43-Year Temporal Trends in Immune Response to Oral Bacteria in a Swedish Population



Abstract
:1. Introduction
2. Results
2.1. Study Group Characteristics
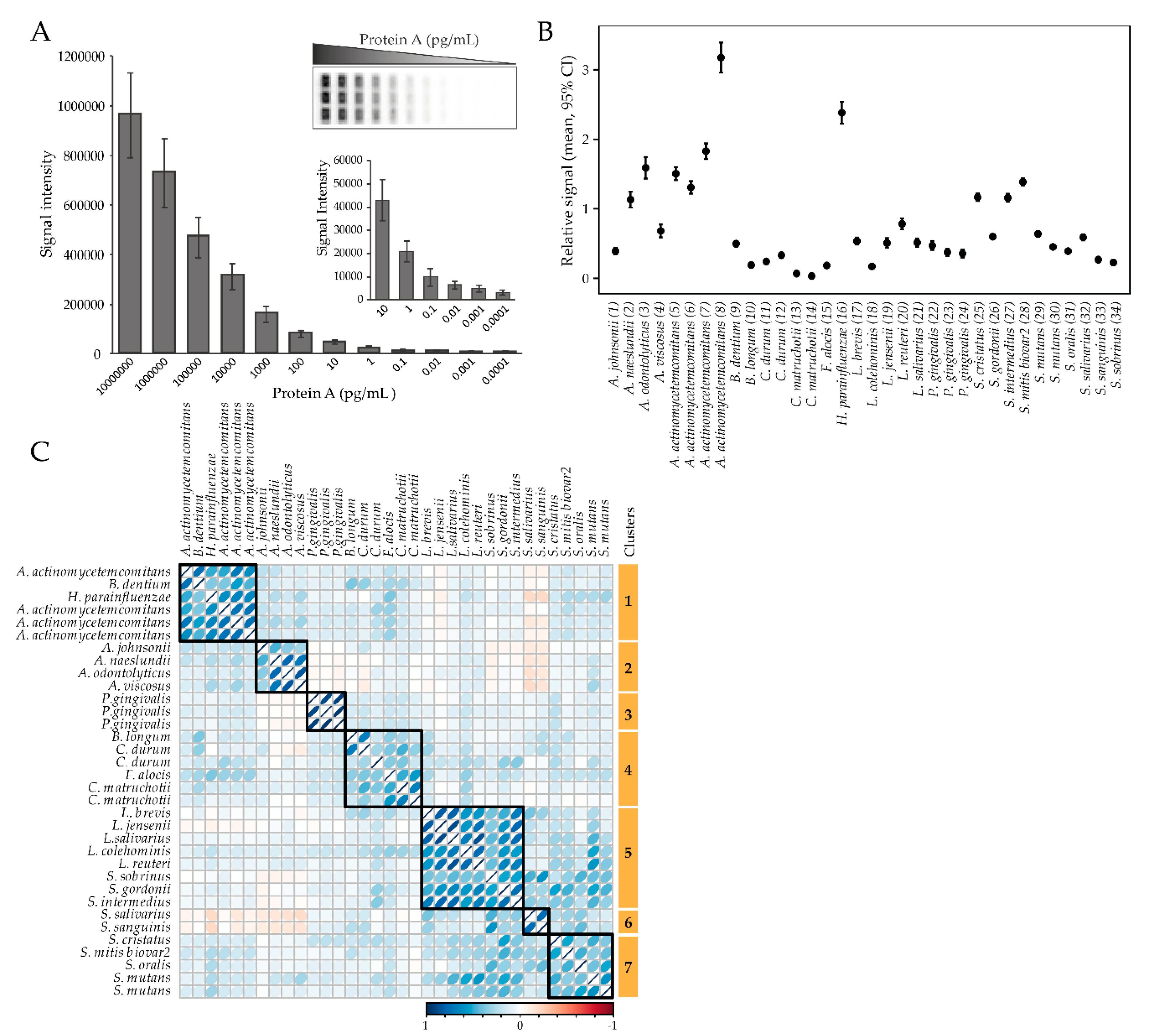
2.2. Overall Plasma Antibody Response Profiles for Oral Bacteria
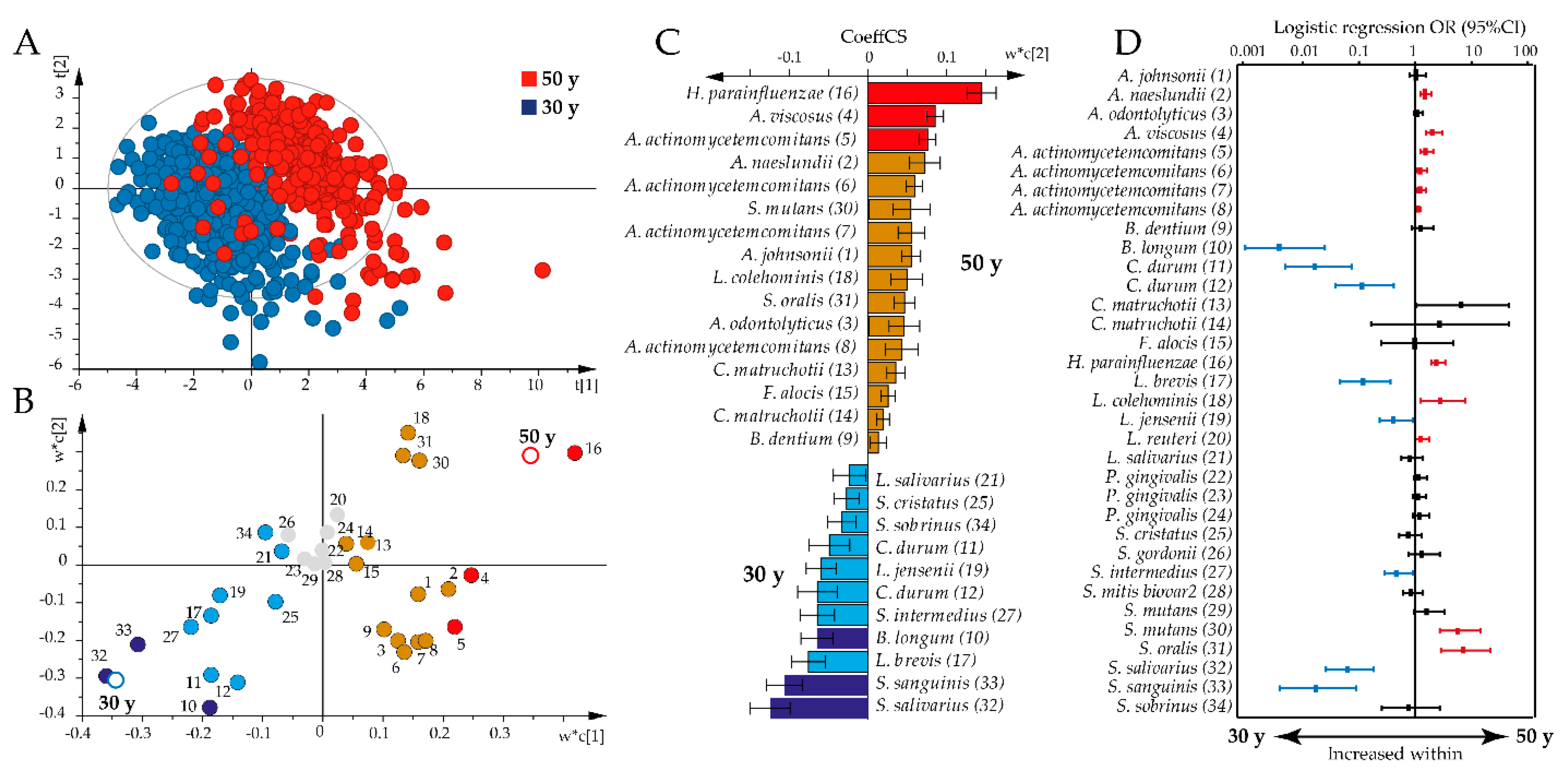
2.3. Age-Related Shifts in Overall Plasma Antibody Profiles
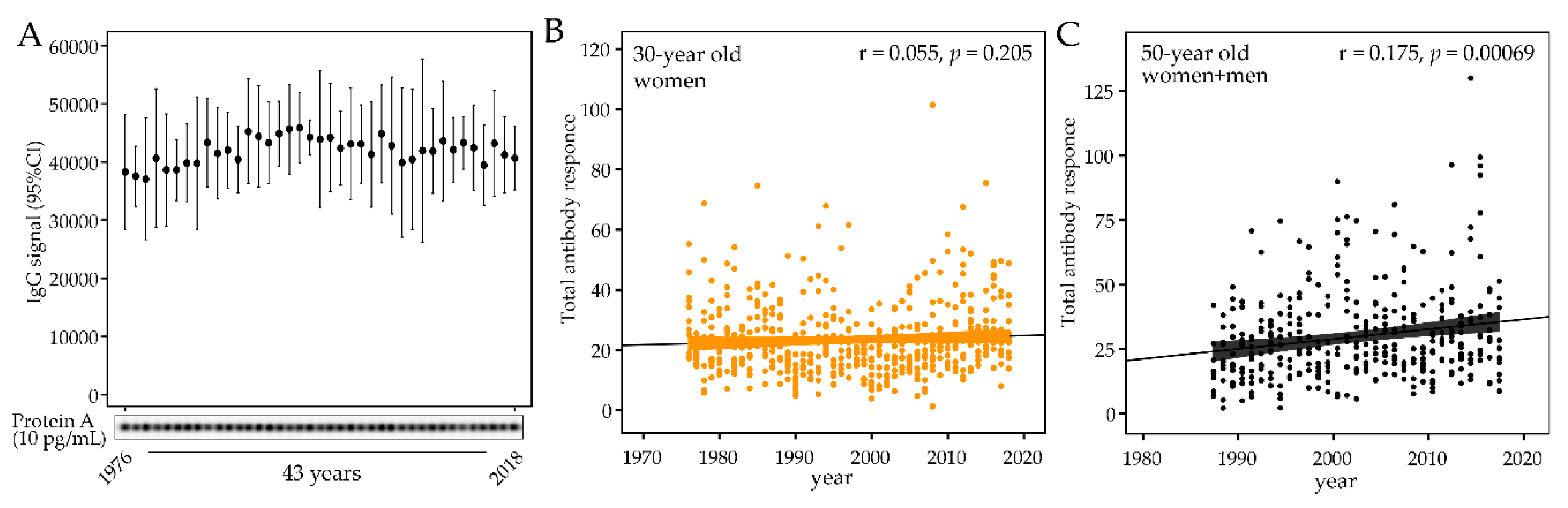
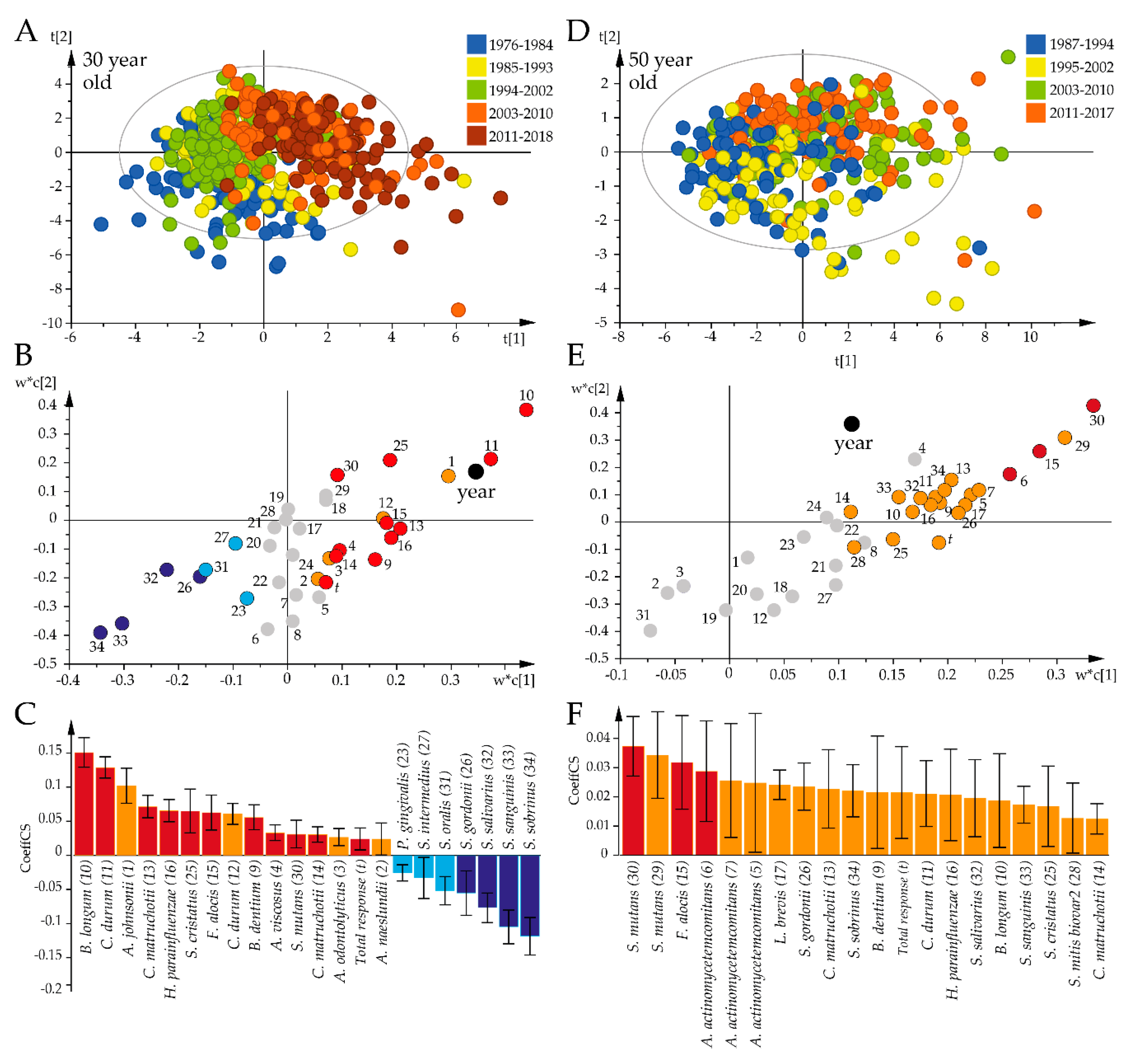
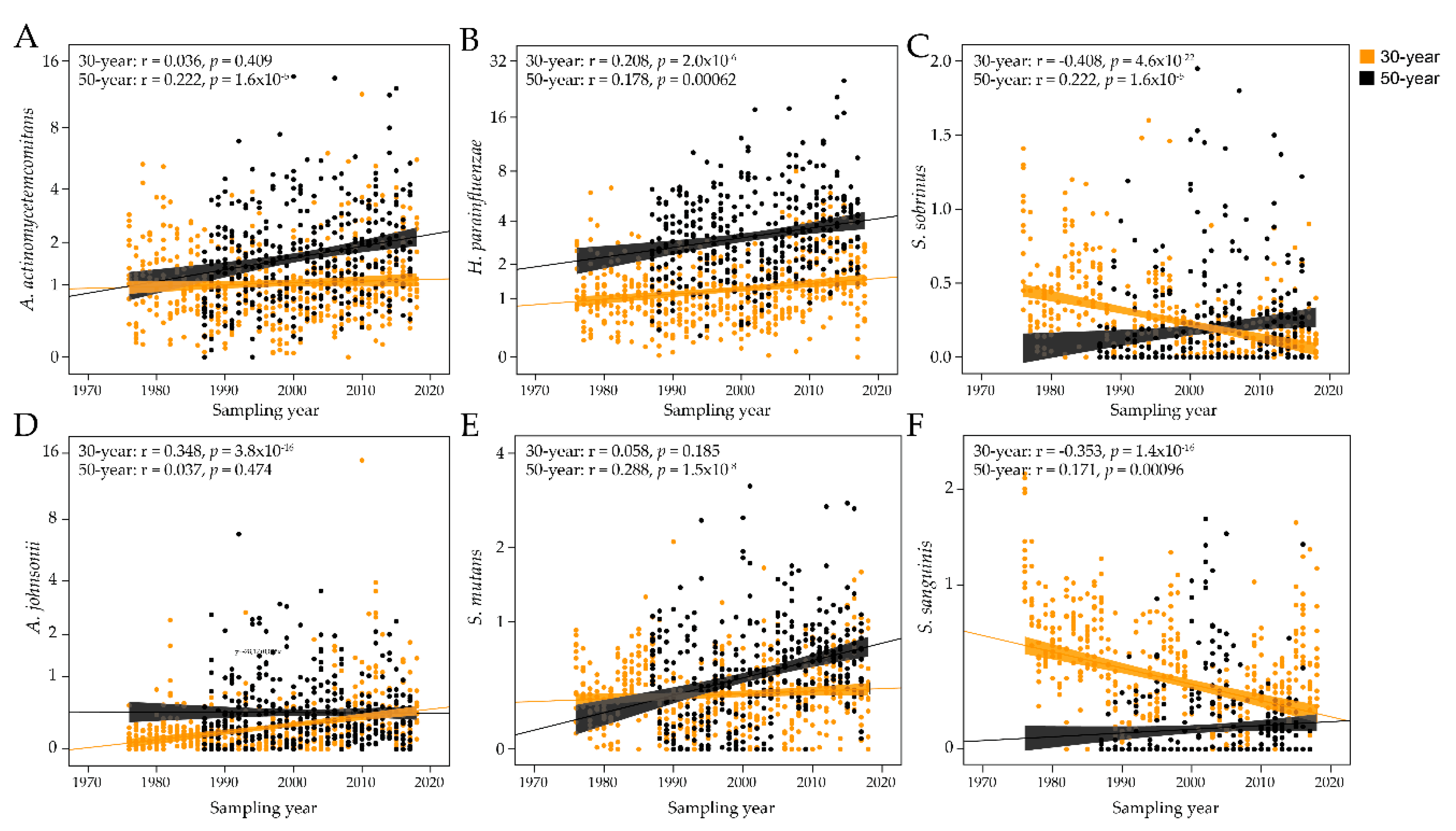
2.4. Forty-Three-Year Temporal Shifts in the Global Plasma Antibody Response to Oral Bacteria
2.5. Species-Specific Antibody Shifts over Time
3. Discussion
4. Materials and Methods
4.1. Study Group
4.2. Bacteria and Culturing
4.3. Antibody Detection
4.4. Estimation of IgG Storage Stability
4.5. Data Handling and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Barraza, J.P.; Arthur, R.A.; Hara, A.; Lewis, K.; Liu, Y.; Scisci, E.L.; Hajishengallis, E.; Whiteley, M.; Koo, H. Spatial mapping of polymicrobial communities reveals a precise biogeography associated with human dental caries. Proc. Natl. Acad. Sci. USA 2020, 117, 12375–12386. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Beighton, D.; Curtis, M.A.; Cury, J.A.; Dige, I.; Dommisch, H.; Ellwood, R.; Giacaman, R.A.; Herrera, D.; Herzberg, M.C.; et al. Role of microbial biofilms in the maintenance of oral health and in the development of dental caries and periodontal diseases. Consensus report of group 1 of the Joint EFP/ORCA workshop on the boundaries between caries and periodontal disease. J. Clin. Periodontol. 2017, 44 (Suppl. 18), S5–S11. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.A.; Diaz, P.I.; Van Dyke, T.E. The role of the microbiota in periodontal disease. Periodontology 2000 2020, 83, 14–25. [Google Scholar] [CrossRef]
- Deo, P.N.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar] [CrossRef]
- Dahlen, G.; Basic, A.; Bylund, J. Importance of Virulence Factors for the Persistence of Oral Bacteria in the Inflamed Gingival Crevice and in the Pathogenesis of Periodontal Disease. J. Clin. Med. 2019, 8, 1339. [Google Scholar] [CrossRef] [Green Version]
- Esberg, A.; Sheng, N.; Marell, L.; Claesson, R.; Persson, K.; Boren, T.; Stromberg, N. Streptococcus Mutans Adhesin Biotypes that Match and Predict Individual Caries Development. EBioMedicine 2017, 24, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Belibasakis, G.N.; Maula, T.; Bao, K.; Lindholm, M.; Bostanci, N.; Oscarsson, J.; Ihalin, R.; Johansson, A. Virulence and Pathogenicity Properties of Aggregatibacter actinomycetemcomitans. Pathogens 2019, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef]
- Fine, D.H.; Markowitz, K.; Fairlie, K.; Tischio-Bereski, D.; Ferrendiz, J.; Furgang, D.; Paster, B.J.; Dewhirst, F.E. A consortium of Aggregatibacter actinomycetemcomitans, Streptococcus parasanguinis, and Filifactor alocis is present in sites prior to bone loss in a longitudinal study of localized aggressive periodontitis. J. Clin. Microbiol. 2013, 51, 2850–2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akrivopoulou, C.; Green, I.M.; Donos, N.; Nair, S.P.; Ready, D. Aggregatibacter actinomycetemcomitans serotype prevalence and antibiotic resistance in a UK population with periodontitis. J. Glob. Antimicrob. Resist. 2017, 10, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Tjokro, N.O.; Kittichotirat, W.; Torittu, A.; Ihalin, R.; Bumgarner, R.E.; Chen, C. Transcriptomic Analysis of Aggregatibacter actinomycetemcomitans Core and Accessory Genes in Different Growth Conditions. Pathogens 2019, 8, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebersole, J.L.; Dawson, D.R., 3rd; Morford, L.A.; Peyyala, R.; Miller, C.S.; Gonzalez, O.A. Periodontal disease immunology: ‘Double indemnity’ in protecting the host. Periodontol 2000 2013, 62, 163–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebersole, J.L.; Dawson, D.A., 3rd; Emecen Huja, P.; Pandruvada, S.; Basu, A.; Nguyen, L.; Zhang, Y.; Gonzalez, O.A. Age and Periodontal Health - Immunological View. Curr. Oral Health Rep. 2018, 5, 229–241. [Google Scholar] [CrossRef]
- Johansson, A.; Eriksson, M.; Ahren, A.M.; Boman, K.; Jansson, J.H.; Hallmans, G.; Johansson, I. Prevalence of systemic immunoreactivity to Aggregatibacter actinomycetemcomitans leukotoxin in relation to the incidence of myocardial infarction. BMC Infect. Dis. 2011, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Michaud, D.S.; Izard, J.; Rubin, Z.; Johansson, I.; Weiderpass, E.; Tjonneland, A.; Olsen, A.; Overvad, K.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; et al. Lifestyle, dietary factors, and antibody levels to oral bacteria in cancer-free participants of a European cohort study. Cancer Causes Control 2013, 24, 1901–1909. [Google Scholar] [CrossRef] [Green Version]
- Lif Holgerson, P.; Esberg, A.; Sjödin, A.; West, C.E.; Johansson, I. Salivary microbiota transformation from 2 days to 18 years of age using 16S ribosomal RNA gene sequencing. Sci. Rep. 2020. accepted. [Google Scholar]
- Eriksson, L.; Lif Holgerson, P.; Johansson, I. Saliva and tooth biofilm bacterial microbiota in adolescents in a low caries community. Sci. Rep. 2017, 7, 5861. [Google Scholar] [CrossRef] [Green Version]
- Johansson, I.; Witkowska, E.; Kaveh, B.; Lif Holgerson, P.; Tanner, A.C. The Microbiome in Populations with a Low and High Prevalence of Caries. J. Dent. Res. 2016, 95, 80–86. [Google Scholar] [CrossRef]
- Norberg, M.; Wall, S.; Boman, K.; Weinehall, L. The Vasterbotten Intervention Programme: Background, design and implications. Glob. Health Act. 2010, 3. [Google Scholar] [CrossRef]
- Murra, M.; Lutzen, L.; Barut, A.; Zbinden, R.; Lund, M.; Villesen, P.; Norskov-Lauritsen, N. Whole-Genome Sequencing of Aggregatibacter Species Isolated from Human Clinical Specimens and Description of Aggregatibacter kilianii sp. nov. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [Green Version]
- Aberg, C.H.; Kelk, P.; Johansson, A. Aggregatibacter actinomycetemcomitans: Virulence of its leukotoxin and association with aggressive periodontitis. Virulence 2015, 6, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, A.P.; Boches, S.K.; Cotton, S.L.; Goodson, J.M.; Kent, R.; Haffajee, A.D.; Socransky, S.S.; Hasturk, H.; Van Dyke, T.E.; Dewhirst, F.; et al. Comparisons of subgingival microbial profiles of refractory periodontitis, severe periodontitis, and periodontal health using the human oral microbe identification microarray. J. Periodontol. 2009, 80, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Norskov-Lauritsen, N. Classification, identification, and clinical significance of Haemophilus and Aggregatibacter species with host specificity for humans. Clin. Microbiol. Rev. 2014, 27, 214–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiranmayi, K.V.; Sirisha, K.; Ramoji Rao, M.V.; Sudhakar, P. Novel Pathogens in Periodontal Microbiology. J. Pharm. Bioallied Sci. 2017, 9, 155–163. [Google Scholar] [CrossRef]
- Scharnow, A.M.; Solinski, A.E.; Wuest, W.M. Targeting S. mutans biofilms: A perspective on preventing dental caries. MedChemComm 2019, 10, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Moossavi, S.; Bishehsari, F. Microbes: Possible link between modern lifestyle transition and the rise of metabolic syndrome. Obes. Rev. 2019, 20, 407–419. [Google Scholar] [CrossRef]
- Willis, J.R.; Gabaldon, T. The Human Oral Microbiome in Health and Disease: From Sequences to Ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Kononen, E.; Gursoy, M.; Gursoy, U.K. Periodontitis: A Multifaceted Disease of Tooth-Supporting Tissues. J. Clin. Med. 2019, 8, 1135. [Google Scholar] [CrossRef] [Green Version]
- Oscarsson, J.; Claesson, R.; Lindholm, M.; Hoglund Aberg, C.; Johansson, A. Tools of Aggregatibacter actinomycetemcomitans to Evade the Host Response. J. Clin. Med. 2019, 8, 1079. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Lee, A.; Santin, E. Microbiome and pathogen interaction with the immune system. Poult. Sci. 2020, 99, 1906–1913. [Google Scholar] [CrossRef]
- Koch, G.; Helkimo, A.N.; Ullbro, C. Caries prevalence and distribution in individuals aged 3-20 years in Jonkoping, Sweden: Trends over 40 years. Eur. Arch Paediatr. Dent. 2017, 18, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Norderyd, O.; Koch, G.; Papias, A.; Kohler, A.A.; Helkimo, A.N.; Brahm, C.O.; Lindmark, U.; Lindfors, N.; Mattsson, A.; Rolander, B.; et al. Oral health of individuals aged 3–80 years in Jonkoping, Sweden during 40 years (1973–2013). II. Review of clinical and radiographic findings. Swed. Dent. J. 2015, 39, 69–86. [Google Scholar]
- Kilian, M.; Chapple, I.L.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome—An update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Lira-Junior, R.; Akerman, S.; Klinge, B.; Bostrom, E.A.; Gustafsson, A. Salivary microbial profiles in relation to age, periodontal, and systemic diseases. PLoS ONE 2018, 13, e0189374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinehall, L.; Hallgren, C.G.; Westman, G.; Janlert, U.; Wall, S. Reduction of selection bias in primary prevention of cardiovascular disease through involvement of primary health care. Scand. J. Prim. Health Care 1998, 16, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Hallmans, G.; Agren, A.; Johansson, G.; Johansson, A.; Stegmayr, B.; Jansson, J.H.; Lindahl, B.; Rolandsson, O.; Soderberg, S.; Nilsson, M.; et al. Cardiovascular disease and diabetes in the Northern Sweden Health and Disease Study Cohort—Evaluation of risk factors and their interactions. Scand. J. Public Health Suppl. 2003, 61, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakellari, D.; Socransky, S.S.; Dibart, S.; Eftimiadi, C.; Taubman, M.A. Estimation of serum antibody to subgingival species using checkerboard immunoblotting. Oral Microbiol. Immunol. 1997, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). 2017. Available online: https://github.com/taiyun/corrplot (accessed on 9 February 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VIP Cohort | Present Study Group | ||||
|---|---|---|---|---|---|
| n = 110,663 | n = 372 | Temporal Trend | |||
| Measure | Trend | Measure | Trend | p-Value | |
| Sample years | 1990–2017 | - | 1987–2017 | - | - |
| Proportion women | 50.7% | - | 50.0% | - | - |
| University level education | 30.4% | ↑ | 30.6% | ↑ | 0.03 |
| BMI a, kg/m2 | 26.6 (26.5, 26.6) | ↑ | 26.1 (25.7, 26.4) | ↑ | 0.018 |
| Fasting blood glucose a, mol/L | 5.52 (5.51, 5.53) | → | 5.29 (5.21, 5.37) | → | 0.149 |
| Never-smoker | 39.2% to 50.7% | ↑ | 41.7% to 71.4% | ↑ | 0.004 |
| Nutrient intake a | |||||
| Total fat, E% | 35.8 (35.7, 35.9) | ↑ | 35.4 (34.7, 36.1) | ↑ | <0.001 |
| Saturated fat, E% | 14.9 (14.9, 15.0) | ↑ | 14.8 (14.3, 15.2) | ↑ | <0.001 |
| Carbohydrates, E% | 46.5 (46.4, 46.5) | ↓ | 46.6 (45.9, 47.4) | ↓ | <0.001 |
| Sucrose, E% | 5.84 (5.82, 5.85) | ↓ | 5.77 (5.50, 6.05) | ↓ | <0.001 |
| Protein, E% | 15.1 (15.1, 15.1) | → | 14.8 (14.6, 15.0) | → | 0.044 |
| Vitamin C, mg/day | 69.9 (69.6, 70.1) | ↓ | 72.8 (68.2, 77.5) | ↓ | 0.017 |
| Vitamin D, mg/day | 5.22 (5.21, 5.23) | → | 5.36 (5.15, 5.57) | → | 0.152 |
| Vitamin E, mg/day | 6.07 (6.06, 6.08) | ↓ | 6.08 (5.92, 6.24) | → | 0.799 |
| Sample Collections | ||||||
|---|---|---|---|---|---|---|
| All Samples (n = 888) | 30-Year-Olds (n = 516) | 50-Year-Olds (n = 372) | ||||
| Species (No.) | β-Value | p-Value | β-Value | p-Value | β-Value | p-Value |
| A. johnsonii (1) | 0.009 | <0.0005 | 0.013 | <0.0005 | −0.002 | 0.518 |
| A. naeslundii (2) | 0.003 | 0.477 | 0.006 | 0.097 | −0.004 | 0.792 |
| A. odontolyticus (3) | 0.011 | 0.117 | 0.015 | 0.009 | −0.002 | 0.933 |
| A. viscosus (4) | 0.014 | <0.0005 | 0.007 | 0.001 | 0.033 | 0.013 |
| A. actinomycetemcomitans (5) | 0.014 | <0.0005 | 0.006 | 0.159 | 0.036 | <0.0005 |
| A. actinomycetemcomitans (6) | 0.005 | 0.272 | −0.007 | 0.231 | 0.038 | <0.0005 |
| A. actinomycetemcomitans (7) | 0.015 | 0.009 | 0.004 | 0.549 | 0.046 | <0.0005 |
| A. actinomycetemcomitans a (8) | 0.012 | 0.205 | 0.000 | 0.990 | 0.045 | 0.024 |
| B. dentium (9) | 0.007 | <0.0005 | 0.006 | 0.001 | 0.009 | 0.029 |
| B. longum (10) | 0.011 | <0.0005 | 0.014 | <0.0005 | 0.002 | 0.010 |
| C. durum (11) | 0.011 | <0.0005 | 0.014 | <0.0005 | 0.003 | 0.001 |
| C. durum (12) | 0.004 | <0.0005 | 0.005 | <0.0005 | 0.001 | 0.548 |
| C. matruchotii (13) | 0.002 | <0.0005 | 0.001 | < 0.0005 | 0.003 | <0.0005 |
| C. matruchotii (14) | 0.001 | 0.006 | 0.000 | 0.107 | 0.002 | 0.021 |
| F. alocis a (15) | 0.003 | <0.0005 | 0.002 | < 0.0005 | 0.006 | <0.0005 |
| H. parainfluenzae (16) | 0.029 | <0.0005 | 0.016 | <0.0005 | 0.066 | <0.0005 |
| L. brevis (17) | 0.001 | 0.586 | 0.000 | 0.982 | 0.005 | 0.010 |
| L. colehominis (18) | 0.001 | 0.245 | 0.001 | 0.283 | 0.001 | 0.582 |
| L. jensenii (19) | −0.001 | 0.557 | −0.002 | 0.584 | −0.001 | 0.829 |
| L. reuteri (20) | −0.002 | 0.404 | −0.004 | 0.089 | 0.003 | 0.744 |
| L. salivarius (21) | 0.000 | 0.927 | −0.002 | 0.461 | 0.007 | 0.156 |
| P. gingivalis (22) | 0.003 | 0.341 | 0.002 | 0.576 | 0.005 | 0.384 |
| P. gingivalis (23) | −0.001 | 0.815 | −0.001 | 0.741 | 0.001 | 0.916 |
| P. gingivalis (24) | 0.002 | 0.380 | 0.002 | 0.387 | 0.002 | 0.724 |
| S. cristatus (25) | 0.011 | <0.0005 | 0.011 | <0.0005 | 0.012 | 0.048 |
| S. gordonii a (26) | −0.003 | 0.012 | −0.008 | <0.0005 | 0.009 | 0.010 |
| S. intermedius (27) | −0.005 | 0.026 | −0.008 | 0.001 | 0.004 | 0.300 |
| S. mitis a (28) | 0.004 | 0.123 | 0.002 | 0.428 | 0.008 | 0.095 |
| S. mutans a (29) | 0.008 | <0.0005 | 0.003 | 0.024 | 0.022 | <0.0005 |
| S. mutans (30) | 0.005 | <0.0005 | 0.002 | 0.026 | 0.014 | <0.0005 |
| S. oralis a (31) | −0.004 | <0.0005 | −0.004 | <0.0005 | −0.004 | 0.053 |
| S. salivarius (32) | −0.012 | <0.0005 | −0.017 | <0.0005 | 0.004 | 0.014 |
| S. sanguinis a (33) | −0.007 | <0.0005 | −0.010 | <0.0005 | 0.002 | 0.041 |
| S. sobrinus (34) | −0.006 | <0.0005 | −0.009 | <0.0005 | 0.005 | 0.003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esberg, A.; Johansson, A.; Claesson, R.; Johansson, I. 43-Year Temporal Trends in Immune Response to Oral Bacteria in a Swedish Population. Pathogens 2020, 9, 544. https://doi.org/10.3390/pathogens9070544
Esberg A, Johansson A, Claesson R, Johansson I. 43-Year Temporal Trends in Immune Response to Oral Bacteria in a Swedish Population. Pathogens. 2020; 9(7):544. https://doi.org/10.3390/pathogens9070544
Chicago/Turabian StyleEsberg, Anders, Anders Johansson, Rolf Claesson, and Ingegerd Johansson. 2020. "43-Year Temporal Trends in Immune Response to Oral Bacteria in a Swedish Population" Pathogens 9, no. 7: 544. https://doi.org/10.3390/pathogens9070544
APA StyleEsberg, A., Johansson, A., Claesson, R., & Johansson, I. (2020). 43-Year Temporal Trends in Immune Response to Oral Bacteria in a Swedish Population. Pathogens, 9(7), 544. https://doi.org/10.3390/pathogens9070544