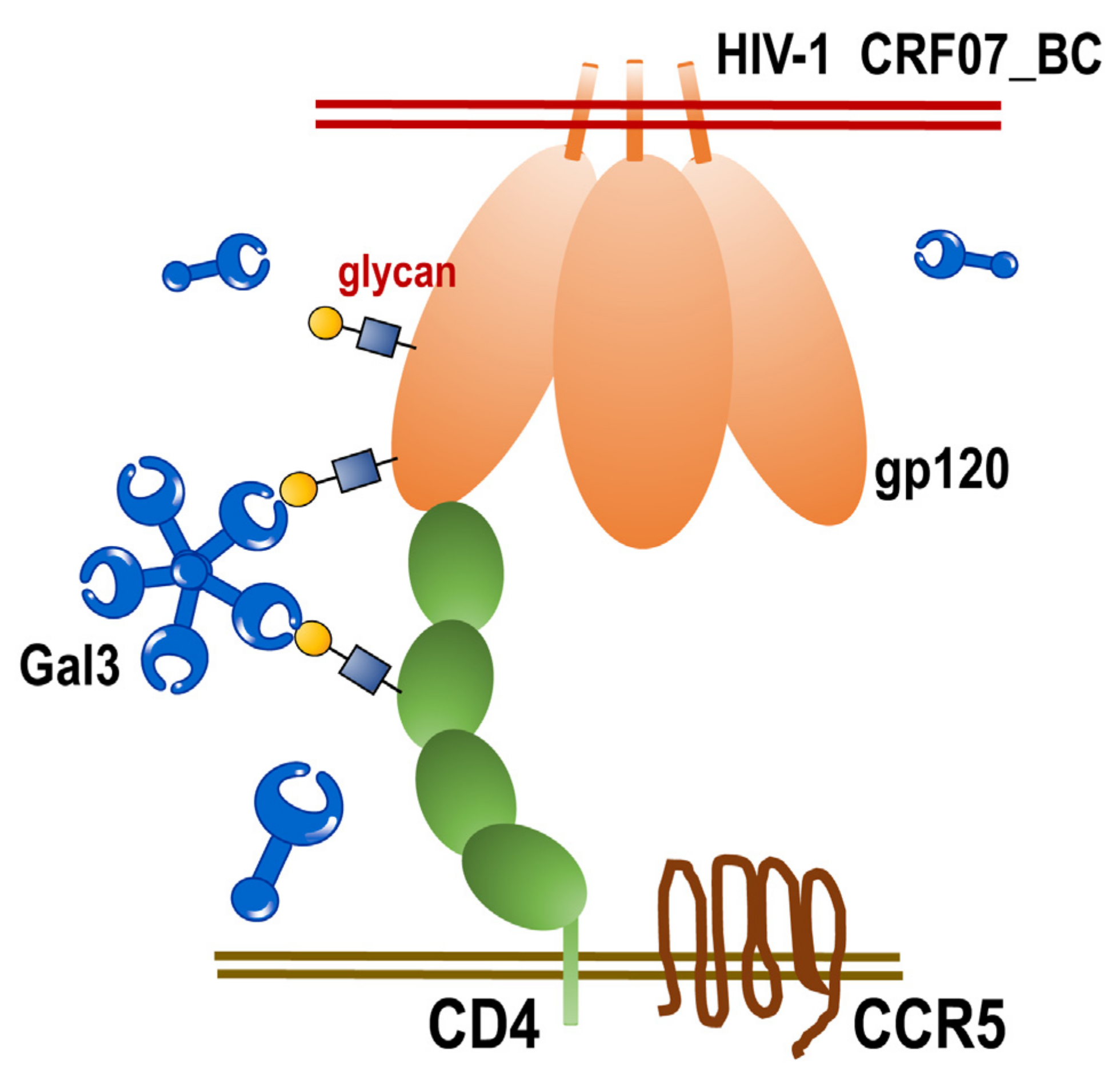
2.5. Galectin-3 Enhanced HIV-1 CRF07_BC Infection by Interacting with Viral Envelope and CD4
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cell Lines and Viruses
4.3. Infectivity, HIV-1 Genotyping Assays
4.4. Viral Growth Kinetic Assay
4.5. Viral Attachment and Internalization Assays
4.6. Enzyme-Linked Immunosorbent Assay (ELISA) and Co-Immunoprecipitation (Co-IP)
4.7. Flow Cytometry and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Electron Microscopy, Life Images and Surface Plasmon Resonance
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Su, L.; Graf, M.; Zhang, Y.; von Briesen, H.; Xing, H.; Kostler, J.; Melzl, H.; Wolf, H.; Shao, Y.; Wagner, R. Characterization of a virtually full-length human immunodeficiency virus type 1 genome of a prevalent intersubtype (C/B’) recombinant strain in China. J. Virol. 2000, 74, 11367–11376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Xia, X.; Kusagawa, S.; Zhang, C.; Ben, K.; Takebe, Y. On-going generation of multiple forms of HIV-1 intersubtype recombinants in the Yunnan Province of China. AIDS 2002, 16, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Pant Pai, N.; Shivkumar, S.; Cajas, J.M. Does genetic diversity of HIV-1 non-B subtypes differentially impact disease progression in treatment-naive HIV-1-infected individuals? A systematic review of evidence: 1996-2010. J. Acquir. Immune. Defic. Syndr. 2012, 59, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.H.; Lai, C.C.; Yang, J.L.; Huang, H.L.; Huang, M.S.; Tsai, M.S.; Yang, C.J.; Cheng, C.L.; Su, Y.C.; Chang, S.F.; et al. Slow immunological progression in HIV-1 CRF07_BC-infected injecting drug users. Emerg. Microbes Infect. 2013, 2, e83. [Google Scholar] [CrossRef]
- Huang, S.W.; Wang, S.F.; Lin, Y.T.; Yen, C.H.; Lee, C.H.; Wong, W.W.; Tsai, H.C.; Yang, C.J.; Hu, B.S.; Lin, Y.H.; et al. Patients infected with CRF07_BC have significantly lower viral loads than patients with HIV-1 subtype B: Mechanism and impact on disease progression. PLoS ONE 2014, 9, e114441. [Google Scholar] [CrossRef]
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. HIV: Cell binding and entry. Cold Spring Harb. Perspect. Med. 2012, 2, a006866. [Google Scholar] [CrossRef]
- Ugolini, S.; Mondor, I.; Sattentau, Q.J. HIV-1 attachment: Another look. Trends Microbiol. 1999, 7, 144–149. [Google Scholar] [CrossRef]
- Hijazi, K.; Wang, Y.; Scala, C.; Jeffs, S.; Longstaff, C.; Stieh, D.; Haggarty, B.; Vanham, G.; Schols, D.; Balzarini, J.; et al. DC-SIGN increases the affinity of HIV-1 envelope glycoprotein interaction with CD4. PLoS ONE 2011, 6, e28307. [Google Scholar] [CrossRef] [Green Version]
- St-Pierre, C.; Manya, H.; Ouellet, M.; Clark, G.F.; Endo, T.; Tremblay, M.J.; Sato, S. Host-soluble galectin-1 promotes HIV-1 replication through a direct interaction with glycans of viral gp120 and host CD4. J. Virol. 2011, 85, 11742–11751. [Google Scholar] [CrossRef] [Green Version]
- Mercier, S.; St-Pierre, C.; Pelletier, I.; Ouellet, M.; Tremblay, M.J.; Sato, S. Galectin-1 promotes HIV-1 infectivity in macrophages through stabilization of viral adsorption. Virology 2008, 371, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, M.; Mercier, S.; Pelletier, I.; Bounou, S.; Roy, J.; Hirabayashi, J.; Sato, S.; Tremblay, M.J. Galectin-1 acts as a soluble host factor that promotes HIV-1 infectivity through stabilization of virus attachment to host cells. J. Immunol. 2005, 174, 4120–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.H.; Lin, C.Y.; Chang, M.R.; Urbina, A.N.; Assavalapsakul, W.; Thitithanyanont, A.; Chen, Y.H.; Liu, F.T.; Wang, S.F. The role of galectins in virus infection-A systemic literature review. J. Microbiol. Immunol. Infect 2019, S1684-1182(19)30149-5. [Google Scholar] [CrossRef] [PubMed]
- Cummings, R.D.; Liu, F.T.; Vasta, G.R. Galectins. In ssentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017; pp. 469–480. [Google Scholar]
- Liu, F.T.; Patterson, R.J.; Wang, J.L. Intracellular functions of galectins. Biochim. Biophys. Acta 2002, 1572, 263–273. [Google Scholar] [CrossRef]
- Yang, R.Y.; Rabinovich, G.A.; Liu, F.T. Galectins: Structure, function and therapeutic potential. Expert Rev. Mol. Med. 2008, 10, e17. [Google Scholar] [CrossRef]
- Vasta, G.R. Roles of galectins in infection. Nat. Rev. Microbiol. 2009, 7, 424–438. [Google Scholar] [CrossRef] [Green Version]
- Ochieng, J.; Furtak, V.; Lukyanov, P. Extracellular functions of galectin-3. Glycoconj. J. 2002, 19, 527–535. [Google Scholar] [CrossRef]
- Woodward, A.M.; Mauris, J.; Argueso, P. Binding of transmembrane mucins to galectin-3 limits herpesvirus 1 infection of human corneal keratinocytes. J. Virol. 2013, 87, 5841–5847. [Google Scholar] [CrossRef] [Green Version]
- Quattroni, P.; Li, Y.; Lucchesi, D.; Lucas, S.; Hood, D.W.; Herrmann, M.; Gabius, H.J.; Tang, C.M.; Exley, R.M. Galectin-3 binds Neisseria meningitidis and increases interaction with phagocytic cells. Cell Microbiol. 2012, 14, 1657–1675. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Zhou, X.; Yuan, D.; Yang, H.; Wei, D.; Qin, G.; Liang, S. Prevalence and patterns of drug-resistance mutations among HIV-1 patients infected with CRF07_BC strains in Sichuan province, China. Virol. Sin. 2014, 29, 237–241. [Google Scholar] [CrossRef]
- Elahi, S.; Niki, T.; Hirashima, M.; Horton, H. Galectin-9 binding to Tim-3 renders activated human CD4+ T cells less susceptible to HIV-1 infection. Blood 2012, 119, 4192–4204. [Google Scholar] [CrossRef]
- Wang, S.F.; Tsao, C.H.; Lin, Y.T.; Hsu, D.K.; Chiang, M.L.; Lo, C.H.; Chien, F.C.; Chen, P.; Arthur Chen, Y.M.; Chen, H.Y.; et al. Galectin-3 promotes HIV-1 budding via association with Alix and Gag p6. Glycobiology 2014, 24, 1022–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.T.; Bevins, C.L. A sweet target for innate immunity. Nat. Med. 2010, 16, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Y.; Chen, M.; Huang, S.W.; Jen, I.A.; Wang, S.F.; Yang, J.Y.; Chen, Y.H.; Chen, Y.A. Molecular epidemiology of HIV-1 infection among men who have sex with men in Taiwan from 2013 to 2015. PLoS ONE 2018, 13, e0202622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.C.; Elbeik, T.; Dileanis, J.; Ng, V.; Chen, Y.J.; Leu, H.S.; Cheng, S.H.; Wong, J.C.; Wong, W.W.; Chen, Y.M. Molecular epidemiology of HIV-1 subtypes and drug resistant strains in Taiwan. J. Med. Virol. 2008, 80, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Lan, Y.C.; Lai, S.F.; Yang, J.Y.; Tsai, S.F.; Kuo, S.H. HIV-1 CRF07_BC infections, injecting drug users, Taiwan. Emerg. Infect. Dis. 2006, 12, 703–705. [Google Scholar] [CrossRef]
- Taylor, M.E.; Drickamer, K. Structure-function analysis of C-type animal lectins. Methods Enzymol. 2003, 363, 3–16. [Google Scholar]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Sharma, B.B.; Yu, L.; Zuberi, R.; Weng, I.C.; Kawakami, Y.; Kawakami, T.; Hsu, D.K.; Liu, F.T. Role of galectin-3 in mast cell functions: Galectin-3-deficient mast cells exhibit impaired mediator release and defective JNK expression. J. Immunol. 2006, 177, 4991–4997. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.Y.; Hsu, D.K.; Liu, F.T. Expression of galectin-3 modulates T-cell growth and apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 6737–6742. [Google Scholar] [CrossRef] [Green Version]
- Fogel, S.; Guittaut, M.; Legrand, A.; Monsigny, M.; Hebert, E. The tat protein of HIV-1 induces galectin-3 expression. Glycobiology 1999, 9, 383–387. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Chertova, E.; Bess, J., Jr.; Lifson, J.D.; Arthur, L.O.; Liu, J.; Taylor, K.A.; Roux, K.H. Electron tomography analysis of envelope glycoprotein trimers on HIV and simian immunodeficiency virus virions. Proc. Natl. Acad. Sci. USA 2003, 100, 15812–15817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checkley, M.A.; Luttge, B.G.; Freed, E.O. HIV-1 envelope glycoprotein biosynthesis, trafficking, and incorporation. J. Mol. Biol. 2011, 410, 582–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.H.; Yeh, C.S.; Lin, C.Y.; Yuan, R.Y.; Urbina, A.N.; Lu, P.L.; Chen, Y.H.; Chen, Y.A.; Liu, F.T.; Wang, S.F. Amino Acid Deletions in p6(Gag) Domain of HIV-1 CRF07_BC Ameliorate Galectin-3 Mediated Enhancement in Viral Budding. Int. J. Mol. Sci. 2020, 21, 2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepur, A.; Salomonsson, E.; Nilsson, U.J.; Leffler, H. Ligand induced galectin-3 protein self-association. J. Biol. Chem. 2012, 287, 21751–21756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janas, A.M.; Wu, L. HIV-1 interactions with cells: From viral binding to cell-cell transmission. Curr. Protoc. Cell Biol. 2009, 43, 26.5.1–26.5.20. [Google Scholar]
- Wei, M.; Guan, Q.; Liang, H.; Chen, J.; Chen, Z.; Hei, F.; Feng, Y.; Hong, K.; Huang, H.; Xing, H.; et al. Simple subtyping assay for human immunodeficiency virus type 1 subtypes B, C, CRF01-AE, CRF07-BC, and CRF08-BC. J. Clin. Microbiol. 2004, 42, 4261–4267. [Google Scholar] [CrossRef] [Green Version]
- Tai, C.J.; Li, C.L.; Tai, C.J.; Wang, C.K.; Lin, L.T. Early Viral Entry Assays for the Identification and Evaluation of Antiviral Compounds. J. Vis. Exp. 2015, 105, e53124. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, Y.; Jia, W.; Kidoya, H.; Muramatsu, F.; Tsukada, Y.; Takakura, N. Galectin-3 Inhibits Cancer Metastasis by Negatively Regulating Integrin beta3 Expression. Am. J. Pathol. 2019, 189, 900–910. [Google Scholar] [CrossRef]
- Li, Y.C.; Chang, Y.F.; Su, L.C.; Chou, C. Differential-phase surface plasmon resonance biosensor. Anal. Chem. 2008, 80, 5590–5595. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HIV-1(+) Injecting Drug Users (IDUs) | |||
|---|---|---|---|
| Characteristics | Male (%) (n = 26) | Female (%) (n = 2) | Total (%) (n = 28) |
| Age (yrs.) | |||
| 15–29 | 8 (30.5) | 1 (50) | 8 (28.6) |
| 30–49 | 16 (61.5) | 1 (50) | 19 (67.9) |
| ≥50 | 2(7.6) | 0 (0) | 1 (3.5) |
| CD4 Count (cells/mm3) | |||
| <200 | 1(3.8) | 0(0) | 1(3.5) |
| 200–500 | 20(76.9) | 2(100) | 22(78.6) |
| >500 | 5(19.2) | 0(0) | 5(17.9) |
| Viral Load (copies/mL) | |||
| <5000 | 1 (3.8) | 0(0) | 1(3.5) |
| 5000–10,000 | 4(15.4) | 1(50) | 5 (17.9) |
| 10,000–100,000 | 19(73.1) | 1(50) | 20(71.4) |
| >100,000 | 2(7.6) | 0(0) | 2(7.1) |
| HIV-1 genotype | |||
| B | 3(11.5) | 0(0) | 3(10.7) |
| CRF01_AE | 0(0) | 1(50) | 1(3.5) |
| CRF07_BC | 23(88.5) | 1(50) | 24(85.7) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.-Y.; Wang, W.-H.; Huang, S.-W.; Yeh, C.-S.; Yuan, R.-Y.; Yang, Z.-S.; Urbina, A.N.; Tseng, S.-P.; Lu, P.-L.; Chen, Y.-H.; et al. The Examination of Viral Characteristics of HIV-1 CRF07_BC and Its Potential Interaction with Extracellular Galectin-3. Pathogens 2020, 9, 425. https://doi.org/10.3390/pathogens9060425
Lin C-Y, Wang W-H, Huang S-W, Yeh C-S, Yuan R-Y, Yang Z-S, Urbina AN, Tseng S-P, Lu P-L, Chen Y-H, et al. The Examination of Viral Characteristics of HIV-1 CRF07_BC and Its Potential Interaction with Extracellular Galectin-3. Pathogens. 2020; 9(6):425. https://doi.org/10.3390/pathogens9060425
Chicago/Turabian StyleLin, Chih-Yen, Wen-Hung Wang, Szu-Wei Huang, Chun-Sheng Yeh, Ruei-Yu Yuan, Zih-Syuan Yang, Aspiro Nayim Urbina, Sung-Pin Tseng, Po-Liang Lu, Yen-Hsu Chen, and et al. 2020. "The Examination of Viral Characteristics of HIV-1 CRF07_BC and Its Potential Interaction with Extracellular Galectin-3" Pathogens 9, no. 6: 425. https://doi.org/10.3390/pathogens9060425
APA StyleLin, C. -Y., Wang, W. -H., Huang, S. -W., Yeh, C. -S., Yuan, R. -Y., Yang, Z. -S., Urbina, A. N., Tseng, S. -P., Lu, P. -L., Chen, Y. -H., & Wang, S. -F. (2020). The Examination of Viral Characteristics of HIV-1 CRF07_BC and Its Potential Interaction with Extracellular Galectin-3. Pathogens, 9(6), 425. https://doi.org/10.3390/pathogens9060425