Immune Control of Herpesvirus Infection in Molluscs



{kind=link}
Abstract
:1. Introduction
2. Herpesvirus Infection in Molluscs
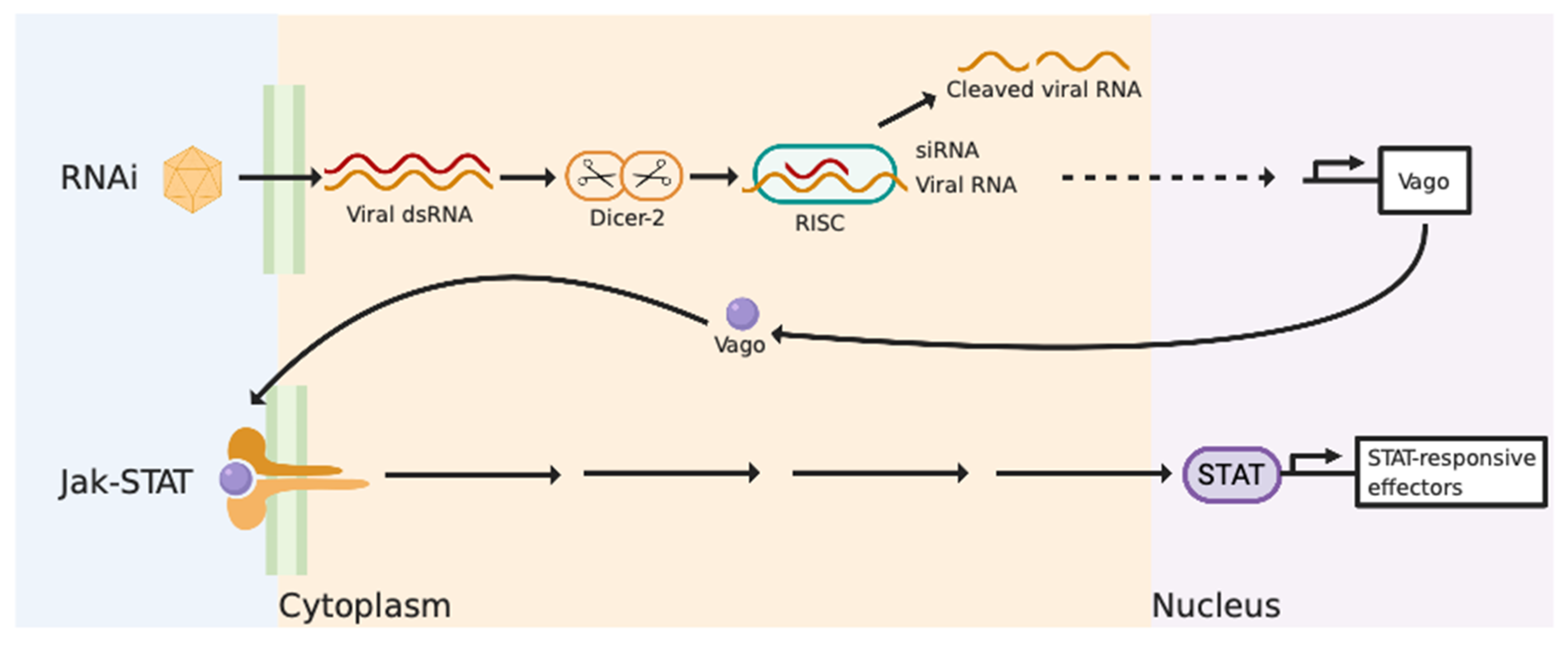
3. Anti-Viral Immunity
4. Immune Based Strategies for Control of Herpesvirus in Molluscs
5. Future Directions for Control of Herpesvirus in Molluscs
Author Contributions
Funding
Conflicts of Interest
References
- Mobsby, D.; Steven, H.A.; Curtotti, R. Australian Fisheries and Aquaculture Outlook 2020; Department of Agriculture WatE ABARES: Canberra, Australia, 2020; p. 9.
- Mobsby, D.; Curtotti, R. Fisheries Outlook to 2023–2024; ABARES: Canberra, Australia, 2019; 9, pp. 87–95.
- Renault, T. Chapter 37-Malacoherpesviruses of Mollusks. In Aquaculture Virology; Kibenge, F.S.B., Godoy, M.G., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 513–524. [Google Scholar]
- Cook, P.A. Recent Trends in Worldwide Abalone Production. J. Shellfish Res. 2016, 35, 581–583. [Google Scholar] [CrossRef]
- Mobsby, D.; Curtotti, R. Fisheries; Agricultural Commodities: Canberra, Australia, 2019; p. 87. [Google Scholar]
- Green, T.J.; Helbig, K.; Speck, P.; Raftos, D.A. Primed for success: Oyster parents treated with poly(I:C) produce offspring with enhanced protection against Ostreid herpesvirus type I infection. Mol. Immunol. 2016, 78, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Green, T.J.; Rolland, J.-L.; Vergnes, A.; Raftos, D.; Montagnani, C. OsHV-1 countermeasures to the Pacific oyster’s anti-viral response. Fish Shellfish Immunol. 2015, 47, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woźniakowski, G.; Samorek-Salamonowicz, E. Animal herpesviruses and their zoonotic potential for cross-species infection. Ann. Agric. Environ. Med. 2015, 22, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.-Y.; Huang, S.; Yao, T.; Gao, F.; Jiang, J.-Z.; Wang, J.-Y. Detection of viruses in abalone tissue using metagenomics technology. Aquac. Res. 2018, 49, 2704–2713. [Google Scholar] [CrossRef]
- Friedman, C.S.; Estes, R.M.; Stokes, N.A.; Burge, C.A.; Hargove, J.S.; Barber, B.J.; Elston, R.A.; Burreson, E.M.; Reece, K.S. Herpes virus in juvenile Pacific oysters Crassostrea gigas from Tomales Bay, California, coincides with summer mortality episodes. Dis. Aquat. Organ. 2005, 63, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clegg, T.A.; Morrissey, T.; Geoghegan, F.; Martin, S.W.; Lyons, K.; Ashe, S.; More, S.J. Risk factors associated with increased mortality of farmed Pacific oysters in Ireland during 2011. Prev. Vet. Med. 2014, 113, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Hick, P.; Gabor, M.; Spiers, Z.; Fell, S.A.; Gu, X.; Read, A.; Go, J.; Dove, M.; Connor, W.O. Identification and characterisation of an ostreid herpesvirus-1 microvariant (OsHV-1 µ-var) in Crassostrea gigas (Pacific oysters) in Australia. Dis. Aquat. Org. 2013, 105, 109–126. [Google Scholar] [CrossRef] [Green Version]
- Bai, C.; Wang, C.; Xia, J.; Sun, H.; Zhang, S.; Huang, J. Emerging and endemic types of Ostreid herpesvirus 1 were detected in bivalves in China. J. Invert. Pathol. 2015, 124, 98–106. [Google Scholar] [CrossRef]
- Hooper, C.; Hardy-Smith, P.; Handlinger, J. Ganglioneuritis causing high mortalities in farmed Australian abalone (Haliotis laevigata and Haliotis rubra). Aust. Vet. J. 2007, 85, 188–193. [Google Scholar] [CrossRef]
- Chang, P.H.; Kuo, S.T.; Lai, S.H.; Yang, H.S.; Ting, Y.Y.; Hsu, C.L.; Chen, H.C. Herpes-like virus infection causing mortality of cultured abalone Haliotis diversicolor supertexta in Taiwan. Dis. Aquat. Org. 2005, 65, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul-Pont, I.; Evans, O.; Dhand, N.K.; Rubio, A.; Coad, P.; Whittington, R.J. Descriptive epidemiology of mass mortality due to Ostreid herpesvirus-1 (OsHV-1) in commercially farmed Pacific oysters (Crassostrea gigas) in the Hawkesbury River estuary, Australia. Aquaculture 2014, 422–423, 146–159. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, N.; Gairin, I.; Pérez, J.; Andree, K.B.; Roque, A.; Fernández-Tejedor, M.; Rodgers, C.J.; Aguilera, C.; Furones, M.D. A Production Calendar Based on Water Temperature, Spat Size, and Husbandry Practices Reduce OsHV-1 μvar Impact on Cultured Pacific Oyster Crassostrea gigas in the Ebro Delta (Catalonia), Mediterranean Coast of Spain. Front. Physiol. 2017, 8, 125. [Google Scholar] [CrossRef] [Green Version]
- Agriculture, D.O. Disease strategy: Abalone viral ganglioneuritis (Version 1.0). In Australian Aquatic Veterinary Emergeny Plan (AQUAVETPLAN); Australian Government Department of Agriculture: Canberra, Australia, 2014. [Google Scholar]
- Segarra, A.; Pépin, J.F.; Arzul, I.; Morga, B.; Faury, N.; Renault, T. Detection and description of a particular Ostreid herpesvirus 1 genotype associated with massive mortality outbreaks of Pacific oysters, Crassostrea gigas, in France in 2008. Virus Res. 2010, 153, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Corbeil, S.; Colling, A.; Williams, L.M.; Wong, F.Y.; Savin, K.; Warner, S.; Murdoch, B.; Cogan, N.O.; Sawbridge, T.I.; Fegan, M. Development and validation of a TaqMan® PCR assay for the Australian abalone herpes-like virus. Dis. Aquat. Org. 2010, 92, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.J.; Trus, B.L.; Cheng, N.; Steven, A.C.; Watson, M.S.; Cunningham, C.; Deuff, R.-M.L.; Renault, T. A novel class of herpesvirus with bivalve hosts. J. Gen. Virol. 2005, 86, 41–53. [Google Scholar] [CrossRef]
- Farley, C.A.; Banfield, W.G.; Kasnic, G., Jr.; Foster, W.S. Oyster herpes-type virus. Science (New York, N.Y.) 1972, 178, 759–760. [Google Scholar] [CrossRef]
- Bai, C.-M.; Rosani, U.; Li, Y.-N.; Zhang, S.-M.; Xin, L.-S.; Wang, C.-M. RNA-seq of HaHV-1-infected abalones reveals a common transcriptional signature of Malacoherpesviruses. Sci. Rep. 2019, 9, 938. [Google Scholar] [CrossRef] [Green Version]
- Rosani, U.; Venier, P. Oyster RNA-seq Data Support the Development of Malacoherpesviridae Genomics. Front. Microbiol. 2017, 8, 1515. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Guo, Z.; Feng, J.; Liu, G.; Xu, L.; Chen, B.; Pan, J. Virus infection in cultured abalone, Haliotis diversicolor Reeve in Guangdong Province, China. J. Shellfish Res. 2004, 23, 1163–1169. [Google Scholar]
- Bai, C.-M.; Li, Y.-N.; Chang, P.-H.; Jiang, J.-Z.; Xin, L.-S.; Li, C.; Wang, J.-Y.; Wang, C.-M. Susceptibility of two abalone species, Haliotis diversicolor supertexta and Haliotis discus hannai, to Haliotid herpesvirus 1 infection. J. Invertebr. Pathol. 2019, 160, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Mushegian, A.; Karin, E.L.; Pupko, T. Sequence analysis of malacoherpesvirus proteins: Pan-herpesvirus capsid module and replication enzymes with an ancient connection to “Megavirales”. Virology 2018, 513, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Corbeil, S.; Williams, L.M.; McColl, K.A.; Crane, M.S. Australian abalone (Haliotis laevigata, H. rubra and H. conicopora) are susceptible to infection by multiple abalone herpesvirus genotypes. Dis. Aquat. Org. 2016, 119, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.H.; Kuo, S.T.; Renault, T.; Friedman, C.S.; Chang, P.H. Development of a polymerase chain reaction for the detection of abalone herpesvirus infection based on the DNA polymerase gene. J. Virol. Methods 2012, 185, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-W.; Chang, P.-H.; Chen, M.-S.; Renault, T.; Chen, M.-M.; Kuo, S.-T.; Cheng, C.-H. Exploring the chronic mortality affecting abalones in taiwan: Differentiation of abalone herpesvirus-associated acute infection from chronic mortality by pcr and in situ hybridization and histopathology. Taiwan Vet. J. 2016, 42, 1–9. [Google Scholar] [CrossRef]
- Bai, C.; Gao, W.; Wang, C.; Yu, T.; Zhang, T.; Qiu, Z.; Wang, Q.; Huang, J. Identification and characterization of ostreid herpesvirus 1 associated with massive mortalities of Scapharca broughtonii broodstocks in China. Dis. Aquat. Org. 2016, 118, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Arzul, I.; Renault, T.; Lipart, C.; Davison, A.J. Evidence for interspecies transmission of oyster herpesvirus in marine bivalves. J. Gen. Virol. 2001, 82, 865–870. [Google Scholar] [CrossRef]
- Martenot, C.; Oden, E.; Travaillé, E.; Malas, J.-P.; Houssin, M. Detection of different variants of Ostreid Herpesvirus 1 in the Pacific oyster, Crassostrea gigas between 2008 and 2010. Virus Res. 2011, 160, 25–31. [Google Scholar] [CrossRef]
- Bai, C.-M.; Wang, Q.-C.; Morga, B.; Shi, J.; Wang, C.-M. Experimental infection of adult Scapharca broughtonii with Ostreid herpesvirus SB strain. J. Invertebr. Pathol. 2017, 143, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Ren, W.; Chen, H.; Renault, T.; Cai, Y.; Bai, C.; Wang, C.; Huang, J. Complete genome sequence of acute viral necrosis virus associated with massive mortality outbreaks in the Chinese scallop, Chlamys farreri. Virol. J. 2013, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Evans, O.; Paul-Pont, I.; Whittington, R.J. Detection of ostreid herpesvirus 1 microvariant DNA in aquatic invertebrate species, sediment and other samples collected from the Georges River estuary, New South Wales, Australia. Dis. Aquat. Org. 2017, 122, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Renault, T.; Moreau, P.; Faury, N.; Pepin, J.-F.; Segarra, A.; Webb, S. Analysis of Clinical Ostreid Herpesvirus 1 Specimens by Sequencing Amplified Fragments from Three Virus Genome Areas. J. Virol. 2012, 86, 5942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayfield, S.; McGarvey, R.; Gorfine, H.K.; Peeters, H.; Burch, P.; Sharma, S. Survey estimates of fishable biomass following a mass mortality in an Australian molluscan fishery. J. Fish Dis. 2011, 34, 287–302. [Google Scholar] [CrossRef]
- Bai, C.-M.; Zhang, S.-M.; Li, Y.-N.; Xin, L.-S.; Rosani, U.; Wang, C.-M. Dual Transcriptomic Analysis Reveals a Delayed Antiviral Response of Haliotis diversicolor supertexta against Haliotid Herpesvirus-1. Viruses 2019, 11, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbeil, S.; McColl, K.A.; Williams, L.M.; Slater, J.; Crane, M.S.J. Innate resistance of New Zealand paua to abalone viral ganglioneuritis. J. Invertebr. Pathol. 2017, 146, 31–35. [Google Scholar] [CrossRef]
- OIE. Infection with Ostreid Herpesvirus 1 microvariants. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2019; p. 14. [Google Scholar]
- Schikorski, D.; Faury, N.; Pepin, J.F.; Saulnier, D.; Tourbiez, D.; Renault, T. Experimental ostreid herpesvirus 1 infection of the Pacific oyster Crassostrea gigas: Kinetics of virus DNA detection by q-PCR in seawater and in oyster samples. Virus Res. 2011, 155, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, T.; Bickham, U.; Bayne, C. Molluscan cells in culture: Primary cell cultures and cell lines. Can. J. Zool. 2013, 91, 391–404. [Google Scholar] [CrossRef]
- Spear, P.G.; Longnecker, R. Herpesvirus Entry: An Update. J. Virol. 2003, 77, 10179. [Google Scholar] [CrossRef] [Green Version]
- Spear, P.G. Herpes simplex virus: Receptors and ligands for cell entry. Cell. Microbiol. 2004, 6, 401–410. [Google Scholar] [CrossRef]
- Martenot, C.; Faury, N.; Morga, B.; Degremont, L.; Lamy, J.-B.; Houssin, M.; Renault, T. Exploring First Interactions Between Ostreid Herpesvirus 1 (OsHV-1) and Its Host, Crassostrea gigas: Effects of Specific Antiviral Antibodies and Dextran Sulfate. Front. Microbiol. 2019, 10. Available online: https://www.readcube.com/articles/10.3389/fmicb.2019.01128 (accessed on 20 July 2020). [CrossRef] [Green Version]
- Jouaux, A.; Lafont, M.; Blin, J.-L.; Houssin, M.; Mathieu, M.; Lelong, C. Physiological change under OsHV-1 contamination in Pacific oyster Crassostrea gigas through massive mortality events on fields. BMC Genomics 2013, 14, 590. [Google Scholar] [CrossRef] [Green Version]
- Green, T.J.; Raftos, D.; Speck, P.; Montagnani, C. Antiviral immunity in marine molluscs. J. Gen. Virol. 2015, 96, 2471–2482. [Google Scholar] [CrossRef] [Green Version]
- Lamiable, O.; Imler, J.-L. Induced antiviral innate immunity in Drosophila. Curr. Opin. Microbiol. 2014, 20, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Fragkoudis, R.; Attarzadeh-Yazdi, G.; Nash, A.A.; Fazakerley, J.K.; Kohl, A. Advances in dissecting mosquito innate immune responses to arbovirus infection. J. Gen. Virol. 2009, 90, 2061–2072. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, J. Memory in the innate and adaptive immune systems. Microbes Infect. 2004, 6, 1410–1417. [Google Scholar] [CrossRef]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Green, T.J.; Speck, P.; Geng, L.; Raftos, D.; Beard, M.R.; Helbig, K.J. Oyster viperin retains direct antiviral activity and its transcription occurs via a signalling pathway involving a heat-stable haemolymph protein. J. Gen. Virol. 2015, 96, 3587–3597. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-H.; Weng, S.-P.; He, J.-G. Nucleic acid-induced antiviral immunity in invertebrates: An evolutionary perspective. Dev. Comp. Immunol. 2015, 48, 291–296. [Google Scholar] [CrossRef]
- Green, T.J.; Montagnani, C. Poly I:C induces a protective antiviral immune response in the Pacific oyster (Crassostrea gigas) against subsequent challenge with Ostreid herpesvirus (OsHV-1 μvar). Fish Shellfish Immunol. 2013, 35, 382–388. [Google Scholar] [CrossRef]
- Milutinovic, B.; Kurtz, J. Immune memory in invertebrates. Semin. Immunol. 2016, 28, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect Antiviral Innate Immunity: Pathways, Effectors, and Connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [Green Version]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffmann, J.A.; Imler, J.-L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in drosophila. Nat. Immunol. 2008, 9, 1425. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; He, Y.; Zhang, L.; Lelong, C.; Jouaux, A. Immune and stress responses in oysters with insights on adaptation. Fish Shellfish Immunol. 2015, 46, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, X.; Song, L. The oyster immunity. Dev. Comp. Immunol. 2018, 80, 99–118. [Google Scholar] [CrossRef]
- Guo, X.; Ford, S.E. Infectious diseases of marine molluscs and host responses as revealed by genomic tools. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150206. [Google Scholar] [CrossRef] [Green Version]
- Green, T.J.; Speck, P. Antiviral defense and innate immune memory in the oyster. Viruses 2018, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Jouaux, A.; Ford, S.E.; Lelong, C.; Sourdaine, P.; Mathieu, M.; Guo, X. Transcriptome analysis reveals strong and complex antiviral response in a mollusc. Fish Shellfish Immunol. 2015, 46, 131–144. [Google Scholar] [CrossRef]
- Rosani, U.; Varotto, L.; Domeneghetti, S.; Arcangeli, G.; Pallavicini, A.; Venier, P. Dual analysis of host and pathogen transcriptomes in ostreid herpesvirus 1-positive Crassostrea gigas. Environ. Microbiol. 2015, 17, 4200–4212. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Rosani, U.; Bai, C.M.; Maso, L.; Shapiro, M.; Abbadi, M.; Domeneghetti, S.; Wang, C.M.; Cendron, L.; MacCarthy, T.; Venier, P. A-to-I editing of Malacoherpesviridae RNAs supports the antiviral role of ADAR1 in mollusks. BMC Evol. Biol. 2019, 19, 149. [Google Scholar] [CrossRef] [PubMed]
- Dang, V.T.; Benkendorff, K.; Corbeil, S.; Williams, L.M.; Hoad, J.; Crane, M.S.J.; Speck, P. Immunological changes in response to herpesvirus infection in abalone Haliotis laevigata and Haliotis rubra hybrids. Fish Shellfish Immunol. 2013, 34, 688–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruzzo, C.; Gallo, G.; Canesi, L. Persistence of vibrios in marine bivalves: The role of interactions with haemolymph components. Environ. Microbiol. 2005, 7, 761–772. [Google Scholar] [CrossRef]
- Segarra, A.; Faury, N.; Pépin, J.-F.; Renault, T. Transcriptomic study of 39 ostreid herpesvirus 1 genes during an experimental infection. J. Invertebr. Pathol. 2014, 119, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Green, T.J.; Vergnes, A.; Montagnani, C.; de Lorgeril, J. Distinct immune responses of juvenile and adult oysters (Crassostrea gigas) to viral and bacterial infections. Vet. Res. 2016, 47, 72. [Google Scholar] [CrossRef] [Green Version]
- Agriculture Department. Abalone Viral Ganglioneuritis (AVG); ABARES: Canberra, Australia, 2019.
- Garcia, C.; Thébault, A.; Dégremont, L.; Arzul, I.; Miossec, L.; Robert, M.; Chollet, B.; François, C.; Joly, J.-P.; Ferrand, S.; et al. Ostreid herpesvirus 1 detection and relationship with Crassostrea gigas spat mortality in France between 1998 and 2006. Vet. Res. 2011, 42, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divilov, K.; Schoolfield, B.; Morga, B.; Dégremont, L.; Burge, C.A.; Mancilla Cortez, D.; Friedman, C.S.; Fleener, G.B.; Dumbauld, B.R.; Langdon, C. First evaluation of resistance to both a California OsHV-1 variant and a French OsHV-1 microvariant in Pacific oysters. BMC Genet. 2019, 20, 96. [Google Scholar] [CrossRef] [Green Version]
- Dégremont, L.; Nourry, M.; Maurouard, E. Mass selection for survival and resistance to OsHV-1 infection in Crassostrea gigas spat in field conditions: Response to selection after four generations. Aquaculture 2015, 446, 111–121. [Google Scholar]
- Kube, P.; Dove, M.; Cunningham, M.; Kirkland, P.; Gu, X.; Hick, P.; O’Connor, W.; Elliot, N. Genetic Selection for Resistance to Pacific Oyster Mortality Syndrome. In CSIRO Marine and Atmospheric Research, FRDC and Seafood CRC Project 2012/760; CSIRO Agriculture and Food, NSW Department of Primary Industries, Australian Seafood Industries, Seafood CRC, and FRDC: Canberra, Australia, 2018; p. 55. [Google Scholar]
- Dégremont, L.; Morga, B.; Trancart, S.; Pépin, J.-F. Resistance to OsHV-1 Infection in Crassostrea gigas Larvae. Front. Mar. Sci. 2016, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Lafont, M.; Petton, B.; Vergnes, A.; Pauletto, M.; Segarra, A.; Gourbal, B.; Montagnani, C. Long-lasting antiviral innate immune priming in the Lophotrochozoan Pacific oyster, Crassostrea gigas. Sci. Rep. 2017, 7, 13143. [Google Scholar] [CrossRef]
- Milutinović, B.; Peuß, R.; Ferro, K.; Kurtz, J. Immune priming in arthropods: An update focusing on the red flour beetle. Zoology 2016, 119, 254–261. [Google Scholar]
- Criscitiello, M.F.; de Figueiredo, P. Fifty shades of immune defense. PLoS Pathog 2013, 9, e1003110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portela, J.; Duval, D.; Rognon, A.; Galinier, R.; Boissier, J.; Coustau, C.; Mitta, G.; Théron, A.; Gourbal, B. Evidence for Specific Genotype-Dependent Immune Priming in the Lophotrochozoan Biomphalaria glabrata Snail. J. Innate Immun. 2013, 5, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Dubief, B.; Nunes, F.L.D.; Basuyaux, O.; Paillard, C. Immune priming and portal of entry effectors improve response to vibrio infection in a resistant population of the European abalone. Fish Shellfish Immunol. 2017, 60, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Lafont, M.; Vergnes, A.; Vidal-Dupiol, J.; de Lorgeril, J.; Gueguen, Y.; Haffner, P.; Petton, B.; Chaparro, C.; Barrachina, C.; Destoumieux-Garzon, D.; et al. A Sustained Immune Response Supports Long-Term Antiviral Immune Priming in the Pacific Oyster, Crassostrea gigas. mBio 2020, 11, e02719–e02777. [Google Scholar] [CrossRef] [Green Version]
- Underwood, D.J.; Cowley, J.A.; Johnson, K.N. Antiviral immunity and protection in penaeid shrimp. Invertebr. Immun. 2013, 1, 2–14. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.N.; van Hulten, M.C.W.; Barnes, A.C. “Vaccination” of shrimp against viral pathogens: Phenomenology and underlying mechanisms. Vaccine 2008, 26, 4885–4892. [Google Scholar] [CrossRef]
- Green, T.J.; Montagnani, C.; Benkendorff, K.; Robinson, N.; Speck, P. Ontogeny and water temperature influences the antiviral response of the Pacific oyster, Crassostrea gigas. Fish Shellfish Immunol. 2014, 36, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Tetreau, G.; Dhinaut, J.; Gourbal, B.; Moret, Y. Trans-generational Immune Priming in Invertebrates: Current Knowledge and Future Prospects. Front. Immunol. 2019, 10, 1938. [Google Scholar] [CrossRef] [Green Version]
- Barbosa-Solomieu, V.; Dégremont, L.; Vázquez-Juárez, R.; Ascencio-Valle, F.; Boudry, P.; Renault, T. Ostreid Herpesvirus 1 (OsHV-1) detection among three successive generations of Pacific oysters (Crassostrea gigas). Virus Res. 2005, 107, 47–56. [Google Scholar] [CrossRef]
- Lafont, M.; Goncalves, P.; Guo, X.; Montagnani, C.; Raftos, D.; Green, T. Transgenerational plasticity and antiviral immunity in the Pacific oyster (Crassostrea gigas) against Ostreid herpesvirus 1 (OsHV-1). Dev. Comp. Immunol. 2019, 91, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, A.N.; Green, T.J. Fitness costs associated with maternal immune priming in the oyster. Fish Shellfish Immunol. 2020, 103, 32–36. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agius, J.R.; Corbeil, S.; Helbig, K.J. Immune Control of Herpesvirus Infection in Molluscs. Pathogens 2020, 9, 618. https://doi.org/10.3390/pathogens9080618
Agius JR, Corbeil S, Helbig KJ. Immune Control of Herpesvirus Infection in Molluscs. Pathogens. 2020; 9(8):618. https://doi.org/10.3390/pathogens9080618
Chicago/Turabian StyleAgius, Jacinta R, Serge Corbeil, and Karla J Helbig. 2020. "Immune Control of Herpesvirus Infection in Molluscs" Pathogens 9, no. 8: 618. https://doi.org/10.3390/pathogens9080618
APA StyleAgius, J. R., Corbeil, S., & Helbig, K. J. (2020). Immune Control of Herpesvirus Infection in Molluscs. Pathogens, 9(8), 618. https://doi.org/10.3390/pathogens9080618