Mycoplasma bovis in Spanish Cattle Herds: Two Groups of Multiresistant Isolates Predominate, with One Remaining Susceptible to Fluoroquinolones

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. M. bovis Circulating in Spanish Beef and Dairy Herds Belongs to STs 2 and 3
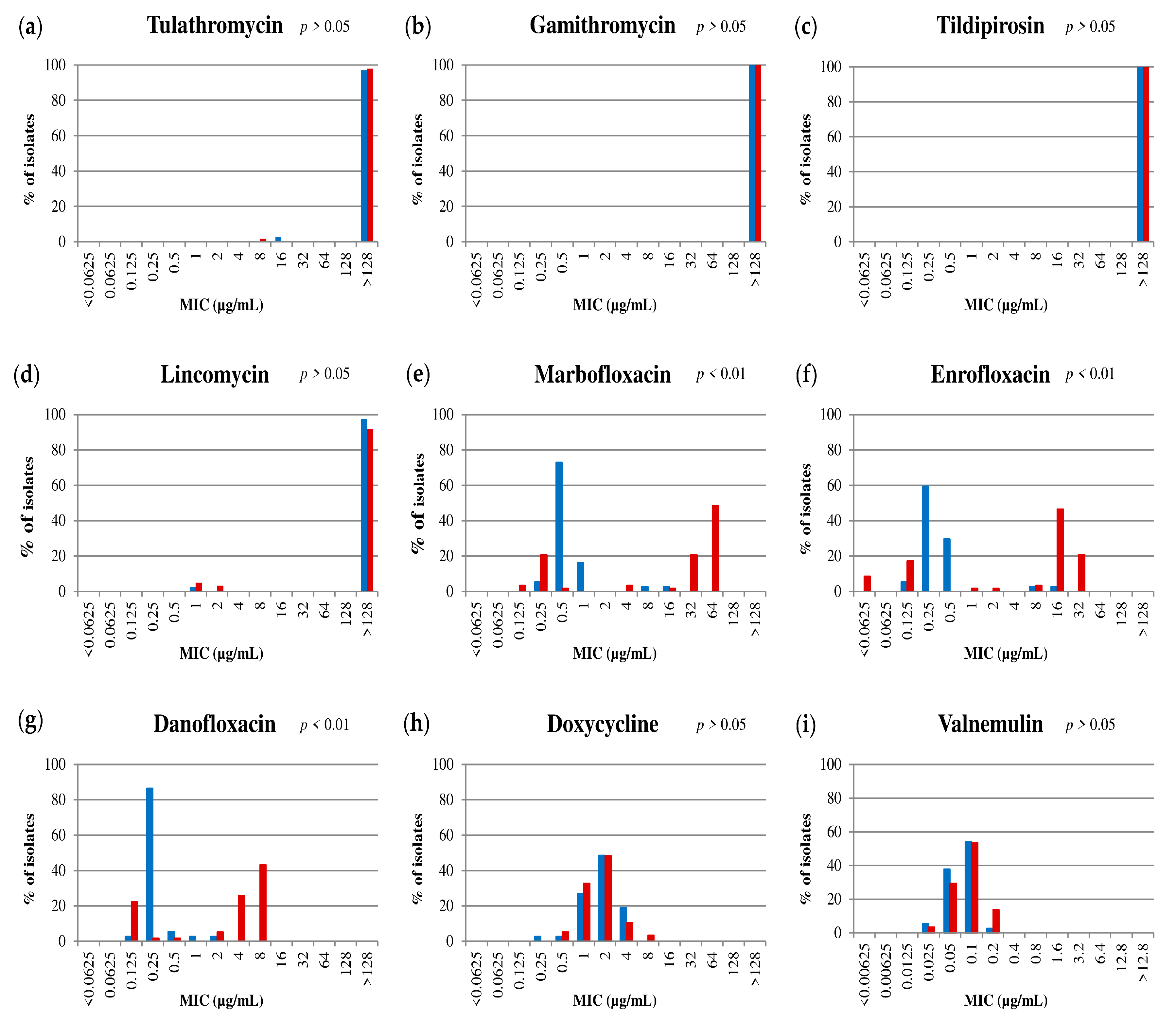
2.2. The Antimicrobial Susceptibility Profiles of The Spanish Isolates to FLQ Differ Between PolC ST2 and ST3
2.3. Analysis of Point Mutations Conferring Resistance to Antimicrobials: The Main Differences between ST2 and ST3 Are Found in The QRDR of GyrA and ParC Genes
3. Discussion
4. Materials and Methods
4.1. Animal Sampling
4.2. Mycoplasma Isolation and Subtyping
4.3. MIC Assays
4.4. Statistical Analysis
4.5. Whole-Genome Sequencing
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Hale, H.H.; Helmboldt, C.F.; Plastridge, W.N.; Stula, E.F. Bovine mastitis caused by a Mycoplasma species. Cornell Vet. 1962, 52, 582–591. [Google Scholar] [PubMed]
- Nicholas, R.A.J.; Ayling, R.D. Mycoplasma bovis: Disease, diagnosis, and control. Res. Vet. Sci. 2003, 74, 105–112. [Google Scholar] [CrossRef]
- Maunsell, F.P.; Woolums, A.R.; Francoz, D.; Rosenbusch, R.F.; Step, D.L.; Wilson, D.J.; Janzen, E.D. Mycoplasma bovis infections in cattle. J. Vet. Intern. Med. 2011, 25, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Perez-Casal, J.; Prysliak, T.; Maina, T.; Suleman, M.; Jimbo, S. Status of the development of a vaccine against Mycoplasma bovis. Vaccine 2017, 35, 2902–2907. [Google Scholar] [CrossRef]
- Gerchman, I.; Levisohn, S.; Mikula, I.; Lysnyansky, I. In vitro antimicrobial susceptibility of Mycoplasma bovis isolated in Israel from local and imported cattle. Vet. Microbiol. 2009, 137, 268–275. [Google Scholar] [CrossRef]
- Soehnlen, M.K.; Kunze, M.E.; Karunathilake, K.E.; Henwood, B.M.; Kariyawasam, S.; Wolfgang, D.R.; Jayarao, B.M. In vitro antimicrobial inhibition of Mycoplasma bovis isolates submitted to the Pennsylvania Animal Diagnostic Laboratory using flow cytometry and a broth microdilution method. J. Vet. Diagn. 2011, 23, 547–551. [Google Scholar] [CrossRef] [Green Version]
- Hendrick, S.H.; Bateman, K.G.; Rosengren, L.B. The effect of antimicrobial treatment and preventive strategies on bovine respiratory disease and genetic relatedness and antimicrobial resistance of Mycoplasma bovis isolates in a western Canadian feedlot. Can. Vet. J. Rev. Vet. Can. 2013, 54, 1146–1156. [Google Scholar]
- Ayling, R.D.; Rosales, R.S.; Barden, G.; Gosney, F.L. Changes in antimicrobial susceptibility of Mycoplasma bovis isolates from Great Britain. Vet. Rec. 2014, 175, 486. [Google Scholar] [CrossRef]
- Gautier-Bouchardon, A.V.; Ferré, S.; Le Grand, D.; Paoli, A.; Gay, E.; Poumarat, F. Overall decrease in the susceptibility of Mycoplasma bovis to antimicrobials over the past 30 years in France. PLoS ONE 2014, 9, e87672. [Google Scholar] [CrossRef] [Green Version]
- Kawai, K.; Higuchi, H.; Iwano, H.; Iwakuma, A.; Onda, K.; Sato, R.; Hayashi, T.; Nagahata, H.; Oshida, T. Antimicrobial susceptibilities of Mycoplasma isolated from bovine mastitis in Japan. Anim. Sci. J. 2014, 85, 96–99. [Google Scholar] [CrossRef]
- Sulyok, K.M.; Kreizinger, Z.; Fekete, L.; Hrivnák, V.; Magyar, T.; Jánosi, S.; Schweitzer, N.; Turcsányi, I.; Makrai, L.; Erdélyi, K.; et al. Antibiotic susceptibility profiles of Mycoplasma bovis strains isolated from cattle in Hungary, Central Europe. BMC Vet. Res. 2014, 10, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuvelink, A.; Reugebrink, C.; Mars, J. Antimicrobial susceptibility of Mycoplasma bovis isolates from veal calves and dairy cattle in the Netherlands. Vet. Microbiol. 2016, 189, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.C.; Gao, D.; Jia, B.Y.; Wang, Z.; Gao, Y.-H.; Pei, Z.-H.; Liu, S.-M.; Xin, J.-Q.; Ma, H.-X. Antimicrobial susceptibility and molecular characterization of macrolide resistance of Mycoplasma bovis isolates from multiple provinces in China. J. Vet. Med. Sci. 2016, 78, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Jelinski, M.; Kinnear, A.; Gesy, K.; Andrés-Lasheras, S.; Zaheer, R.; Weese, S.; McAllister, T.A. Antimicrobial sensitivity testing of Mycoplasma bovis isolates derived from Western Canadian feedlot cattle. Microorganisms 2020, 8, 124. [Google Scholar] [CrossRef] [Green Version]
- Citti, C.; Dordet-Frisoni, E.; Nouvel, L.X.; Kuo, C.; Baranowski, E. Horizontal gene transfers in mycoplasmas (Mollicutes). Curr. Issues Mol. Biol. 2018, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faucher, M.; Nouvel, L.-X.; Dordet-Frisoni, E.; Sagné, E.; Baranowski, E.; Hygonenq, M.-C.; Marenda, M.-S.; Tardy, F.; Citti, C. Mycoplasmas under experimental antimicrobial selection: The unpredicted contribution of horizontal chromosomal transfer. PLoS Genet. 2019, 15, e1007910. [Google Scholar] [CrossRef] [Green Version]
- McCormack, W.M. Susceptibility of mycoplasmas to antimicrobial agents: Clinical implications. Clin. Infect. Dis. 1993, 17 (Suppl. S1), S200–S201. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadeau, A.P.; Mouches, C.; Bove, J.M. Probable insensitivity of mollicutes to rifampin and characterization of spiroplasmal DNA-dependent RNA polymerase. J. Bacteriol. 1986, 166, 824–828. [Google Scholar] [CrossRef] [Green Version]
- Pellegrin, J.L.; Maugein, J.; Clerc, M.T.; Leng, B.; Bové, J.M.; Bébéar, C. Activity of rifampin against Mollicutes, clostridia and L forms. Recent advances in mycoplasmology. Zentralbl. Bakteriol. Suppl. 1990, 20, 810–812. [Google Scholar]
- Shepard, M.C.; Lunceford, C.D.; Ford, D.K.; Purcell, R.H.; Taylor-Robinson, D.; Razin, S.; Black, F.T. Ureaplasma urealyticum gen. nov., sp. nov.: Proposed nomenclature for the human T (T-Strain) mycoplasmas. Int. J. Syst. Evol. Bacteriol. 1974, 24, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Gautier-Bouchardon, A.V. Antimicrobial resistance in mycoplasma spp. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Hannan, P.C. Guidelines and recommendations for antimicrobial minimum inhibitory concentration (MIC) testing against veterinary mycoplasma species. International research programme on comparative mycoplasmology. Vet. Res. 2000, 31, 373–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lysnyansky, I.; Ayling, R.D. Mycoplasma bovis: Mechanisms of resistance and trends in antimicrobial susceptibility. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Arcangioli, M.-A.; Duet, A.; Meyer, G.; Dernburg, A.; Bézille, P.; Poumarat, F.; Le Grand, D. The role of Mycoplasma bovis in bovine respiratory disease outbreaks in veal calf feedlots. Vet. J. 2008, 177, 89–93. [Google Scholar] [CrossRef]
- Nicholas, R.A.J. Bovine mycoplasmosis: Silent and deadly. Vet. Rec. 2011, 168, 459–462. [Google Scholar] [CrossRef]
- Radaelli, E.; Luini, M.; Loria, G.R.; Nicholas, R.A.J.; Scanziani, E. Bacteriological, serological, pathological and immunohistochemical studies of Mycoplasma bovis respiratory infection in veal calves and adult cattle at slaughter. Res. Vet. Sci. 2008, 85, 282–290. [Google Scholar] [CrossRef]
- Caswell, J.L.; Bateman, K.G.; Cai, H.Y.; Castillo-Alcala, F. Mycoplasma bovis in respiratory disease of feedlot cattle. Vet. Clin. North Am. Food Anim. Pract. 2010, 26, 365–379. [Google Scholar] [CrossRef]
- Nielsen, P.K.; Petersen, M.B.; Nielsen, L.R.; Halasa, T.; Toft, N. Latent class analysis of bulk tank milk PCR and ELISA testing for herd level diagnosis of Mycoplasma bovis. Prev. Vet. Med. 2015, 121, 338–342. [Google Scholar] [CrossRef]
- Spergser, J.; Macher, K.; Kargl, M.; Lysnyansky, I.; Rosengarten, R. Emergence, re-emergence, spread and host species crossing of Mycoplasma bovis in the Austrian Alps caused by a single endemic strain. Vet. Microbiol. 2013, 164, 299–306. [Google Scholar] [CrossRef]
- Van Engelen, E.; Dijkman, R.; Holzhauer, M.; Junker, K.; Van Wuyckhuise, L.; Gonggrijp, M. Typing of Mycoplasma bovis from arthritis outbreaks in dairy herds. In Proceedings of the European Mycoplasma Meeting: Progress in Human and Animal Mycoplasmology, Pendik, Istanbul, Turkey, 3–5 June 2015. [Google Scholar]
- Aebi, M.; van den Borne, B.H.P.; Raemy, A.; Steiner, A.; Pilo, P.; Bodmer, M. Mycoplasma bovis infections in Swiss dairy cattle: A clinical investigation. Acta Vet. Scand. 2015, 57, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arcangioli, M.A.; Chazel, M.; Sellal, E.; Botrel, M.A.; Bezille, P.; Poumarat, F.; Calavas, D.; Le Grand, D. Prevalence of Mycoplasma bovis udder infection in dairy cattle: Preliminary field investigation in southeast France. N. Z. Vet. J. 2011, 59, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Filioussis, G.; Christodoulopoulos, G.; Thatcher, A.; Petridou, V.; Bourtzi-Chatzopoulou, E. Isolation of Mycoplasma bovis from bovine clinical mastitis cases in Northern Greece. Vet. J. 2007, 173, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, R.A.J.; Fox, L.K.; Lysnyansky, I. Mycoplasma mastitis in cattle: To cull or not to cull. Vet. J. 2016, 216, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Passchyn, P.; Piepers, S.; De Meulemeester, L.; Boyen, F.; Haesebrouck, F.; De Vliegher, S. Between-herd prevalence of Mycoplasma bovis in bulk milk in Flanders, Belgium. Res. Vet. Sci. 2012, 92, 219–220. [Google Scholar] [CrossRef] [Green Version]
- Klein, U.; de Jong, A.; Moyaert, H.; El Garch, F.; Leon, R.; Richard-Mazet, A.; Rose, M.; Maes, D.; Pridmore, A.; Thomson, J.R.; et al. Antimicrobial susceptibility monitoring of Mycoplasma hyopneumoniae and Mycoplasma bovis isolated in Europe. Vet. Microbiol. 2017, 204, 188–193. [Google Scholar] [CrossRef]
- Klein, U.; de Jong, A.; Youala, M.; El Garch, F.; Stevenin, C.; Moyaert, H.; Rose, M.; Catania, S.; Gyuranecz, M.; Pridmore, A.; et al. New antimicrobial susceptibility data from monitoring of Mycoplasma bovis isolated in Europe. Vet. Microbiol. 2019, 238, 108432. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y Alimentación. Available online: https://www.mapa.gob.es/ (accessed on 20 April 2020).
- Haapala, V.; Pohjanvirta, T.; Vähänikkilä, N.; Halkilahti, J.; Simonen, H.; Pelkonen, S.; Soveri, T.; Simojoki, H.; Autio, T. Semen as a source of Mycoplasma bovis mastitis in dairy herds. Vet. Microbiol. 2018, 216, 60–66. [Google Scholar] [CrossRef]
- Becker, C.A.M.; Thibault, F.M.; Arcangioli, M.-A.; Tardy, F. Loss of diversity within Mycoplasma bovis isolates collected in France from bovines with respiratory diseases over the last 35 years. Infect. Genet. Evol. 2015, 33, 118–126. [Google Scholar] [CrossRef]
- Khalil, D.; Becker, C.A.M.; Tardy, F. Alterations in the quinolone resistance-determining regions and fluoroquinolone resistance in clinical isolates and laboratory-derived Mutants of Mycoplasma bovis: Not all genotypes may be equal. Appl. Environ. Microbiol. 2016, 82, 1060–1068. [Google Scholar] [CrossRef] [Green Version]
- Khalil, D.; Becker, C.A.M.; Tardy, F. Monitoring the decrease in susceptibility to ribosomal RNAs targeting antimicrobials and its molecular basis in clinical Mycoplasma bovis isolates over time. Microb. Drug Resist. Larchmt. N 2017, 23, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Lysnyansky, I.; Mikula, I.; Gerchman, I.; Levisohn, S. Rapid detection of a point mutation in the parC gene associated with decreased susceptibility to fluoroquinolones in Mycoplasma bovis. Antimicrob. Agents Chemother. 2009, 53, 4911–4914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, R.; Qi, J.; Ba, X.; Chen, Y.; Hu, C.; Liu, X.; Tu, L.; Peng, Q.; Chen, H.; Guo, A. In vitro quinolones susceptibility analysis of Chinese Mycoplasma bovis isolates and their phylogenetic scenarios based upon QRDRs of DNA topoisomerases revealing a unique transition in ParC. Pak. Vet. J. 2013, 33, 364–369. [Google Scholar]
- Sato, T.; Okubo, T.; Usui, M.; Higuchi, H.; Tamura, Y. Amino acid substitutions in GyrA and ParC are associated with fluoroquinolone resistance in Mycoplasma bovis isolates from Japanese dairy calves. J. Vet. Med. Sci. 2013, 75, 1063–1065. [Google Scholar] [CrossRef] [Green Version]
- Sulyok, K.M.; Kreizinger, Z.; Wehmann, E.; Lysnyansky, I.; Bányai, K.; Marton, S.; Jerzsele, Á.; Rónai, Z.; Turcsányi, I.; Makrai, L.; et al. Mutations associated with decreased susceptibility to seven antimicrobial families in field and laboratory-derived Mycoplasma bovis strains. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Hata, E.; Harada, T.; Itoh, M. Relationship between antimicrobial susceptibility and multilocus sequence type of Mycoplasma bovis isolates and development of a method for rapid detection of point mutations involved in decreased susceptibility to macrolides, lincosamides, tetracyclines, and spectinomycin. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Lerner, U.; Amram, E.; Ayling, R.D.; Mikula, I.; Gerchman, I.; Harrus, S.; Teff, D.; Yogev, D.; Lysnyansky, I. Acquired resistance to the 16-membered macrolides tylosin and tilmicosin by Mycoplasma bovis. Vet. Microbiol. 2014, 168, 365–371. [Google Scholar] [CrossRef]
- Lysnyansky, I.; Gerchman, I.; Flaminio, B.; Catania, S. Decreased susceptibility to macrolide-lincosamide in mycoplasma synoviae is associated with mutations in 23S ribosomal RNA. Microb. Drug Resist. 2015, 21, 581–589. [Google Scholar] [CrossRef]
- Amram, E.; Mikula, I.; Schnee, C.; Ayling, R.D.; Nicholas, R.A.J.; Rosales, R.S.; Harrus, S.; Lysnyansky, I. 16S rRNA gene mutations associated with decreased susceptibility to tetracycline in Mycoplasma bovis. Antimicrob. Agents Chemother. 2015, 59, 796–802. [Google Scholar] [CrossRef] [Green Version]
- CIMAVet. Centro de Información de Medicamentos para Veterinaria. Available online: https://cimavet.aemps.es/ (accessed on 20 April 2020).
- Stipkovits, L.; Ripley, P.H.; Tenk, M.; Glávits, R.; Molnár, T.; Fodor, L. The efficacy of valnemulin (Econor) in the control of disease caused by experimental infection of calves with Mycoplasma bovis. Res. Vet. Sci. 2005, 78, 207–215. [Google Scholar] [CrossRef]
- Waites, K.B.; Bébéar, C.M.; Robertson, J.A.; Talkington, D.F.; Kenny, G.E. Cumitech 34: Laboratory Diagnosis of Mycoplasmal Infections; American Society for Microbiology: Washington, WA, USA, 2001. [Google Scholar]
- Tola, S.; Angioi, A.; Rocchigiani, A.M.; Idini, G.; Manunta, D.; Galleri, G.; Leori, G. Detection of Mycoplasma agalactiae in sheep milk samples by polymerase chain reaction. Vet. Microbiol. 1997, 54, 17–22. [Google Scholar] [CrossRef]
- Foddai, A.; Idini, G.; Fusco, M.; Rosa, N.; De la Fe, C.; Zinellu, S.; Corona, L.; Tola, S. Rapid differential diagnosis of Mycoplasma agalactiae and Mycoplasma bovis based on a multiplex-PCR and a PCR-RFLP. Mol. Cell. Probes 2005, 19, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Martín, A.; De la Fe, C.; Amores, J.; Sánchez, A.; Contreras, A.; Paterna, A.; Buendía, A.J.; Corrales, J.C. Anatomic location of Mycoplasma mycoides subsp. capri and Mycoplasma agalactiae in naturally infected goat male auricular carriers. Vet. Microbiol. 2012, 157, 355–362. [Google Scholar] [CrossRef]
- Albers, A.C.; Fletcher, R.D. Simple method for quantitation of viable mycoplasmas. Appl. Environ. Microbiol. 1982, 43, 958–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epi InfoTM|CDC. Available online: https://www.cdc.gov/epiinfo/index.html (accessed on 20 April 2020).
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 20 April 2020).
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Deatherage, D.E.; Barrick, J.E. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. In Engineering and Analyzing Multicellular Systems; Humana Press: New York, NY, USA, 2014; Volume 1151, pp. 165–188. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| polCa ST | MIC Parameter | Macrolides | Lincosamide | Fluoroquinolones | Tetracycline | Pleuromutilin | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tul | Gam | Tild | Lin | Enr | Marb | Dan | Dox | Val | ||
| 1 PG45 | MIC | 1 | 8 | 1 | 1 | 0.125 | 0.5 | 0.125 | 0.0625 | 0.025 |
| 2 (n = 37) | MIC Range | 16–>128 | >128 | >128 | 1–>128 | 0.125–16 | 0.25–16 | 0.125–2 | 0.25–4 | 0.025–0.2 |
| MIC50 | >128 | >128 | >128 | >128 | 0.25 | 0.5 | 0.25 | 2 | 0.1 | |
| MIC90 | >128 | >128 | >128 | >128 | 0.5 | 1 | 0.5 | 4 | 0.1 | |
| 3 (n = 58) | MIC Range | 8–>128 | >128 | >128 | 1–>128 | <0.0625–32 | 0.125–64 | 0.125–8 | 0.5–8 | 0.025–0.2 |
| MIC50 | >128 | >128 | >128 | >128 | 16 | 32 | 4 | 2 | 0.1 | |
| MIC90 | >128 | >128 | >128 | >128 | 32 | 64 | 8 | 4 | 0.2 | |
| Isolate | polCa ST | gyrA | gyrB | parC | MIC (µg/mL) b | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 83 c | 362 | 323 | 10 | 80 c | 81 c | 84 c | 116 | 156 | Enr | Marb | Dan | ||
| PG45 | 1 | Ser | Asp | Val | Gln | Ser | Ser | Asp | Ala | Val | 0.125 | 0.5 | 0.125 |
| J335 | 3 | - | Asn | - | Arg | - | - | - | - | - | <0.0625 | 0.25 | 0.125 |
| J403 | 3 | - | Asn | - | Arg | - | - | - | - | - | <0.0625 | 0.25 | 0.125 |
| J414 | 3 | - | Asn | - | Arg | - | - | - | - | - | <0.0625 | 0.25 | 0.125 |
| J433 | 3 | - | Asn | - | Arg | - | - | - | - | - | 0.125 | 0.25 | 0.125 |
| J341 | 2 | - | Asn | - | - | - | - | - | - | - | 0.125 | 0.25 | 0.25 |
| J6 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J103 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J175 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J226 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J276 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J319 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J330 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J336 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.5 |
| J356 | 2 | - | Asn | - | - | - | - | - | - | - | 0.25 | 0.5 | 0.25 |
| J136 | 2 | - | Asn | - | - | - | - | - | - | - | 0.5 | 1 | 0.25 |
| J137 | 2 | - | Asn | - | - | - | - | - | - | - | 0.5 | 0.5 | 0.125 |
| J368 | 2 | - | Asn | - | - | - | - | - | - | - | 0.5 | 1 | 0.25 |
| J377 | 2 | - | Asn | - | - | - | - | - | - | - | 0.5 | 1 | 0.25 |
| J391 | 2 | - | Asn | - | - | - | - | - | - | - | 0.5 | 0.5 | 0.25 |
| J410 | 2 | - | Asn | - | - | - | - | - | - | - | 0.5 | 0.5 | 0.25 |
| J279 | 3 | Phe | Asn | - | Arg | - | - | - | Pro | - | 1 | 4 | 4 |
| J228 | 3 | Phe | Asn | - | Arg | - | - | - | Pro | - | 2 | 4 | 2 |
| J115 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 8 | 32 | 2 |
| J28 | 3 | Phe | Asn | - | Arg | - | Pro | Asn | - | - | 16 | 64 | 8 |
| J69 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 16 | 32 | 4 |
| J72 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 16 | 64 | 8 |
| J81 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 16 | 32 | 4 |
| J96 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 16 | 32 | 4 |
| J131 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 16 | 64 | 8 |
| J305 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 16 | 64 | 8 |
| J178 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 32 | 64 | 8 |
| J233 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 32 | 64 | 8 |
| J295 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 32 | 64 | 8 |
| J388 | 3 | Phe | Asn | - | Arg | Ile | - | - | - | - | 32 | 64 | 8 |
| J479 | 3 | Phe | - | Ala | Arg | Ile | - | - | - | Ile | 32 | 64 | 8 |
| J482 | 3 | Phe | - | Ala | Arg | Ile | - | - | - | Ile | 32 | 64 | 8 |
| Isolate | polCa ST | 23S rRNA, rrl alleles b | L3 c | L4 c | L22 c | MIC (µg/mL) d | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 452 | 534 | 748 e | 954 | 1249 | 1251 | 2058 e,f | 2157 | 2848 | 265 | 11 | 24 | 36 | 44 | 62 | 63 | 68 | 79 | 94 | 178 | 178 | 178 | 5 | 93 e | Tul | Gam | Tild | Lin | Val | ||
| PG45 | 1 | G | A | G | G | T | A | A | G | G | Ala | Ser | Thr | Thr | Ala | Val | Ala | Glu | Ala | Ala | Gly | Gly | Gly | Gln | Gln | 1 | 8 | 1 | 1 | 0.025 |
| J137 | 2 | - | T ** | A ** | - | C * | - | - | - | - | Val | Thr | - | Ala | Thr | Ala | Thr | Ala | Thr | Thr | - | - | Val | - | His | 16 | >128 | >128 | 1 | 0.1 |
| J28 | 3 | - | T ** | A ** | - | C * | - | - | - | - | Val | Thr | - | Ala | Thr | Ala | Thr | Ala | Thr | Thr | - | Leu | - | - | His | >128 | >128 | >128 | 1 | 0.05 |
| J403 | 3 | - | T ** | A ** | - | C * | - | G * | - | - | Val | Thr | - | - | Thr | Ala | Thr | Ala | Thr | Thr | Arg | - | - | - | His | >128 | >128 | >128 | >128 | 0.1 |
| J414 | 3 | - | T ** | A ** | - | C * | - | G * | - | - | Val | Thr | - | Ala | Thr | Ala | Thr | Ala | Thr | Thr | Arg | - | - | - | His | >128 | >128 | >128 | >128 | 0.1 |
| J433 | 3 | - | T ** | A ** | - | C * | - | G * | - | - | Val | Thr | - | Ala | Thr | Ala | Thr | Ala | Thr | Thr | Arg | - | - | - | His | >128 | >128 | >128 | >128 | 0.1 |
| J6 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J103 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | Arg | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.025 |
| J136 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | Lys | - | >128 | >128 | >128 | >128 | 0.05 |
| J175 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J226 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J276 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J319 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J330 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J336 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J341 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J356 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J368 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J377 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J391 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.2 |
| J410 | 2 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J96 | 3 | - | T ** | A ** | A * | C * | - | G ** | A * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J178 | 3 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J228 | 3 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | Thr | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J233 | 3 | - | T ** | A ** | A * | C * | - | G ** | A * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J279 | 3 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J295 | 3 | - | T ** | A ** | A * | C * | - | G ** | A * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J305 | 3 | - | T ** | A ** | A * | C * | - | G ** | A * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J335 | 3 | - | T ** | A ** | A * | C * | T* | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J388 | 3 | - | T ** | A ** | A * | C * | - | G ** | A * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J479 | 3 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.2 |
| J482 | 3 | - | T ** | A ** | A * | C * | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.2 |
| J69 | 3 | A * | T ** | A ** | A ** | - | - | G ** | - | - | - | - | - | - | Thr | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J72 | 3 | A * | T ** | A ** | A ** | - | - | G ** | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.05 |
| J81 | 3 | A * | T ** | A ** | A ** | - | - | G ** | - | T * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| J115 | 3 | A * | T ** | A ** | A ** | - | - | G ** | - | T * | - | - | - | - | Thr | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.025 |
| J131 | 3 | A * | T ** | A ** | A ** | - | - | G ** | - | - | - | - | - | - | Thr | - | - | - | - | - | - | - | - | - | - | >128 | >128 | >128 | >128 | 0.1 |
| Isolate | polCa ST | 16S rRNA, rrs alleles b | MIC (µg/mL) c | |||||
|---|---|---|---|---|---|---|---|---|
| 335 | 859 | 965 d | 967 d | 1192 e | 1199 | Dox | ||
| PG45 | 1 | C | C | A | A | C | T | 0.0625 |
| J137 | 2 | T * | T * | T ** | T ** | - | - | 1 |
| J28 | 3 | T * | - | T ** | T ** | - | - | 1 |
| J403 | 3 | T * | - | T ** | T ** | - | - | 1 |
| J414 | 3 | T * | - | T ** | T ** | - | - | 1 |
| J433 | 3 | T * | - | T ** | T ** | - | - | 1 |
| J276 | 2 | - | - | T ** | T ** | A ** | C ** | 1 |
| J319 | 2 | - | - | T ** | T ** | A ** | C ** | 1 |
| J341 | 2 | - | - | T ** | T ** | A ** | C ** | 1 |
| J115 | 3 | - | - | T ** | T ** | A ** | C ** | 1 |
| J335 | 3 | - | - | T ** | T ** | A ** | C ** | 1 |
| J6 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J103 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J136 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J175 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J226 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J336 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J356 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J377 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J391 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J410 | 2 | - | - | T ** | T ** | A ** | C ** | 2 |
| J69 | 3 | - | - | T ** | T ** | A ** | C ** | 2 |
| J72 | 3 | - | - | T ** | T ** | A ** | C ** | 2 |
| J81 | 3 | - | - | T ** | T ** | A ** | C ** | 2 |
| J178 | 3 | - | - | T ** | T ** | A ** | C ** | 2 |
| J228 | 3 | - | - | T ** | T ** | A ** | C * | 2 |
| J279 | 3 | - | - | T ** | T ** | A ** | C * | 2 |
| J295 | 3 | - | - | T ** | T ** | A ** | C ** | 2 |
| J305 | 3 | - | - | T ** | T ** | A ** | C ** | 2 |
| J479 | 3 | - | - | T ** | T ** | A ** | C * | 2 |
| J482 | 3 | - | - | T ** | T ** | A ** | C * | 2 |
| J330 | 2 | - | - | T ** | T ** | A ** | C ** | 4 |
| J368 | 2 | - | - | T ** | T ** | A ** | C ** | 4 |
| J131 | 3 | - | - | T ** | T ** | A ** | C ** | 4 |
| J233 | 3 | - | - | T ** | T ** | A ** | C ** | 4 |
| J96 | 3 | - | - | T ** | T ** | A ** | C ** | 8 |
| J388 | 3 | - | - | T ** | T ** | A ** | C ** | 8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Galán, A.; Nouvel, L.-X.; Baranowski, E.; Gómez-Martín, Á.; Sánchez, A.; Citti, C.; de la Fe, C. Mycoplasma bovis in Spanish Cattle Herds: Two Groups of Multiresistant Isolates Predominate, with One Remaining Susceptible to Fluoroquinolones. Pathogens 2020, 9, 545. https://doi.org/10.3390/pathogens9070545
García-Galán A, Nouvel L-X, Baranowski E, Gómez-Martín Á, Sánchez A, Citti C, de la Fe C. Mycoplasma bovis in Spanish Cattle Herds: Two Groups of Multiresistant Isolates Predominate, with One Remaining Susceptible to Fluoroquinolones. Pathogens. 2020; 9(7):545. https://doi.org/10.3390/pathogens9070545
Chicago/Turabian StyleGarcía-Galán, Ana, Laurent-Xavier Nouvel, Eric Baranowski, Ángel Gómez-Martín, Antonio Sánchez, Christine Citti, and Christian de la Fe. 2020. "Mycoplasma bovis in Spanish Cattle Herds: Two Groups of Multiresistant Isolates Predominate, with One Remaining Susceptible to Fluoroquinolones" Pathogens 9, no. 7: 545. https://doi.org/10.3390/pathogens9070545
APA StyleGarcía-Galán, A., Nouvel, L. -X., Baranowski, E., Gómez-Martín, Á., Sánchez, A., Citti, C., & de la Fe, C. (2020). Mycoplasma bovis in Spanish Cattle Herds: Two Groups of Multiresistant Isolates Predominate, with One Remaining Susceptible to Fluoroquinolones. Pathogens, 9(7), 545. https://doi.org/10.3390/pathogens9070545