Cyclic Hypoxia Exposure Accelerates the Progression of Amoebic Gill Disease


Abstract
:1. Introduction
2. Results
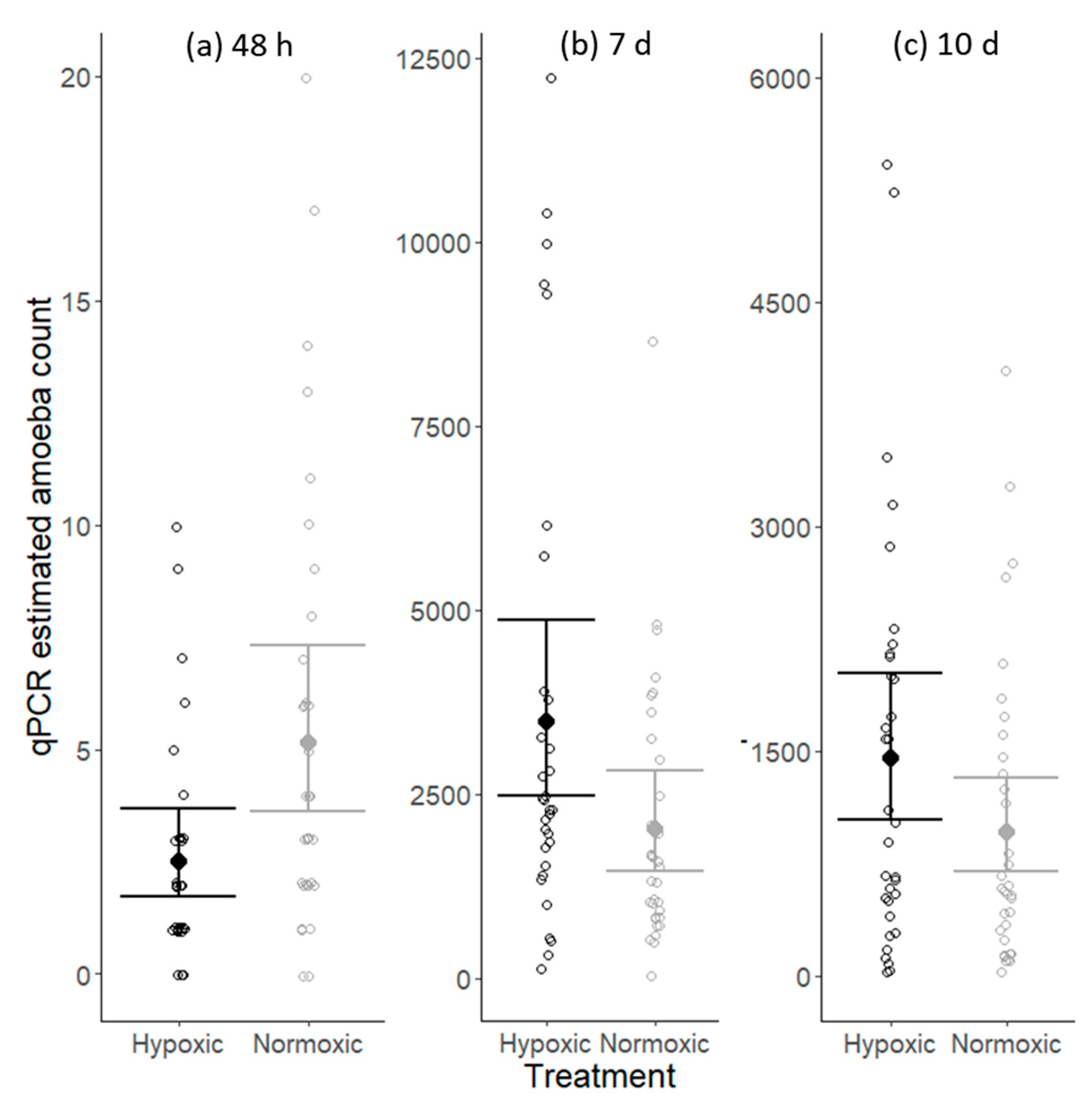
2.1. Amoeba Acquisition and Proliferation
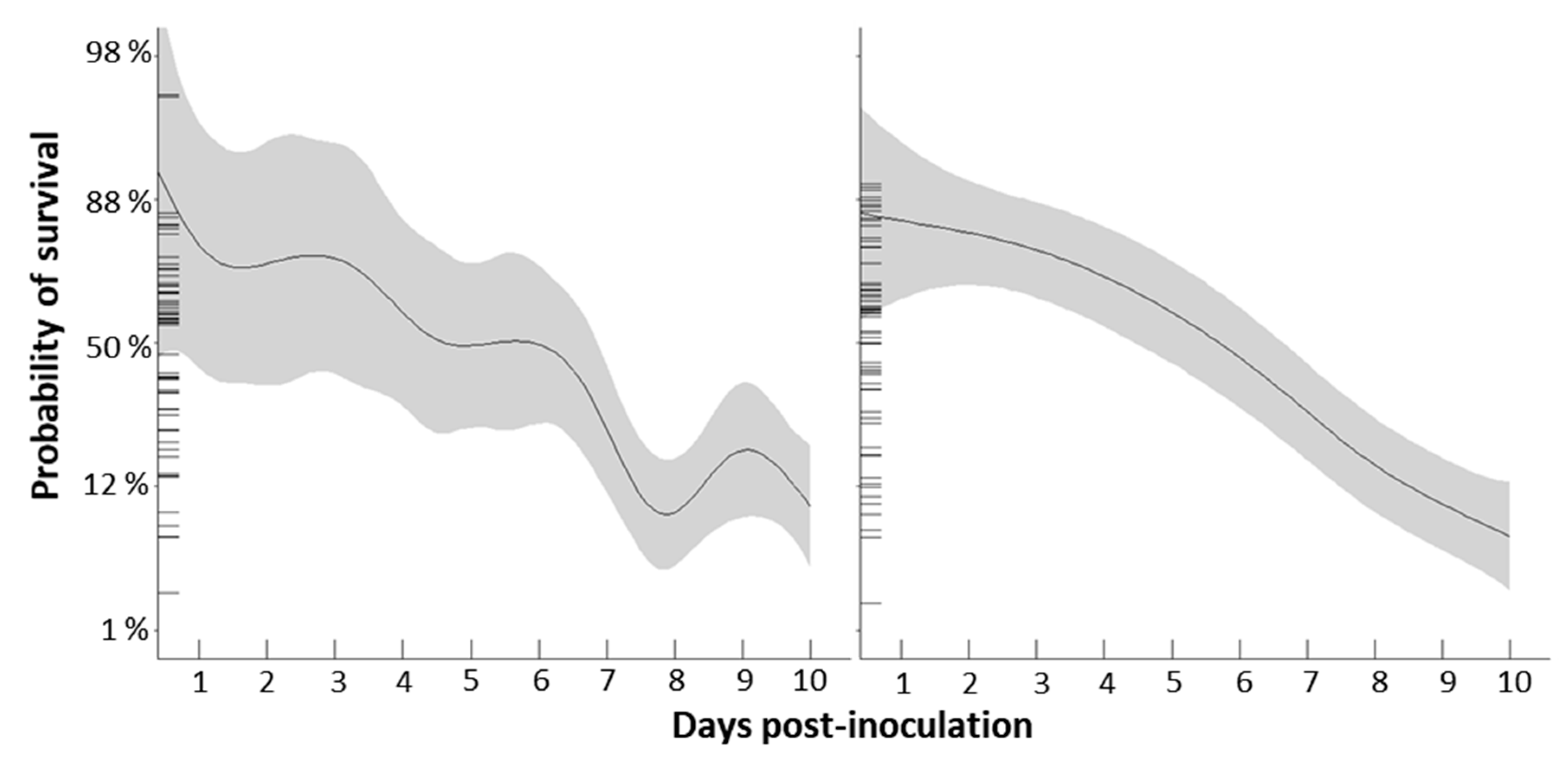
2.2. Mortality
2.3. AGD Progression
2.4. Gross Lesion Morphometry
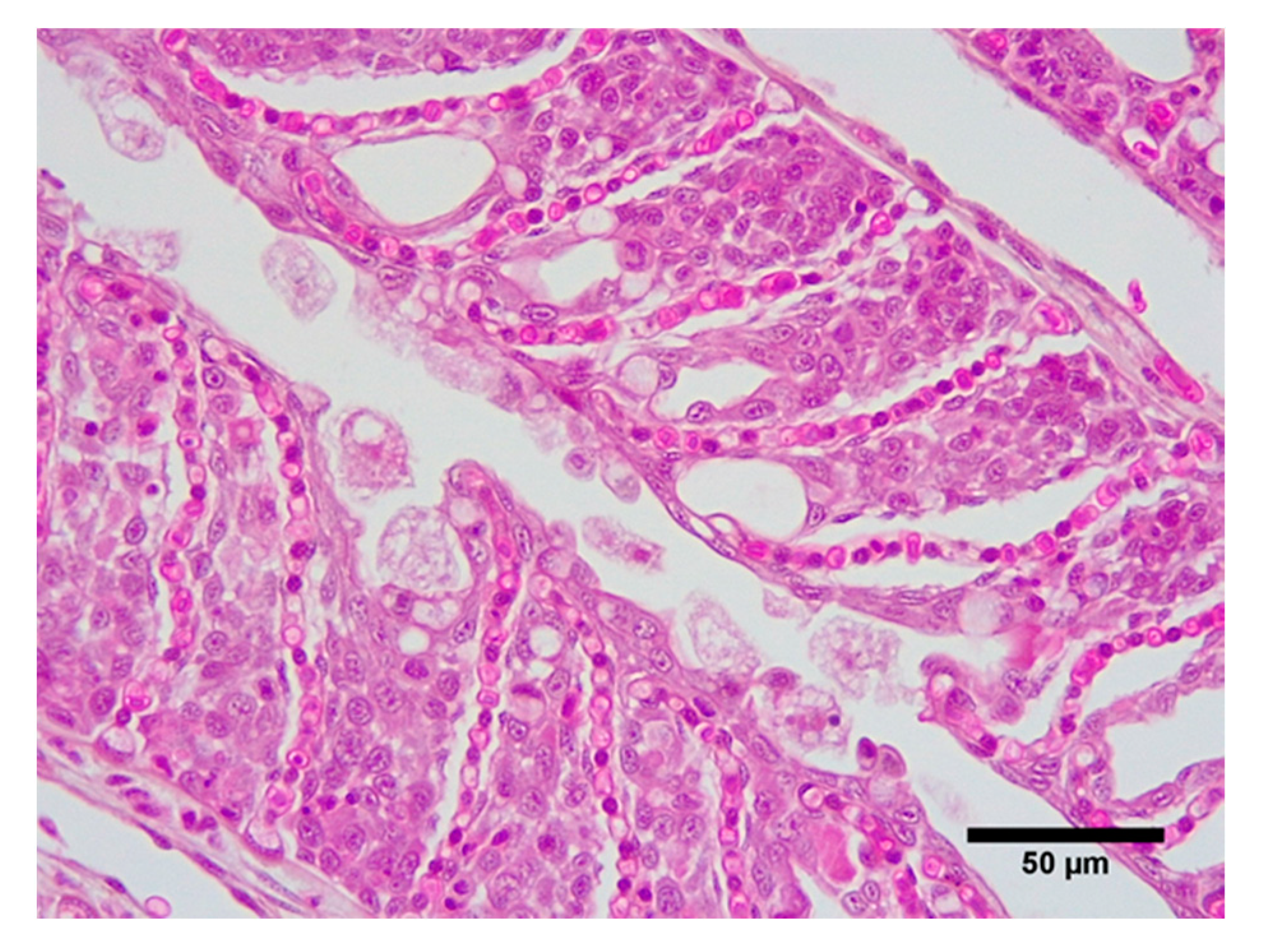
2.5. Histology
3. Discussion.
3.1. Effects of Hypoxia on Amoeba Acquisition
3.2. Effects of Hypoxia on Disease Progression and Prognosis
3.3. Industry Implications
4. Materials and Methods
4.1. Animals & Husbandry
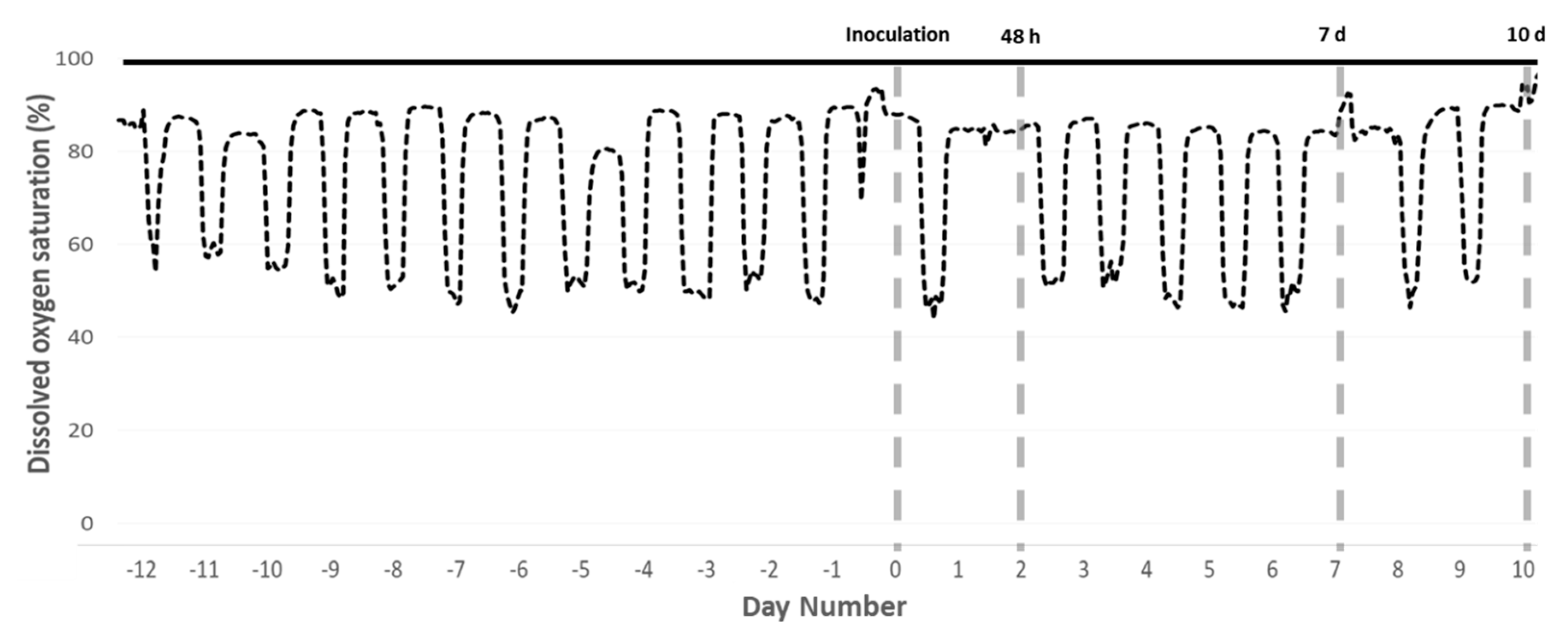
4.2. Hypoxia Induction
4.3. Amoeba Inoculation
4.4. Sampling Protocol
4.5. Gross Lesion Morphometry
4.6. Histological Analysis
4.7. N. Perurans Enumeration
4.8. Data Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burt, K.; Hamoutene, D.; Mabrouk, G.; Lang, C.; Puestow, T.; Drover, D.; Losier, R.; Page, F. Environmental conditions and occurrence of hypoxia within production cages of Atlantic salmon on the south coast of Newfoundland. Aquac. Res. 2012, 43, 607–620. [Google Scholar] [CrossRef]
- Solstorm, D.; Oldham, T.; Solstorm, F.; Klebert, P.; Stien, L.H.; Vågseth, T.; Oppedal, F. Dissolved oxygen variability in a commercial sea-cage exposes farmed Atlantic salmon to growth limiting conditions. Aquaculture 2018, 486, 122–129. [Google Scholar] [CrossRef]
- Stehfest, K.M.; Carter, C.G.; McAllister, J.D.; Ross, J.D.; Semmens, J.M. Response of Atlantic salmon Salmo salar to temperature and dissolved oxygen extremes established using animal-borne environmental sensors. Sci. Rep. 2017, 7, 4545. [Google Scholar] [CrossRef]
- Priede, I.; Solbé, J.D.L.; Nott, J.; O’Grady, K.; Cragg-Hine, D. Behaviour of adult Atlantic salmon, Salmo salar L., in the estuary of the River Ribble in relation to variations in dissolved oxygen and tidal flow. J. Fish Biol. 1988, 33, 133–139. [Google Scholar] [CrossRef]
- Oldham, T.; Dempster, T.; Fosse, J.O.; Oppedal, F. Oxygen gradients affect behavior of caged Atlantic salmon Salmo salar. Aquac. Environ. Interact. 2017, 9, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Johansson, D.; Juell, J.-E.; Oppedal, F.; Stiansen, J.-E.; Ruohonen, K. The influence of the pycnocline and cage resistance on current flow, oxygen flux and swimming behaviour of Atlantic salmon (Salmo salar L.) in production cages. Aquaculture 2007, 265, 271–287. [Google Scholar] [CrossRef]
- Makrinos, D.L.; Bowden, T.J. Natural environmental impacts on teleost immune function. Fish Shellfish Immunol. 2016, 53, 50–57. [Google Scholar] [CrossRef]
- Fry, F. 1 The effect of environmental factors on the physiology of fish. Fish Physiol 1971, 6, 1–98. [Google Scholar]
- Claireaux, G.; Chabot, D. Responses by fishes to environmental hypoxia: Integration through Fry’s concept of aerobic metabolic scope. J. Fish Biol. 2016, 88, 232–251. [Google Scholar] [CrossRef]
- Oldham, T.; Nowak, B.; Hvas, M.; Oppedal, F. Metabolic and functional impacts of hypoxia vary with size in Atlantic salmon. Comp. Biochem. Physiol. Part A Mol Integr. Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Burt, K.; Hamoutene, D.; Perez-Casanova, J.; Kurt Gamperl, A.; Volkoff, H. The effect of intermittent hypoxia on growth, appetite and some aspects of the immune response of Atlantic salmon (Salmo salar). Aquac. Res. 2013, 45, 124–137. [Google Scholar] [CrossRef]
- Kvamme, B.O.; Gadan, K.; Finne-Fridell, F.; Niklasson, L.; Sundh, H.; Sundell, K.; Taranger, G.L.; Evensen, Ø. Modulation of innate immune responses in Atlantic salmon by chronic hypoxia-induced stress. Fish Shellfish Immunol. 2013, 34, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Remen, M.; Oppedal, F.; Torgersen, T.; Imsland, A.K.; Olsen, R.E. Effects of cyclic environmental hypoxia on physiology and feed intake of post-smolt Atlantic salmon: Initial responses and acclimation. Aquaculture 2012, 326, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.J.; Shoemaker, C.A.; Klesius, P.H. Effects of sublethal dissolved oxygen stress on blood glucose and susceptibility to Streptococcus agalactiae in Nile tilapia Oreochromis niloticus. J. Aquat. Anim. Health 2003, 15, 202–208. [Google Scholar] [CrossRef]
- Welker, T.L.; Mcnulty, S.T.; Klesius, P.H. Effect of sublethal hypoxia on the immune response and susceptibility of channel catfish, Ictalurus punctatus, to enteric septicemia. J. World Aquac. Soc. 2007, 38, 12–23. [Google Scholar] [CrossRef]
- Niklasson, L.; Sundh, H.; Olsen, R.-E.; Jutfelt, F.; Skjødt, K.; Nilsen, T.O.; Sundell, K.S. Effects of cortisol on the intestinal mucosal immune response during cohabitant challenge with IPNV in Atlantic salmon (Salmo salar). PLoS ONE 2014, 9, e94288. [Google Scholar] [CrossRef] [Green Version]
- Andersen, L.; Hodneland, K.; Nylund, A. No influence of oxygen levels on pathogenesis and virus shedding in Salmonid alphavirus (SAV)-challenged Atlantic salmon (Salmo salar L.). Virol. J. 2010, 7, 198. [Google Scholar] [CrossRef] [Green Version]
- Lund, M.; Dahle, M.K.; Timmerhaus, G.; Alarcon, M.; Powell, M.; Aspehaug, V.; Rimstad, E.; Jørgensen, S.M. Hypoxia tolerance and responses to hypoxic stress during heart and skeletal muscle inflammation in Atlantic salmon (Salmo salar). PLoS ONE 2017, 12, e0181109. [Google Scholar] [CrossRef]
- Oldham, T.; Rodger, H.; Nowak, B.F. Incidence and distribution of amoebic gill disease (AGD)—An epidemiological review. Aquaculture 2016, 457, 35–42. [Google Scholar] [CrossRef]
- Hvas, M.; Karlsbakk, E.; Mæhle, S.; Wright, D.W.; Oppedal, F. The gill parasite Paramoeba perurans compromises aerobic scope, swimming capacity and ion balance in Atlantic salmon. Conserv. Physiol. 2017, 5, cox066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.; Nowak, B. Distribution and structure of lesions in the gills of Atlantic salmon, Salmo salar L., affected with amoebic gill disease. J. Fish Dis. 2001, 24, 535–542. [Google Scholar] [CrossRef]
- Crosbie, P.; Bridle, A.; Cadoret, K.; Nowak, B. In vitro cultured Neoparamoeba perurans causes amoebic gill disease in Atlantic salmon and fulfils Koch’s postulates. Int. J. Parasitol. 2012, 42, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Borch, K.; Jensen, F.B.; Andersen, B.B. Cardiac activity, ventilation rate and acid-base regulation in rainbow trout exposed to hypoxia and combined hypoxia and hypercapnia. Fish Physiol. Biochem. 1993, 12, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.; Gamperl, A.; Birtwell, I. Prolonged swimming, recovery and repeat swimming performance of mature sockeye salmon Oncorhynchus nerka exposed to moderate hypoxia and pentachlorophenol. J. Exp. Biol. 1998, 201, 2183–2193. [Google Scholar]
- Hvas, M.; Oppedal, F. Physiological responses of farmed Atlantic salmon and two cohabitant species of cleaner fish to progressive hypoxia. Aquaculture 2019, 512, 734353. [Google Scholar] [CrossRef]
- Mikheev, V.N.; Pasternak, A.F.; Valtonen, E.; Taskinen, J. Increased ventilation by fish leads to a higher risk of parasitism. Parasites Vectors 2014, 7, 281. [Google Scholar] [CrossRef] [Green Version]
- Crosbie, P.B.; Bridle, A.R.; Leef, M.J.; Nowak, B.F. Effects of different batches of Neoparamoeba perurans and fish stocking densities on the severity of amoebic gill disease in experimental infection of Atlantic salmon, Salmo salar L. Aquac. Res. 2010, 41, e505–e516. [Google Scholar] [CrossRef]
- Bridle, A.R.; Davenport, D.L.; Crosbie, P.B.; Polinski, M.; Nowak, B.F. Neoparamoeba perurans loses virulence during clonal culture. Int. J. Parasitol. 2015, 45, 575–578. [Google Scholar] [CrossRef]
- Roberts, S.; Powell, M. Freshwater bathing alters the mucous layer of marine Atlantic salmon Salmo salar L. J. Fish Biol. 2008, 72, 1864–1870. [Google Scholar] [CrossRef]
- Sveen, L.R. Aquaculture Relevant Stressors and Their Impacts on Skin and Wound Healing in Post-Smolt Atlantic Salmon (Salmo salar L.). Ph.D. Thesis, The University of Bergen, Bergen, Norway, November 2018. [Google Scholar]
- Vatsos, I.; Kotzamanis, Y.; Henry, M.; Angelidis, P.; Alexis, M. Monitoring stress in fish by applying image analysis to their skin mucous cells. Eur. J. Histochem. EJH 2010, 54, e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haond, C.; Nolan, D.; Ruane, N.; Rotllant, J.; Bonga, S.W. Cortisol influences the host-parasite interaction between the rainbow trout (Oncorhynchus mykiss) and the crustacean ectoparasite Argulus japonicus. Parasitology 2003, 127, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Munday, B.; Foster, C.; Roubal, F.; Lester, R.; Perkins, F.; Cheng, T. Paramoebic gill infection and associated pathology of Atlantic salmon, Salmo salar and rainbow trout, Salmo gairdneri in Tasmania. In Proceedings of the Third International Colloquium on Pathology in Marine Aquaculture, Gloucester Point, VA, USA, 2–6 October 1988; pp. 215–222. [Google Scholar]
- Fisk, D.; Powell, M.; Nowak, B. The effect of amoebic gill disease and hypoxia on survival and metabolic rate of Atlantic salmon (Salmo salar). Bull. Eur. Assoc. Fish Pathol. 2002, 22, 190–194. [Google Scholar]
- Powell, M.; Nowak, B. Acid-base and respiratory effects of confinement in Atlantic salmon affected with amoebic gill disease. J. Fish Biol. 2003, 62, 51–63. [Google Scholar] [CrossRef]
- Richards, J.G.; Farrell, A.P.; Brauner, C.J. Hypoxia; Academic Press: Cambridge, MA, USA, 2009; Volume 27. [Google Scholar]
- Hing, S.; Narayan, E.J.; Thompson, R.A.; Godfrey, S.S. The relationship between physiological stress and wildlife disease: Consequences for health and conservation. Wildl. Res. 2016, 43, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Cecchini, S.; Saroglia, M. Antibody response in sea bass (Dicentrarchus labrax L.) in relation to water temperature and oxygenation. Aquac. Res. 2002, 33, 607–613. [Google Scholar] [CrossRef]
- Fukuda, Y.; Maita, M.; Satoh, K.; Okamoto, N. Influence of Dissolved Oxygen Concentration on the Mortality of Yellowtail Experimentally Infected with Enterococcus seriolicida. Fish Pathol. 1997, 32, 129–130. [Google Scholar] [CrossRef]
- Steinum, T.; Kvellestad, A.; Rønneberg, L.; Nilsen, H.; Asheim, A.; Fjell, K.; Nygård, S.; Olsen, A.; Dale, O. First cases of amoebic gill disease (AGD) in Norwegian seawater farmed Atlantic salmon, Salmo salar L., and phylogeny of the causative amoeba using 18S cDNA sequences. J. Fish Dis. 2008, 31, 205–214. [Google Scholar] [CrossRef]
- Roberts, R.; Pearson, M. Infectious pancreatic necrosis in Atlantic salmon, Salmo salar L. J. Fish Dis. 2005, 28, 383–390. [Google Scholar] [CrossRef]
- Johansson, D.; Ruohonen, K.; Kiessling, A.; Oppedal, F.; Stiansen, J.-E.; Kelly, M.; Juell, J.-E. Effect of environmental factors on swimming depth preferences of Atlantic salmon (Salmo salar L.) and temporal and spatial variations in oxygen levels in sea cages at a fjord site. Aquaculture 2006, 254, 594–605. [Google Scholar] [CrossRef]
- Rodger, H.D.; Henry, L.; Mitchell, S.O. Non-infectious gill disorders of marine salmonid fish. Rev. Fish Biol. Fish. 2011, 21, 423–440. [Google Scholar] [CrossRef]
- Fitridge, I.; Dempster, T.; Guenther, J.; de Nys, R. The impact and control of biofouling in marine aquaculture: A review. Biofouling 2012, 28, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Bloecher, N.; Powell, M.; Hytterød, S.; Gjessing, M.; Wiik-Nielsen, J.; Mohammad, S.N.; Johansen, J.; Hansen, H.; Floerl, O.; Gjevre, A.-G. Effects of cnidarian biofouling on salmon gill health and development of amoebic gill disease. PLoS ONE 2018, 13, e0199842. [Google Scholar] [CrossRef] [Green Version]
- Oliver, E.C.; Benthuysen, J.A.; Bindoff, N.L.; Hobday, A.J.; Holbrook, N.J.; Mundy, C.N.; Perkins-Kirkpatrick, S.E. The unprecedented 2015/16 Tasman Sea marine heatwave. Nat. Commun. 2017, 8, 16101. [Google Scholar] [CrossRef] [Green Version]
- Oldham, T.; Oppedal, F.; Dempster, T. Cage size affects dissolved oxygen distribution in salmon aquaculture. Aquac. Environ. Interact. 2018, 10, 149–156. [Google Scholar] [CrossRef]
- Mitchell, S.O.; Rodger, H.D. A review of infectious gill disease in marine salmonid fish. J. Fish Dis. 2011, 34, 411–432. [Google Scholar] [CrossRef]
- Herrero, A.; Thompson, K.; Ashby, A.; Rodger, H.; Dagleish, M. Complex Gill Disease: An Emerging Syndrome in Farmed Atlantic Salmon (Salmo salar L.). J. Comp. Pathol. 2018, 163, 23–28. [Google Scholar] [CrossRef]
- Stien, L.H.; Nilsson, J.; Hevrøy, E.M.; Oppedal, F.; Kristiansen, T.S.; Lien, A.M.; Folkedal, O. Skirt around a salmon sea cage to reduce infestation of salmon lice resulted in low oxygen levels. Aquac. Eng. 2012, 51, 21–25. [Google Scholar] [CrossRef]
- Wright, D.; Stien, L.; Dempster, T.; Vågseth, T.; Nola, V.; Fosseidengen, J.-E.; Oppedal, F. “Snorkel”lice barrier technology reduced two co-occurring parasites, the salmon louse (Lepeophtheirus salmonis) and the amoebic gill disease causing agent (Neoparamoeba perurans), in commercial salmon sea-cages. Prev. Vet. Med. 2017, 140, 97–105. [Google Scholar] [CrossRef]
- Oppedal, F.; Samsing, F.; Dempster, T.; Wright, D.W.; Bui, S.; Stien, L.H. Sea lice infestation levels decrease with deeper “snorkel” barriers in Atlantic salmon sea-cages. Pest Manag. Sci. 2017, 73, 1935–1943. [Google Scholar] [CrossRef] [PubMed]
- Dempster, T.; Wright, D.; Oppedal, F. Identifying the Nature, Extent and Duration of Critical Production Periods for Atlantic Salmon in Macquarie Harbour, Tasmania, During Summer; Fisheries Research and Development Corporation: Canberra, Australia, 2016; ISBN 9780734053022. [Google Scholar]
- Morrison, R.; Crosbie, P.; Nowak, B. The induction of laboratory-based amoebic gill disease revisited. J. Fish Dis. 2004, 27, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W.; Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; et al. NIH image to imageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bridle, A.; Crosbie, P.; Cadoret, K.; Nowak, B. Rapid detection and quantification of Neoparamoeba perurans in the marine environment. Aquaculture 2010, 309, 56–61. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. Error | z-Value | p-Value | |
|---|---|---|---|---|
| Intercept | 0.920 | 0.194 | 4.736 | <0.001 |
| Treatment_Normoxic | 0.718 | 0.264 | 2.721 | <0.01 |
| 7 days post-inoculation | 7.232 | 0.240 | 30.102 | <0.001 |
| 10 days post-inoculation | 6.361 | 0.238 | 26.659 | <0.001 |
| Treatment_Normoxic: 7 days | −1.258 | 0.328 | −3.826 | <0.001 |
| Treatment_Normoxic: 10 days | −1.133 | 0.325 | −3.482 | <0.001 |
| Estimate | edf | Chi.sq | p-Value | |
|---|---|---|---|---|
| Intercept | 3.7609 | |||
| Day: Hypoxic | 6.478 | 54.666 | <0.001 | |
| Day: Normoxic | 2.593 | 57.605 | <0.001 | |
| Tank | 2.074 | 3.858 | 0.103 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oldham, T.; Dempster, T.; Crosbie, P.; Adams, M.; Nowak, B. Cyclic Hypoxia Exposure Accelerates the Progression of Amoebic Gill Disease. Pathogens 2020, 9, 597. https://doi.org/10.3390/pathogens9080597
Oldham T, Dempster T, Crosbie P, Adams M, Nowak B. Cyclic Hypoxia Exposure Accelerates the Progression of Amoebic Gill Disease. Pathogens. 2020; 9(8):597. https://doi.org/10.3390/pathogens9080597
Chicago/Turabian StyleOldham, Tina, Tim Dempster, Philip Crosbie, Mark Adams, and Barbara Nowak. 2020. "Cyclic Hypoxia Exposure Accelerates the Progression of Amoebic Gill Disease" Pathogens 9, no. 8: 597. https://doi.org/10.3390/pathogens9080597
APA StyleOldham, T., Dempster, T., Crosbie, P., Adams, M., & Nowak, B. (2020). Cyclic Hypoxia Exposure Accelerates the Progression of Amoebic Gill Disease. Pathogens, 9(8), 597. https://doi.org/10.3390/pathogens9080597