Molecular Characterization of Clistobothrium sp. Viable Plerocercoids in Fresh Longfin Inshore Squid (Doryteuthis pealeii) and Implications for Cephalopod Inspection

 and
and
Abstract
:1. Introduction
2. Results and discussion
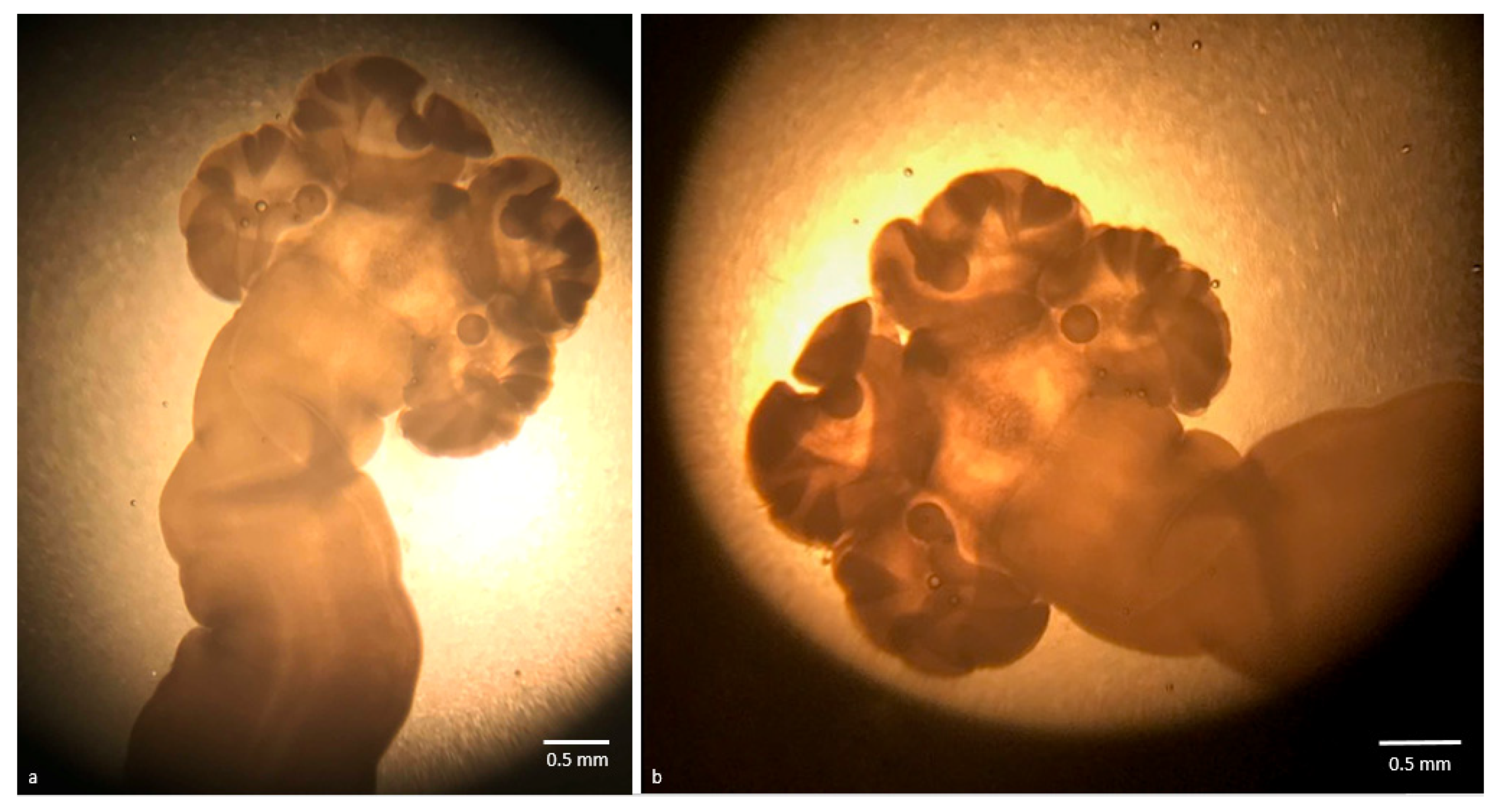
2.1. Morphological Identification
2.2. Molecular Analysis
2.2.1. Molecular Target Selection
2.2.2. Large Subunit (LSU) rRNA Gene Analysis
2.2.3. Cytochrome c Oxidase Subunit I (COI) Gene Analysis
2.3. Viable and Visible Larval Cestodes of Clistobothrium sp: Epidemiology and Implication for Cephalopod Inspection
3. Materials and Methods
3.1. Squid Sampling
3.2. Parasite Detection
3.3. DNA Extraction and Evaluation
3.4. Large Subunit (LSU) rRNA Gene Analysis
3.4.1. PCR Amplification, Sequencing and Sequences Editing
3.4.2. Comparison with Databases and Phylogenetic Analysis
3.5. Cytochrome c Oxidase Subunit I (COI) Gene Analysis
3.5.1. Primers Projecting
3.5.2. PCR Amplification, Sequencing and Sequences Editing
3.5.3. Comparison with Databases, and Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hochberg, F.G. Diseases of Mollusca: Cephalopoda. In Diseases of Marine Animals; Kinne, O., Ed.; Biologisches Anstalt Helgoland: Helgoland, Germany, 1990; Volume 3, pp. 47–227. [Google Scholar]
- Roumbedakis, K.; Krausova, M.; Tyml, T.; Di Cristo, C. A perspective around cephalopods and their parasites, and suggestions on how to increase knowledge in the field. Front. Physiol. 2018, 9, 1573. [Google Scholar] [CrossRef] [PubMed]
- Brickle, P.; Olson, P.D.; Littlewood, D.T.J.; Bishop, A.; Arkhipkin, A.I. Parasites of Loligo gahi from waters off the Falkland Islands, with a phylogenetically based identification of their cestode larvae. Can. J. Zool. 2001, 79, 2289–2296. [Google Scholar] [CrossRef] [Green Version]
- Guillén-Hernández, S.; López-Struck, A.; González-Salas, C.; Aguirre-Macedo, M.L. Octopus maya parasites off the Yucatán Peninsula, Mexico. I. Faunal assemblages. Dis. Aquat. Organ. 2018, 130, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Vieites, J.M.; Ruiz, C.S.; Fernández, F.; Alonso, R.C. Importance of cephalopod health and welfare for the commercial sector. In Handbook of Pathogens and Diseases in Cephalopods; Gestal, C., Pascual, S., Guerra, A., Fiorito, G., Vieites, J.M., Eds.; Springer: Berlin, Germany, 2019; pp. 5–7. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals. Rome 2018. Available online: http://www.fao.org/3/i9540en/I9540EN.pdf (accessed on 28 December 2019).
- FAO. Tight Supply Situation Continues. Information and Analysis on World Fish Trade. Rome 2019. Available online: http://www.fao.org/in-action/globefish/market-reports/resource-detail/en/c/1176219/ (accessed on 28 December 2019).
- Regulation (EC) No 178/2002 of the European Parliament and of the Council of 28 January 2002, laying down the general principles and requirements of food law, establishing the European Food Safety Authority, and laying down procedures in matters of food safety. Off. J. Eur. Communities 2002, 31, 1–24.
- Regulation (EC) No 852/2004 of the European Parliament and of the Council of 29 April 2004 on the hygiene of foodstuffs. Off. J. Eur. Communities 2004, 139, 1–54.
- Regulation (EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Off. J. Eur. Communities 2004, 139, 55–205.
- Council Regulation (EC) No 2406/96 of 26 November 1996 laying down common marketing standards for certain fishery products. Off. J. Eur. Communities 1996, 334, 1–15.
- Bao, M.; Pierce, G.J.; Strachan, N.J.; Pascual, S.; González-Muñoz, M.; Levsen, A. Human health, legislative and socioeconomic issues caused by the fish-borne zoonotic parasite Anisakis: Challenges in risk assessment. Trends Food Sci. Technol. 2019, 86, 298–310. [Google Scholar] [CrossRef]
- D’Amico, P.; Malandra, R.; Costanzo, F.; Castigliego, L.; Guidi, A.; Gianfaldoni, D.; Armani, A. Evolution of the Anisakis risk management in the European and Italian context. Food Res. Int. 2014, 64, 348–362. [Google Scholar] [CrossRef]
- Guardone, L.; Bilska-Zajac, E.; Castiglione, D.; Giusti, A.; Malandra, R.; Armani, A. Visible parasites in fresh cephalopods sold on the Italian market: Impact on consumers’ perception on safety and quality. In Proceedings of the Annual Scientific Conference and Annual General Meeting of the European College of Veterinary Public Health, Edinburgh, UK, 2–4 October 2019; p. 37. [Google Scholar]
- Cavaleiro, M.N.C. Parasite Fauna of Octopus vulgaris (Cephalopoda: Octopodidae) and Platichthys flesus (Actinopterygii: Oleuronectidae): Morphology, Systematics, Life History Strategies and Ecology. Ph.D. Thesis, Porto University, Porto, Portugal, 2013. Available online: https://core.ac.uk/download/pdf/143398568.pdf (accessed on 28 December 2019).
- Petrić, M.; Mladineo, I.; Šifner, S.K. Insight into the short-finned squid Illex coindetii (Cephalopoda: Ommastrephidae) feeding ecology: Is there a link between helminth parasites and food composition? J. Parasitol. 2011, 97, 55–62. [Google Scholar] [CrossRef]
- Pardo-Gandarillas, M.C.; Lohrmann, K.B.; Valdivia, A.L.; Ibáñez, C.M. First record of parasites of Dosidicus gigas (d’Orbigny, 1835) (Cephalopoda: Ommastrephidae) from the Humboldt Current system off Chile. Rev. Biol. Mar. Oceanog. 2009, 44, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Nigmatullin, C.M.; Shchetinnikov, A.S.; Shukhgalter, O.A. On feeding and helminth fauna of neon flying squid Ommastrephes bartramii (Lesueur, 1821) (Cephalopoda: Ommastrephidae) in the southeastern Pacific. Rev. Biol. Mar. Oceanog. 2009, 44, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Shukhgalter, O.A.; Nigmatullin, C.M. Parasitic helminths of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) in open waters of the central east Pacific. Fish. Res. 2001, 54, 95–110. [Google Scholar] [CrossRef]
- González, R.A.; Kroeck, M.A. Enteric helminths of the shortfin squid Illex argentinus in San Matias Gulf (Argentina) as stock discriminants. Acta Parasitol. 2000, 45, 89–93. [Google Scholar]
- Gestal, C.; Abollo, E.; Arlas, C.; Pascual, S. Estudio al MEB de larvas plerocercoides de Phyllobothrium sp. (Tetraphyllidea, Phyllobotriidae) y Nybelinia lingualis (Trypanorhyncha, Tentaculariidae), cestodos parásitos de Octopus vulgaris (Mollusca, Cephalopoda) en la Ría de Vigo. Iberus 1998, 16, 125–132. [Google Scholar]
- Pascual, S.; Gestal, C.; Estévez, J.M.; Rodríguez, H.; Soto, M.; Abollo, E.; Arias, C. Parasites in commercially-exploited cephalopods (Mollusca, Cephalopoda) in Spain: An updated perspective. Aquaculture 1996, 142, 1–10. [Google Scholar] [CrossRef]
- Pascual, S.; Lores, B.; Arias, C.; González, Á.F.; Guerra, Á. Helminth parasites of Illex coindetii (Cephalopoda: Ommastrephidae) off the Galician coast (NE Atlantic). Sci. Mar. 1994, 58, 269–272. [Google Scholar]
- Bower, S.M.; Margolis, L. Potential use of helminth parasites in stock identification of flying squid, Ommastrephes bartrami, in North Pacific waters. Can. J. Zool. 1991, 69, 1124–1126. [Google Scholar] [CrossRef]
- Caira, J.N.; Jensen, K.; Waeschenbach, A.; Olson, P.D.; Littlewood, D.T. Orders out of chaos—Molecular phylogenetics reveals the complexity of shark and stingray tapeworm relationships. Int. J. Parasitol. 2014, 44, 55–73. [Google Scholar] [CrossRef]
- Jensen, K.; Bullard, S.A. Characterization of a diversity of tetraphyllidean and rhinebothriidean cestode larval types, with comments on host associations and life-cycles. Int. J. Parasitol. 2010, 40, 889–910. [Google Scholar] [CrossRef]
- Dollfus, R.-P. Énumération des cestodes du plancton et des invertébrés marins (6e contribution). Ann. Parasitol. Hum. Comp. 1964, 39, 329–379. [Google Scholar] [CrossRef] [PubMed]
- Scholz, T.; Euzet, L.; Moravec, F. Taxonomic status of Pelichnibothrium speciosum Monticelli, 1889 (Cestoda: Tetraphyllidea), a mysterious parasite of Alepisaurus ferox Lowe (Teleostei: Alepisauridae) and Prionace glauca (L.) (Euselachii: Carcharinidae). Syst. Parasitol. 1998, 41, 1–8. [Google Scholar] [CrossRef]
- Caira, J.N.; Pickering, M. Cestodes from deep-water squaliform sharks in the Azores. Deep Sea Res. II 2013, 98, 170–177. [Google Scholar] [CrossRef]
- Caira, J.N.; Jensen, K. A digest of elasmobranch tapeworms. J. Parasitol. 2014, 100, 373–391. [Google Scholar] [CrossRef]
- Stunkard, H.W. Studies on Tetraphyllidean and Tetrarhynchidean metacestodes from squids taken on the New England coast. Bio. Bull. 1977, 153, 387–412. [Google Scholar] [CrossRef]
- Agusti, C.; Aznar, F.J.; Olson, P.D.; Littlewood, D.T.J.; Kostadinova, A.; Raga, J.A. Morphological and molecular characterization of tetraphyllidean merocercoids (Platyhelminthes: Cestoda) of striped dolphins (Stenella coeruleoalba) from the Western Mediterranean. Parasitology 2005, 130, 461–474. [Google Scholar] [CrossRef] [Green Version]
- Aznar, F.J.; Agustí, C.; Littlewood, D.T.J.; Raga, J.A.; Olson, P.D. Insight into the role of cetaceans in the life cycle of the tetraphyllideans (Platyhelminthes: Cestoda). Int. J. Parasitol. 2007, 37, 243–255. [Google Scholar] [CrossRef]
- Caira, J.N.; Jensen, K.; Hayes, C.; Ruhnke, T.R. Insights from new cestodes of the crocodile shark, Pseudocarcharias kamoharai (Lamniformes: Pseudocarchariidae), prompt expansion of Scyphyophyllidum and formal synonymization of seven phyllobothriidean genera–at last! J. Helminthol. 2020, 94, 1–25. [Google Scholar] [CrossRef]
- Klotz, D.; Hirzmann, J.; Bauer, C.; Schöne, J.; Iseringhausen, M.; Wohlsein, P.; Baumgärtner, W.; Herder, V. Subcutaneous merocercoids of Clistobothrium sp. in two Cape fur seals (Arctocephalus pusillus pusillus). Int. J. Parasitol. Parasites Wildl. 2018, 7, 99–105. [Google Scholar] [CrossRef]
- Randhawa, H.S.; Brickle, P. Larval parasite gene sequence data reveal cryptic trophic links in life cycles of porbeagle shark tapeworms. Mar. Ecol. Prog. Ser. 2011, 431, 215–222. [Google Scholar] [CrossRef]
- Randhawa, H.S. Insights using a molecular approach into the life cycle of a tapeworm infecting great white sharks. J. Parasitol. 2011, 97, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Ruhnke, T.R.; Daniel, V.; Jensen, K. Four new species of Paraorygmatobothrium (Eucestoda: Phyllobothriidea) from sharks of the Gulf of Mexico and the Atlantic Ocean, with comments on their host specificity. J. Parasitol. 2020, 106, 133–156. [Google Scholar] [CrossRef] [PubMed]
- Littlewood, D.T.J.; Curini-Galletti, M.; Herniou, E.A. The interrelationships of Proseriata (Platyhelminthes: Seriata) tested with molecules and morphology. Mol. Phylogenetics Evol. 2000, 16, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Olson, P.D.; Littlewood, D.T.J.; Bray, R.A.; Mariaux, J. Interrelationships and evolution of the tapeworms (Platyhelminthes: Cestoda). Mol. Phylogenetics Evol. 2001, 19, 443–467. [Google Scholar] [CrossRef] [Green Version]
- Waeschenbach, A.; Webster, B.L.; Bray, R.A.; Littlewood, D.T.J. Added resolution among ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with complete small and large subunit nuclear ribosomal RNA genes. Mol. Phylogenetics Evol. 2007, 45, 311–325. [Google Scholar] [CrossRef]
- Trevisan, B.; Primon, J.F.; Marques, F.P. Systematics and diversification of Anindobothrium Marques, Brooks & Lasso, 2001 (Eucestoda: Rhinebothriidea). PLoS ONE 2017, 12, e0184632. [Google Scholar]
- Van Steenkiste, N.; Locke, S.A.; Castelin, M.; Marcogliese, D.J.; Abbott, C.L. New primers for DNA barcoding of digeneans and cestodes (Platyhelminthes). Mol. Ecol. Resour. 2015, 15, 945–952. [Google Scholar] [CrossRef]
- Ruhnke, T.R. A new species of Clistobothrium (Cestoda: Tetraphyllidea), with an evaluation of the systematic status of the genus. J. Parasitol. 1993, 79, 37–43. [Google Scholar] [CrossRef]
- Kuris, A.M.; Jaramillo, A.G.; McLaughlin, J.P.; Weinstein, S.B.; Garcia-Vedrenne, A.E.; Poinar, G.O.; Pickering, M.; Steinauer, M.L.; Espinoza, M.; Ashford, J.E.; et al. Monsters of the sea serpent: Parasites of an oarfish, Regalecus russellii. J. Parasitol. 2015, 101, 41–45. [Google Scholar] [CrossRef]
- Ruhnke, T.R. A Monograph on the Phyllobothriidae (Platyhelminthes, Cestoda). Bull. Univ. Neb. State Museum 2011, 25, 1–208. Available online: https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1041&context=museumbulletin (accessed on 13 June 2020).
- Dailey, M.D.; Vogelbein, W. Clistobothrium carcharodoni gen. et sp. n. (Cestoda: Tetraphyllidea) from the spiral valve of the great white shark (Carcharodon carcharias). Proc. Helminthol. Soc. Wash. 1990, 57, 108–112. [Google Scholar]
- Penadés-Suay, J.; Tomás, J.; Merchán, M.; Aznar, F.J. Intestinal helminth fauna of the shortfin mako Isurus oxyrinchus (Elasmobranchii: Lamnidae) in the northeast Atlantic Ocean. Dis. Aquat. Organ. 2017, 123, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arkhipkin, A.I.; Rodhouse, P.G.K.; Pierce, G.J.; Sauer, W.; Sakai, M.; Allcock, L.; Arguelles, J.; Bower, J.R.; Castillo, G.; Ceriola, L. World Squid Fisheries. Rev. Fish. Sci. Aquac. 2015, 23, 92–252. [Google Scholar] [CrossRef] [Green Version]
- Giarratana, F.; Ziino, G.; D’Andrea, V.; Panebianco, A.; Giuffrida, A. Quality assessment of Zeus faber (Peter’s fish) ovaries regularly commercialized for human consumption. Ital. J. Food Saf. 2018, 7, 28–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, H.; Abollo, E.; González, Á.F.; Pascual, S. Scoring the parasite risk in highly-valuable fish species from southern ICES areas. Fish. Res. 2018, 202, 134–139. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 2074/2005 of 5 December 2005 laying down implementing measures for certain products under Regulation (EC) No 853/2004 of the European Parliament and of the Council and for the organisation of official controls under Regulation (EC) No 854/2004 of the European Parliament and of the Council and Regulation (EC) No 882/2004 of the European Parliament and of the Council, derogating from Regulation (EC) No 852/2004 of the European Parliament and of the Council and amending Regulations (EC) No 853/2004 and (EC) No 854/2004. Off. J. Eur. Communities 2005, 338, 27–59.
- Agustí, C.; Aznar, F.J.; Raga, J.A. Tetraphyllidean plerocercoids from western Mediterranean cetaceans and other marine mammals around the world: A comprehensive morphological analysis. J. Parasitol. 2005, 91, 83–92. [Google Scholar] [CrossRef]
- Williams, H.H. The taxonomy, ecology and host specificity of some Phyllobothriidae (Cestoda: Tetraphyllidea), a critical revision of Phyllobothrium Beneden, 1849 and comments on some allied genera. Philos. Trans. R. Soc. B 1968, 253, 231–307. [Google Scholar]
- Guardone, L.; Malandra, R.; Costanzo, F.; Castigliego, L.; Tinacci, L.; Gianfaldoni, D.; Guidi, A.; Armani, A. Assessment of a sampling plan based on visual inspection for the detection of anisakid larvae in fresh anchovies (Engraulis encrasicolus). A first step towards official validation? Food Anal. Methods 2016, 9, 1418–1427. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- WoRMS. Phyllobothriidae Braun, 1900. 2020. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=104946 (accessed on 29 May 2020).
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. | Samp. Period | Geographical Area | Cephalopod Common Name (Scientific Name), n of Examined Specimens | Species (Family, Order) | Localization | P (%) | Parasite ID |
|---|---|---|---|---|---|---|---|
| Guillén-Hernández et al., (2018) [4] | August, 2009–June, 2010 | Yucatán Peninsula, Mexico (FAO 31) | Mexican four-eyed octopus (Octopus maya), 1202 | Prochristianella sp. (Eutetrarhynchidae, Trypanorhynca) | buccal mass, oesophagus, cecum, intestine | 57.0–98.0 | Morph. |
| Eutetrarhynchus sp. (Eutetrarhynchidae, Trypanorhynca) | digestive gland, esophagus, intestine, ink sac | 7.0–59.1 | |||||
| Nybelinia sp. (Tentaculariidae, Trypanorhynca) | buccal mass, esophagus, intestine | 0.4–51.2 | |||||
| Echeneibothrium sp. (Echeinebothriidae, Rhinebothriidea) | cecum, intestine | 4.0–21.0 | |||||
| Prosobothrium sp. (Prosobothriidea, Onchoproteocephalidea) | digestive gland, ink sac | 16.8–27.0 | |||||
| Tetraphyllidea | cecum, intestine | 1.0–7.0 | |||||
| Unidentified plerocercoid | digestive gland, ink sac, gills | 10–26.6 | |||||
| Cavaleiro, (2013) [15] | 2010 | Matosinhos, Portugal, NE Atlantic (FAO 27) | common octopus (Octopus vulgaris), 120 | Nybelinia sp. (Tentaculariidae, Trypanorhynca) | stomach, intestine | 4.2 | Morph. |
| Petrić et al., (2011) [16] | October, 2007–October, 2008 | Central Adriatic Sea (FAO 37.2.1) | shortfin squid (Illex coindetti), 439 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | stomach | 2.3 | Morph. |
| Pardo-Gandarillas et al., (2009) [17] | July, 2003– February, 2004 | Central-Southern Chile (FAO 87) | jumbo flying squid (Dosidicus gigas), 124 | Hepatoxylon trichiuri (Sphyriocephalidae, Trypanorhynca) | mantle cavity, gonads, stomach | 70.2 | Morph. |
| Tentacularia coryphaenae (Tentaculariidae, Trypanorhynca) | mantle cavity, gonads | 5.6 | |||||
| Plerocercoid larvae (Tetraphyllidea) | stomach, cecum and intestine | 83.1 | |||||
| Pelichnibothrium speciosum (Phyllobothriidae, Phyllobothriidea) | intestine | NR | |||||
| Nigmatullin et al., (2009) [18] | 1981–1984 | south part of the eastern Pacific (FAO 87) | neon flying squid (Ommastrephes bartramii),60 | Tentacularia coryphaenae (Tentaculariidae, Trypanorhynca) | whole mantle cavity | 9.1 | Morph. |
| Scyphophyllidium sp. (Phyllobothriidae, Phyllobothriidea) | stomach cavity and cecum | 4.5 | |||||
| Brickle et al., (2001) [3] | February, 1999–June, 2000 | Falkland Islands (South Atlantic Ocean) (FAO 41.3.2) | longfin Patagonian squid (Doryteuthis gahi), 1096 | Clistobothrium montaukensis (Phyllobothriidae, Phyllobothriidea) | cecum, intestine, stomach, mantle | 5.75 | Molec. |
| Ceratobothrium xanthocephalum (Gastrolecithidae, Tetraphyllidea) | |||||||
| Shukhgalter and Nigmatullin, (2001) [19] | 1981–1989 | East Pacific Ocean (FAO 77 and 87) | jumbo squid (Dosidicus gigas), 849 | Pelichnibothrium speciosum (Phyllobothriidae, Phyllobothriidea) | rectum, cecum, stomach | 75.2 | Morph. |
| Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | rectum, cecum, stomach, mantle cavity, buccal cone | 1.2 | |||||
| Tentacularia coryphaenae (Tentaculariidae, Trypanorhynca) | mantle cavity, mantle | 6.6 | |||||
| Gonzalez and Kroeck, (2000) [20] | July–November, 1993 | South West Atlantic St. Matias gulf (FAO 41.3.1) | Argentine short-fin squid (Illex argentinus), 91 | Prosobothrium sp. (Prosobothriidea, Onchoproteocephalidea) | viscera | 100.0 | Morph. |
| Onchobotriidae (Onchoproteocephalidea) | 0.0–100.0 | ||||||
| Nybelinia linguaris (Tentaculariidae, Trypanorhynca) | 0.0–21.0 | ||||||
| Gestal et al., (1998) [21] | December, 1994–December, 1995 | Galician coast, Spain (FAO area 27.9) | Common octopus (Octopus vulgaris), 100 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | Intestine, cecum | 3.0 | Morph. |
| Nybelinia lingualis (Tentaculariidae, Trypanorhynca) | Mouth, stomach, cecum | 3.0 | |||||
| Pascual et al., (1996) [22] | 1992–1995 | coast of Galicia, Spain (FAO 27.9) | broadtailed short-fin squid (Illex coindetti), 600 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | NR | 48.0 | Morph. |
| Pelichnibothrium speciosum (Phyllobothriidae, Phyllobothriidea) | 0.3 | ||||||
| Dinobothrium sp. (Gastrolecithidae, Tetraphyllidea) | 1.0 | ||||||
| Nybelinia yamagutii (Tentaculariidae, Trypanorhynca) | 0.7 | ||||||
| European squid (Loligo vulgaris), 8 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 62.5 | |||||
| European flying squid (Todarodes sagittatus), 65 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 20.0 | |||||
| lesser flying squid (Todaropsis eblanae), 600 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 31.2 | |||||
| Pelichnibothrium speciosum (Phyllobothriidae, Phyllobothriidea) | 0.7 | ||||||
| Nybelinia linguaris (Tentaculariidae, Trypanorhynca) | 0.3 | ||||||
| common cuttlefish (Sepia officinalis), 38 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 2.6 | |||||
| pink cuttlefish (Sepia orbignyana), 22 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 9.0 | |||||
| common octopus (Octopus vulgaris), 70 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 4.3 | |||||
| lesser octopus (Eledone cirrhosa), 67 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | 10.4 | |||||
| Pascual et al., (1994) [23] | October, 1991–April, 1992 | North Galician Shelf waters (FAO 27.8) | broadtailed short-fin squid (Illex coindetti), 70 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | cecum, stomach | 87 | Morph. |
| Dinobothrium septaria (Gastrolecithidae, Tetraphyllidea) | |||||||
| Bower and Margolis (1991) [24] | Summer 1987 | West coast of North America (FAO 21) | flying squid (Ommastrephes bartramii), 68 | Phyllobothrium sp. (Phyllobothriidae, Phyllobothriidea) | esophagus, stomach, cecum, intestine, rectum, gills, gonads | 94.1 | Morph. |
| Tentacularia sp. (Tentaculariidae, Trypanorhynca) | NR | 1.5 | |||||
| Rhadinorhynchus sp. (Rhadinorynchidae, Echinorynchida) | NR | NR |
| Host Sample Code | Total Weight | Total Length | Mantle Lengths | Viscera Weight | Mantle Weight | N Plerocercoid Larvae |
|---|---|---|---|---|---|---|
| D. PEA-23 | 117 | 57 | 22 | 8 | 109 | 1 |
| D. PEA-30 | 87 | 36 | 12 | 18 | 45 | 1 |
| D. PEA-33 | 141 | 41 | 12 | 6 | 92 | 2 |
| D. PEA-34 | 136 | 44 | 12 | 20 | 70 | 1 |
| D. PEA-35 | 153 | 52 | 23 | 26 | 94 | 1 |
| D. PEA-45 | 175 | 43 | 15 | 15 | 79 | 2 |
| D. PEA-58 | 119 | 36 | 24 | 6 | 73 | 3 |
| D. PEA-63 | 163 | 48 | 24 | 17 | 97 | 1 |
| D. PEA-65 | 152 | 46 | 23 | 13 | 89 | 4 |
| D. PEA-67 | 136 | 44 | 22 | 13 | 83 | 2 |
| Overall | 18 | |||||
| Mean (ds) | 137.9 (25.5) | 44.7 (6.5) | 18.9 (5.4) | 14.2 (6.4) | 83.1 (17.8) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guardone, L.; Giusti, A.; Bilska-Zajac, E.; Malandra, R.; Różycki, M.; Armani, A. Molecular Characterization of Clistobothrium sp. Viable Plerocercoids in Fresh Longfin Inshore Squid (Doryteuthis pealeii) and Implications for Cephalopod Inspection. Pathogens 2020, 9, 596. https://doi.org/10.3390/pathogens9070596
Guardone L, Giusti A, Bilska-Zajac E, Malandra R, Różycki M, Armani A. Molecular Characterization of Clistobothrium sp. Viable Plerocercoids in Fresh Longfin Inshore Squid (Doryteuthis pealeii) and Implications for Cephalopod Inspection. Pathogens. 2020; 9(7):596. https://doi.org/10.3390/pathogens9070596
Chicago/Turabian StyleGuardone, Lisa, Alice Giusti, Ewa Bilska-Zajac, Renato Malandra, Miroslaw Różycki, and Andrea Armani. 2020. "Molecular Characterization of Clistobothrium sp. Viable Plerocercoids in Fresh Longfin Inshore Squid (Doryteuthis pealeii) and Implications for Cephalopod Inspection" Pathogens 9, no. 7: 596. https://doi.org/10.3390/pathogens9070596
APA StyleGuardone, L., Giusti, A., Bilska-Zajac, E., Malandra, R., Różycki, M., & Armani, A. (2020). Molecular Characterization of Clistobothrium sp. Viable Plerocercoids in Fresh Longfin Inshore Squid (Doryteuthis pealeii) and Implications for Cephalopod Inspection. Pathogens, 9(7), 596. https://doi.org/10.3390/pathogens9070596