Monitoring Mycoplasma bovis Diversity and Antimicrobial Susceptibility in Calf Feedlots Undergoing a Respiratory Disease Outbreak

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
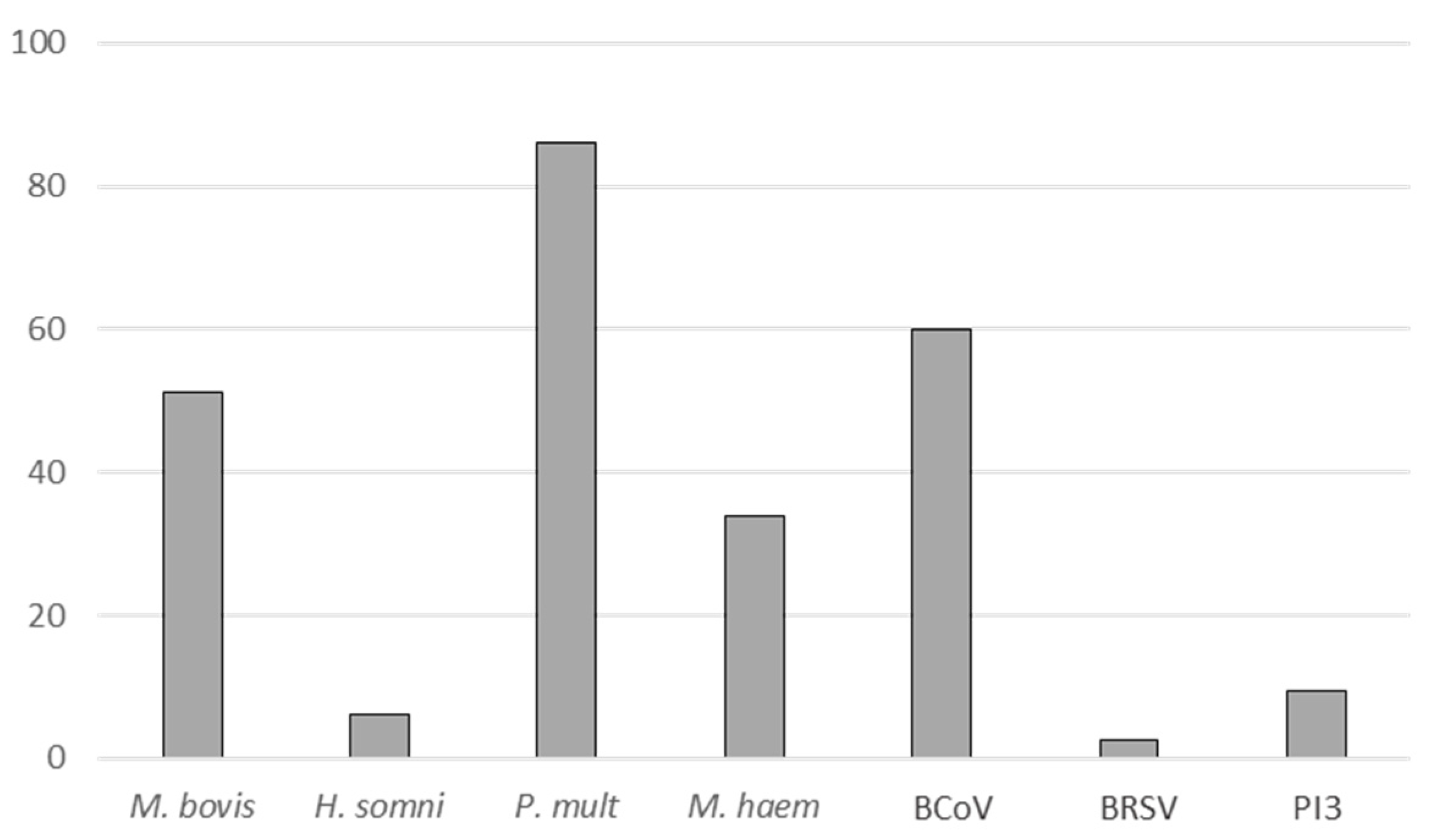
2.1. M. bovis Was the Third Most Frequently Isolated Pathogen (in Association with Others) in Calf Feedlots at BRD Onset
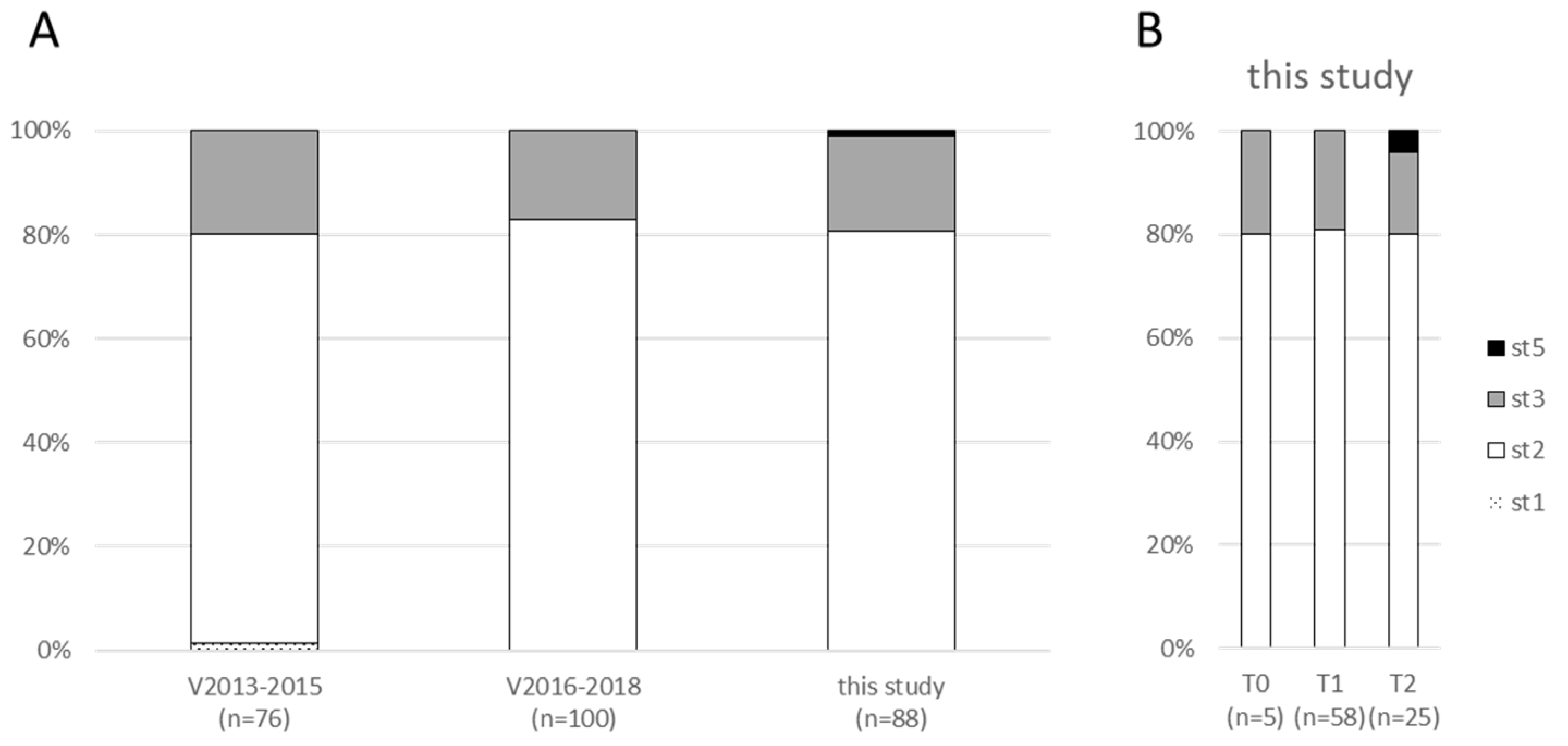
2.2. The Diversity of Clones Does Not Differ at Different Sampling Times and Is Similar to the Reference Population of Vigimyc Isolates
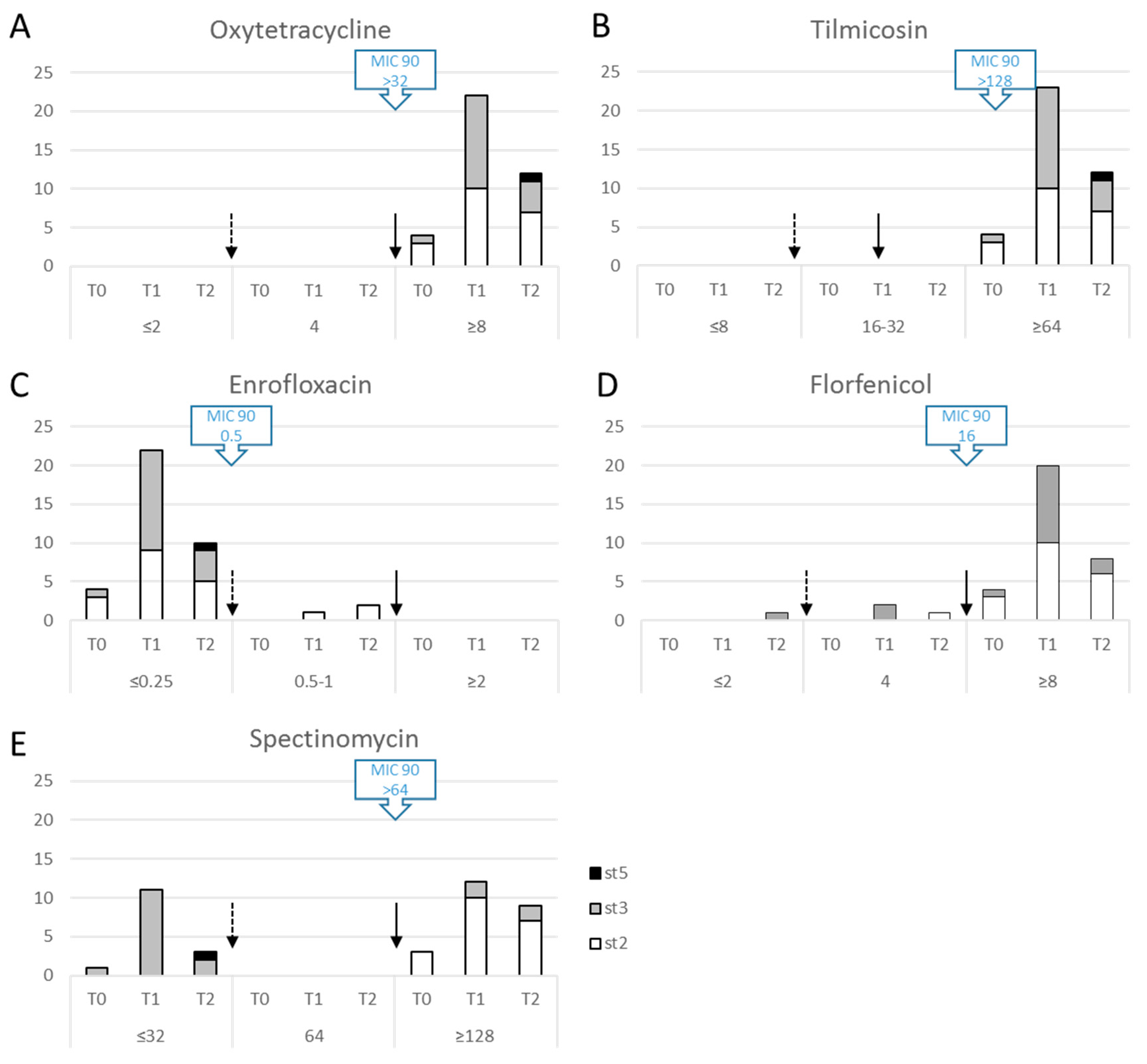
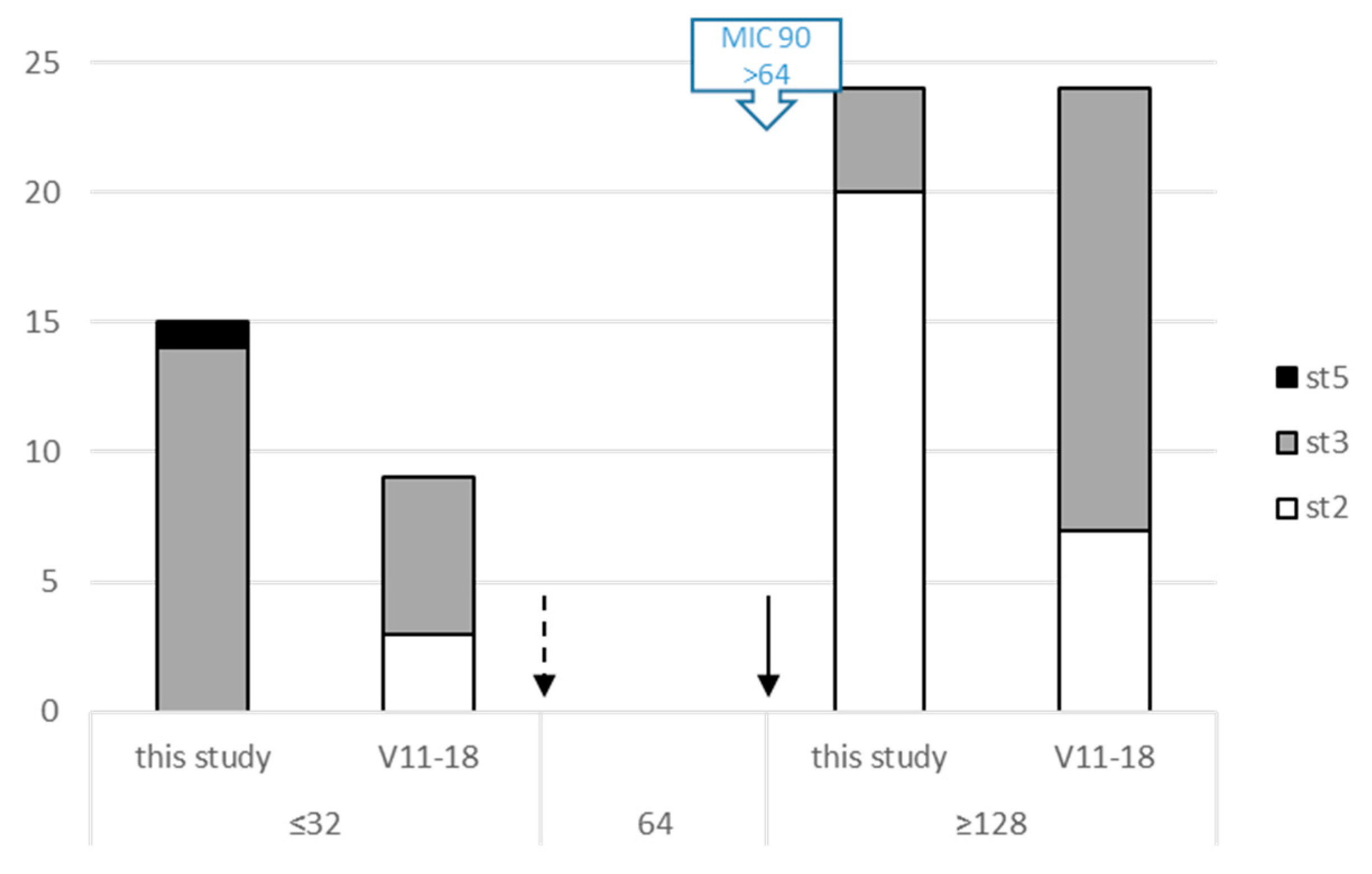
2.3. M. bovis Field Isolates Are Already Multiresistant to Antimicrobials before Any Treatment in the Lot
2.4. Antimicrobial Resistance Profile of Pasteurellaceae Is Different from That of M. bovis
3. Discussion
4. Materials and Methods
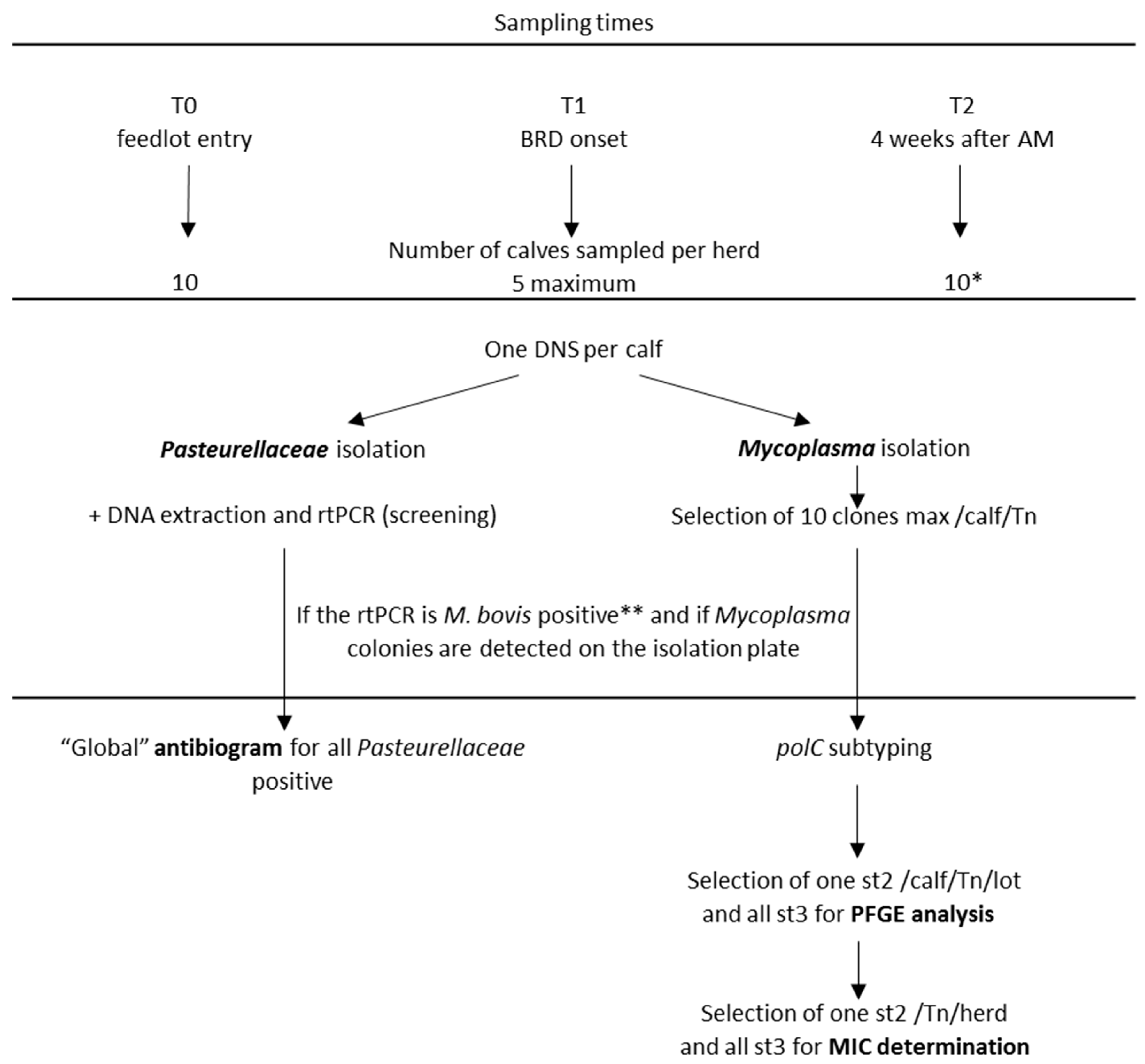
4.1. Sampling Campaigns
4.2. Nucleic Acid Extraction from Swabs and rtPCR Amplifications
4.3. Isolation and Identification of M. bovis Isolates
4.4. Strain Subtyping by Sequence Analysis of the Housekeeping Gene polC and Pulse Field Gel Electrophoresis (PFGE)
4.5. Antimicrobial Susceptibility Testing of M. bovis and Pasteurellaceae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Urban-Chmiel, R.; Grooms, D.L. Prevention and Control of Bovine Respiratory Disease. J. Livest. Sci. 2012, 3, 27–36. [Google Scholar]
- Snowder, G.D.; Van Vleck, L.D.; Cundiff, L.V.; Bennett, G.L. Bovine respiratory disease in feedlot cattle: Environmental, genetic, and economic factors. J. Anim. Sci. 2006, 84, 1999–2008. [Google Scholar] [CrossRef] [Green Version]
- Cirone, F.; Padalino, B.; Tullio, D.; Capozza, P.; Losurdo, M.; Lanave, G.; Pratelli, A. Prevalence of Pathogens Related to Bovine Respiratory Disease Before and After Transportation in Beef Steers: Preliminary Results. Animals 2019, 9, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, J.L.; Sacco, R.E. The Immunology of Bovine Respiratory Disease: Recent Advancements. Vet. Clin. N. Am. Food Anim. Pract. 2020, in press. [Google Scholar] [CrossRef]
- Grissett, G.P.; White, B.J.; Larson, R.L. Structured Literature Review of Responses of Cattle to Viral and Bacterial Pathogens Causing Bovine Respiratory Disease Complex. J. Vet. Intern. Med. 2015, 29, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.; Holman, D.B.; Timsit, E.; Schwinghamer, T.; Alexander, T.W. Evaluation of the Nasopharyngeal Microbiota in Beef Cattle Transported to a Feedlot, With a Focus on Lactic Acid-Producing Bacteria. Front. Microbiol. 2019, 10, 1988. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The epidemiology of bovine respiratory disease: What is the evidence for predisposing factors? Can. Vet. J. 2010, 51, 1095–1102. [Google Scholar] [PubMed]
- Maunsell, F.P.; Woolums, A.R.; Francoz, D.; Rosenbusch, R.F.; Step, D.L.; Wilson, D.J.; Janzen, E.D. Mycoplasma bovis infections in cattle. J. Vet. Intern. Med. 2011, 25, 772–783. [Google Scholar] [CrossRef]
- Holman, D.B.; Timsit, E.; Amat, S.; Abbott, D.W.; Buret, A.G.; Alexander, T.W. The nasopharyngeal microbiota of beef cattle before and after transport to a feedlot. BMC Microbiol. 2017, 17, 70. [Google Scholar] [CrossRef] [Green Version]
- Calcutt, M.J.; Lysnyansky, I.; Sachse, K.; Fox, L.K.; Nicholas, R.A.J.; Ayling, R.D. Gap analysis of Mycoplasma bovis disease, diagnosis and control: An aid to identify future development requirements. Transbound. Emerg. Dis. 2018, 65, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Amat, S.; Timsit, E.; Baines, D.; Yanke, J.; Alexander, T.W. Development of Bacterial Therapeutics against the Bovine Respiratory Pathogen Mannheimia haemolytica. Appl. Environ. Microbiol. 2019, 85, e01359-19. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.; Zaheer, R.; Klima, C.; McAllister, T.; Peters, D.; Niu, Y.D.; Ralston, B. Antimicrobial Resistance in Members of the Bacterial Bovine Respiratory Disease Complex Isolated from Lung Tissue of Cattle Mortalities Managed with or without the Use of Antimicrobials. Microorganisms 2020, 8, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Donder, K.D.; Apley, M.D. A Review of the Expected Effects of Antimicrobials in Bovine Respiratory Disease Treatment and Control Using Outcomes from Published Randomized Clinical Trials with Negative Controls. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 97–111. [Google Scholar] [CrossRef]
- Gautier-Bouchardon, A.V. Antimicrobial Resistance in Mycoplasma spp. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Becker, C.A.M.; Thibault, F.M.; Arcangioli, M.-A.; Tardy, F. Genotypic evolution of Mycoplasma bovis isolates collected in France over the last 35 years. In Proceedings of the Progress in Human and Animal Mycoplasmology, Istanbul, Turkey, 3–5 June 2015; p. 60. [Google Scholar]
- Gautier-Bouchardon, A.V.; Ferre, S.; Le Grand, D.; Paoli, A.; Gay, E.; Poumarat, F. Overall Decrease in the Susceptibility of Mycoplasma bovis to Antimicrobials over the Past 30 Years in France. PLoS ONE 2014, 9, e87672. [Google Scholar] [CrossRef] [Green Version]
- Khalil, D.; Becker, C.A.M.; Tardy, F. Monitoring the decrease in susceptibility to ribosomal RNAs targeting antimicrobials and its molecular basis in clinical Mycoplasma bovis isolates over time. Microb. Drug Resist. 2017, 23, 799–811. [Google Scholar] [CrossRef]
- Poumarat, F.; Jarrige, N.; Tardy, F. Principe et bilan du réseau Vigimyc consacré à l’épidémiosurveillance des mycoplasmoses des ruminants en France. Euroref. Cah. Réf. 2014, 12, 24–29. [Google Scholar]
- Khalil, D.; Becker, C.A.M.; Tardy, F. Alterations in the Quinolone Resistance-Determining Regions and Fluoroquinolone Resistance in Clinical Isolates and Laboratory-Derived Mutants of Mycoplasma bovis: Not All Genotypes May Be Equal. Appl. Environ. Microbiol. 2016, 82, 1060–1068. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Higuchi, H.; Yokota, S.-i.; Tamura, Y. Mycoplasma bovis isolates from dairy calves in Japan have less susceptibility than a reference strain to all approved macrolides associated with a point mutation (G748A) combined with multiple species-specific nucleotide alterations in 23S rRNA. Microbiol. Immunol. 2017, 61, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Okubo, T.; Usui, M.; Higuchi, H.; Tamura, Y. Amino Acid Substitutions in GyrA and ParC Are Associated with Fluoroquinolone Resistance in Mycoplasma bovis Isolates from Japanese Dairy Calves. J. Vet. Med. Sci. 2013, 75, 1063–1065. [Google Scholar] [CrossRef] [Green Version]
- Sulyok, K.M.; Kreizinger, Z.; Wehmann, E.; Lysnyansky, I.; Bányai, K.; Marton, S.; Jerzsele, Á.; Rónai, Z.; Turcsányi, I.; Makrai, L.; et al. Mutations Associated with Decreased Susceptibility to Seven Antimicrobial Families in Field and Laboratory-Derived Mycoplasma bovis Strains. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Register, K.B.; Lysnyansky, I.; Jelinski, M.D.; Boatwright, W.D.; Waldner, M.; Bayles, D.O.; Pilo, P.; Alt, D.P. Comparison of Two Multilocus Sequence Typing Schemes for Mycoplasma bovis and Revision of the PubMLST Reference Method. J. Clin. Microbiol. 2020, in press. [Google Scholar] [CrossRef]
- Register, K.B.; Thole, L.; Rosenbush, R.F.; Minion, F.C. Multilocus sequence typing of Mycoplasma bovis reveals host-specific genotypes in cattle versus bison. Vet. Microbiol. 2015, 175, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Clinical and laboratory standards institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Caswell, J.L.; Bateman, K.G.; Cai, H.Y.; Castillo-Alcala, F. Mycoplasma bovis in respiratory disease of feedlot cattle. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Campolo, M.; Desario, C.; Cirone, F.; D’abramo, M.; Lorusso, E.; Greco, G.; Mari, V.; Colaianni, M.L.; Elia, G.; et al. Respiratory Disease Associated with Bovine Coronavirus Infection in Cattle Herds in Southern Italy. J. Vet. Diagn. Investig. 2008, 20, 28–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, D.; Credille, B.; Lehenbauer, T.W.; Berghaus, R.; Aly, S.S.; Champagne, J.; Blanchard, P.; Crossley, B.; Berghaus, L.; Cochran, S.; et al. Agreement Among 4 Sampling Methods to Identify Respiratory Pathogens in Dairy Calves with Acute Bovine Respiratory Disease. J. Vet. Intern. Med. 2017, 31, 954–959. [Google Scholar] [CrossRef]
- Arcangioli, M.; Chazel, M.; Sellal, E.; Botrel, M.; Bezille, P.; Poumarat, F.; Calavas, D.; Le Grand, D. Prevalence of Mycoplasma bovis udder infection in dairy cattle: Preliminary field investigation in southeast France. N. Z. Vet. J. 2011, 59, 75–78. [Google Scholar] [CrossRef]
- Arcangioli, M.A.; Aslan, H.; Tardy, F.; Poumarat, F.; Le Grand, D. The use of pulsed-field gel electrophoresis to investigate the epidemiology of Mycoplasma bovis in French calf feedlots. Vet. J. 2012, 192, 96–100. [Google Scholar] [CrossRef]
- Register, K.B.; Jelinski, M.D.; Waldner, M.; Boatwright, W.D.; Anderson, T.K.; Hunter, D.L.; Hamilton, R.G.; Burrage, P.; Shury, T.; Bildfell, R.; et al. Comparison of multilocus sequence types found among North American isolates of Mycoplasma bovis from cattle, bison, and deer, 2007–2017. J. Vet. Diagn. Investig. 2019, 31, 899–904. [Google Scholar] [CrossRef]
- Yair, Y.; Borovok, I.; Mikula, I.; Falk, R.; Fox, L.K.; Gophna, U.; Lysnyansky, I. Genomics-based epidemiology of bovine Mycoplasma bovis strains in Israel. BMC Genom. 2020, 21, 70. [Google Scholar] [CrossRef]
- Cai, H.Y.; McDowall, R.; Parker, L.; Kaufman, E.I.; Caswell, J.L. Changes in antimicrobial susceptibility profiles of Mycoplasma bovis over time. Can. J. Vet. Res. 2019, 83, 34–41. [Google Scholar] [PubMed]
- Hata, E.; Harada, T.; Itoh, M. Relationship between Antimicrobial Susceptibility and Multilocus Sequence Type of Mycoplasma bovis Isolates and Development of a Method for Rapid Detection of Point Mutations Involved in Decreased Susceptibility to Macrolides, Lincosamides, Tetracyclines, and Spectinomycin. Appl. Environ. Microbiol. 2019, 85, e00575-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuvelink, A.; Reugebrink, C.; Mars, J. Antimicrobial susceptibility of Mycoplasma bovis isolates from veal calves and dairy cattle in the Netherlands. Vet. Microbiol. 2016, 189, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Klein, U.; de Jong, A.; Youala, M.; El Garch, F.; Stevenin, C.; Moyaert, H.; Rose, M.; Catania, S.; Gyuranecz, M.; Pridmore, A.; et al. New antimicrobial susceptibility data from monitoring of Mycoplasma bovis isolated in Europe. Vet. Microbiol. 2019, 238, 108432. [Google Scholar] [CrossRef]
- Lysnyansky, I.; Ayling, R.D. Mycoplasma bovis: Mechanisms of resistance and trends in antimicrobial susceptibility. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Sulyok, K.M.; Kreizinger, Z.; Fekete, L.; Hrivnak, V.; Magyar, T.; Janosi, S.; Schweitzer, N.; Turcsanyi, I.; Makrai, L.; Erdelyi, K.; et al. Antibiotic susceptibility profiles of Mycoplasma bovis strains isolated from cattle in Hungary, Central Europe. BMC Vet. Res. 2014, 10, 256. [Google Scholar] [CrossRef] [Green Version]
- Jelinski, M.; Kinnear, A.; Gesy, K.; Andrés-Lasheras, S.; Zaheer, R.; Weese, S.; McAllister, A.T. Antimicrobial Sensitivity Testing of Mycoplasma bovis Isolates Derived from Western Canadian Feedlot Cattle. Microorganisms 2020, 8, 124. [Google Scholar] [CrossRef] [Green Version]
- Anholt, R.M.; Klima, C.; Allan, N.; Matheson-Bird, H.; Schatz, C.; Ajitkumar, P.; Otto, S.J.; Peters, D.; Schmid, K.; Olson, M.; et al. Antimicrobial Susceptibility of Bacteria That Cause Bovine Respiratory Disease Complex in Alberta, Canada. Front. Vet. Sci. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Klein, U.; de Jong, A.; Moyaert, H.; El Garch, F.; Leon, R.; Richard-Mazet, A.; Rose, M.; Maes, D.; Pridmore, A.; Thomson, J.R.; et al. Antimicrobial susceptibility monitoring of Mycoplasma hyopneumoniae and Mycoplasma bovis isolated in Europe. Vet. Microbiol. 2017, 204, 188–193. [Google Scholar] [CrossRef]
- Woolums, A.R.; Karisch, B.B.; Frye, J.G.; Epperson, W.; Smith, D.R.; Blanton, J.; Austin, F.; Kaplan, R.; Hiott, L.; Woodley, T.; et al. Multidrug resistant Mannheimia haemolytica isolated from high-risk beef stocker cattle after antimicrobial metaphylaxis and treatment for bovine respiratory disease. Vet. Microbiol. 2018, 221, 143–152. [Google Scholar] [CrossRef]
- Van Driessche, L.; Valgaeren, B.R.; Gille, L.; Boyen, F.; Ducatelle, R.; Haesebrouck, F.; Deprez, P.; Pardon, B. A Deep Nasopharyngeal Swab Versus Nonendoscopic Bronchoalveolar Lavage for Isolation of Bacterial Pathogens from Preweaned Calves With Respiratory Disease. J. Vet. Intern. Med. 2017, 31, 946–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilei, E.M.; Bonvin-Klotz, L.; Zimmermann, L.; Ryser-Degiorgis, M.P.; Giacometti, M.; Frey, J. Validation and diagnostic efficacy of a TaqMan real-time PCR for the detection of Mycoplasma conjunctivae in the eyes of infected Caprinae. J. Microbiol. Methods 2007, 70, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Wisselink, H.J.; Smid, B.; Plater, J.; Ridley, A.; Andersson, A.M.; Aspan, A.; Pohjanvirta, T.; Vahanikkila, N.; Larsen, H.; Hogberg, J.; et al. A European interlaboratory trial to evaluate the performance of different PCR methods for Mycoplasma bovis diagnosis. BMC Vet. Res. 2019, 15, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poumarat, F.; Longchambon, D.; Martel, J.L. Application of dot immunobinding on membrane filtration (MF dot) to the study of relationships within “M. mycoides cluster” and within “glucose and arginine-negative cluster” of ruminant mycoplasmas. Vet. Microbiol. 1992, 32, 375–390. [Google Scholar] [CrossRef]
- Shimizu, T. Selective medium for the isolation of Mycoplasma bovis from nasal discharges of pneumonic calves. Res. Vet. Sci. 1983, 34, 371–373. [Google Scholar] [CrossRef]
- Poumarat, F.; Perrin, B.; Longchambon, D. Identification of ruminant mycoplasmas by dot immunobinding on membrane filtration (MF dot). Vet. Microbiol. 1991, 29, 329–338. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T0 | T1 | T2 | Total | |
|---|---|---|---|---|
| DNS | 271 | 115 | 151 | 537 |
| Calves with M. bovis rtPCR | 6 (2%) | 59 (51%) | 39/79 * (49%) | 104 |
| Calves with M. bovis culture | 5 (2%) | 60 (52%) | 28 (19%) | 93 |
| Isolated clones | 38 | 251 | 125 | 414 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, C.A.M.; Ambroset, C.; Huleux, A.; Vialatte, A.; Colin, A.; Tricot, A.; Arcangioli, M.-A.; Tardy, F. Monitoring Mycoplasma bovis Diversity and Antimicrobial Susceptibility in Calf Feedlots Undergoing a Respiratory Disease Outbreak. Pathogens 2020, 9, 593. https://doi.org/10.3390/pathogens9070593
Becker CAM, Ambroset C, Huleux A, Vialatte A, Colin A, Tricot A, Arcangioli M-A, Tardy F. Monitoring Mycoplasma bovis Diversity and Antimicrobial Susceptibility in Calf Feedlots Undergoing a Respiratory Disease Outbreak. Pathogens. 2020; 9(7):593. https://doi.org/10.3390/pathogens9070593
Chicago/Turabian StyleBecker, Claire A.M., Chloé Ambroset, Anthéa Huleux, Angélique Vialatte, Adélie Colin, Agnès Tricot, Marie-Anne Arcangioli, and Florence Tardy. 2020. "Monitoring Mycoplasma bovis Diversity and Antimicrobial Susceptibility in Calf Feedlots Undergoing a Respiratory Disease Outbreak" Pathogens 9, no. 7: 593. https://doi.org/10.3390/pathogens9070593
APA StyleBecker, C. A. M., Ambroset, C., Huleux, A., Vialatte, A., Colin, A., Tricot, A., Arcangioli, M. -A., & Tardy, F. (2020). Monitoring Mycoplasma bovis Diversity and Antimicrobial Susceptibility in Calf Feedlots Undergoing a Respiratory Disease Outbreak. Pathogens, 9(7), 593. https://doi.org/10.3390/pathogens9070593