
Antimicrobial Susceptibility of Persister Biofilm Cells of Bacillus cereus and Pseudomonas fluorescens





Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Biocides
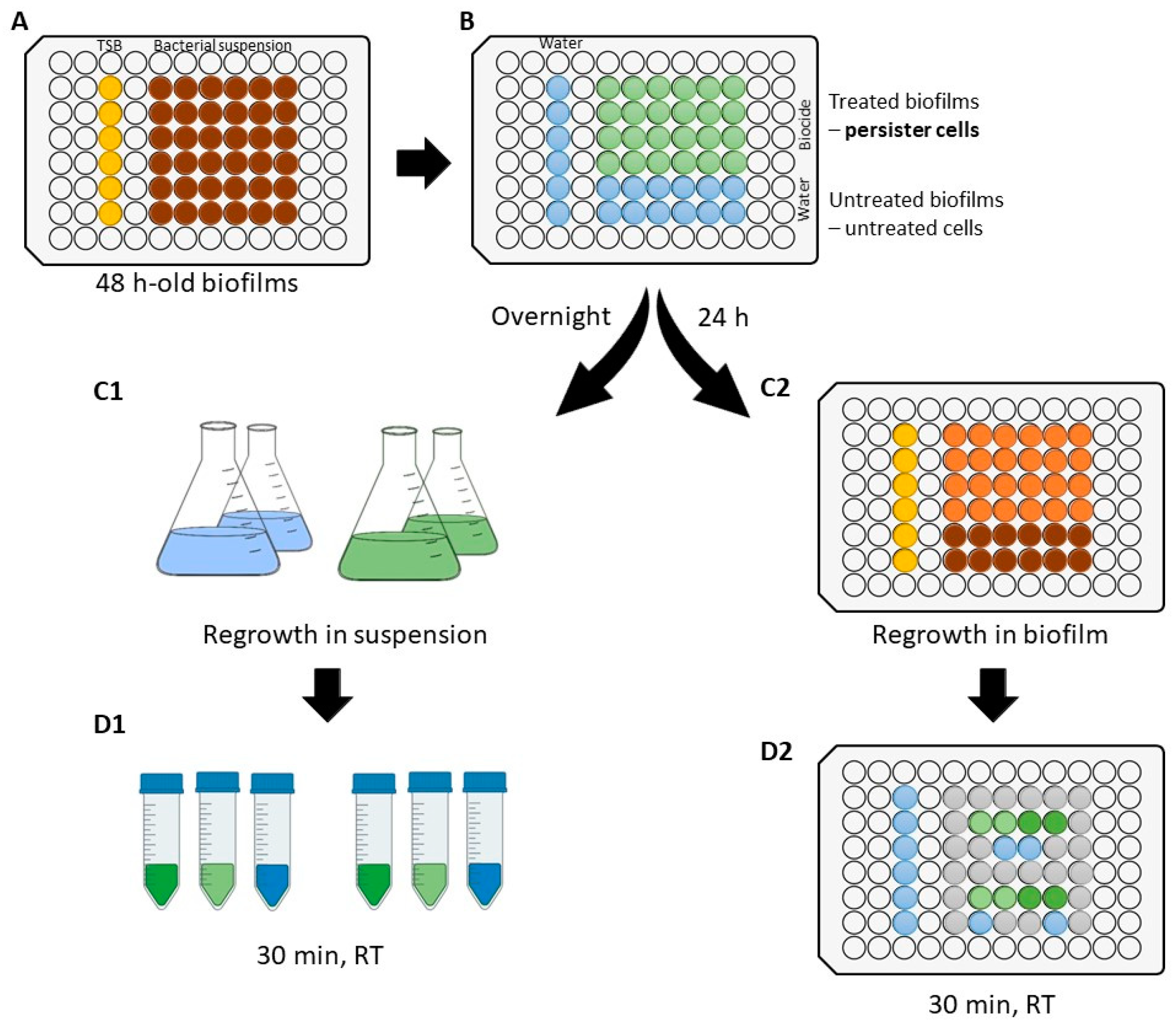
2.3. Experimental Setup
2.4. Development of Persister Cells after Biocide Treatment
2.5. Antimicrobial Activity against Regrown Cultures
2.5.1. Susceptibility of Regrown Cultures in Suspension
2.5.2. Susceptibility of Regrown Biofilms
2.6. Statistical Analysis
3. Results and Discussion
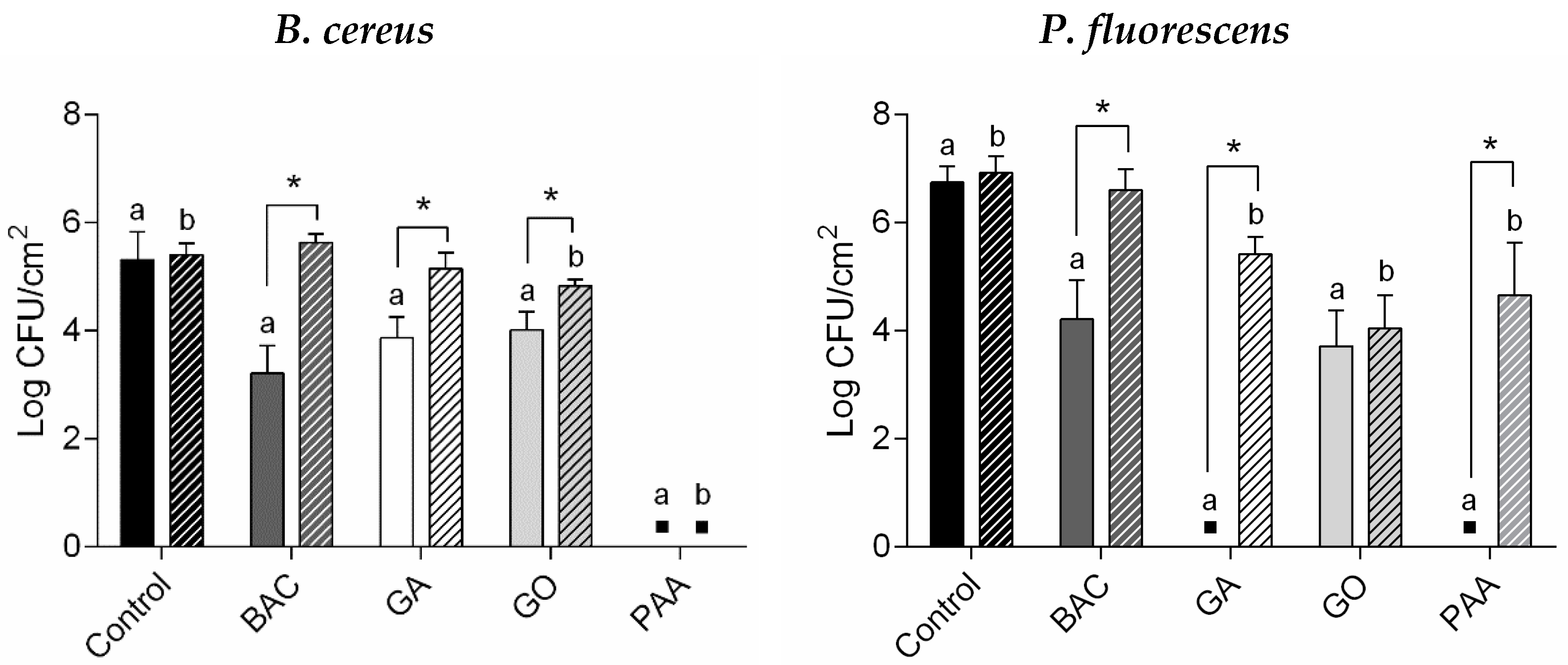
3.1. Persister Biofilm Cells after Biocide Treatment
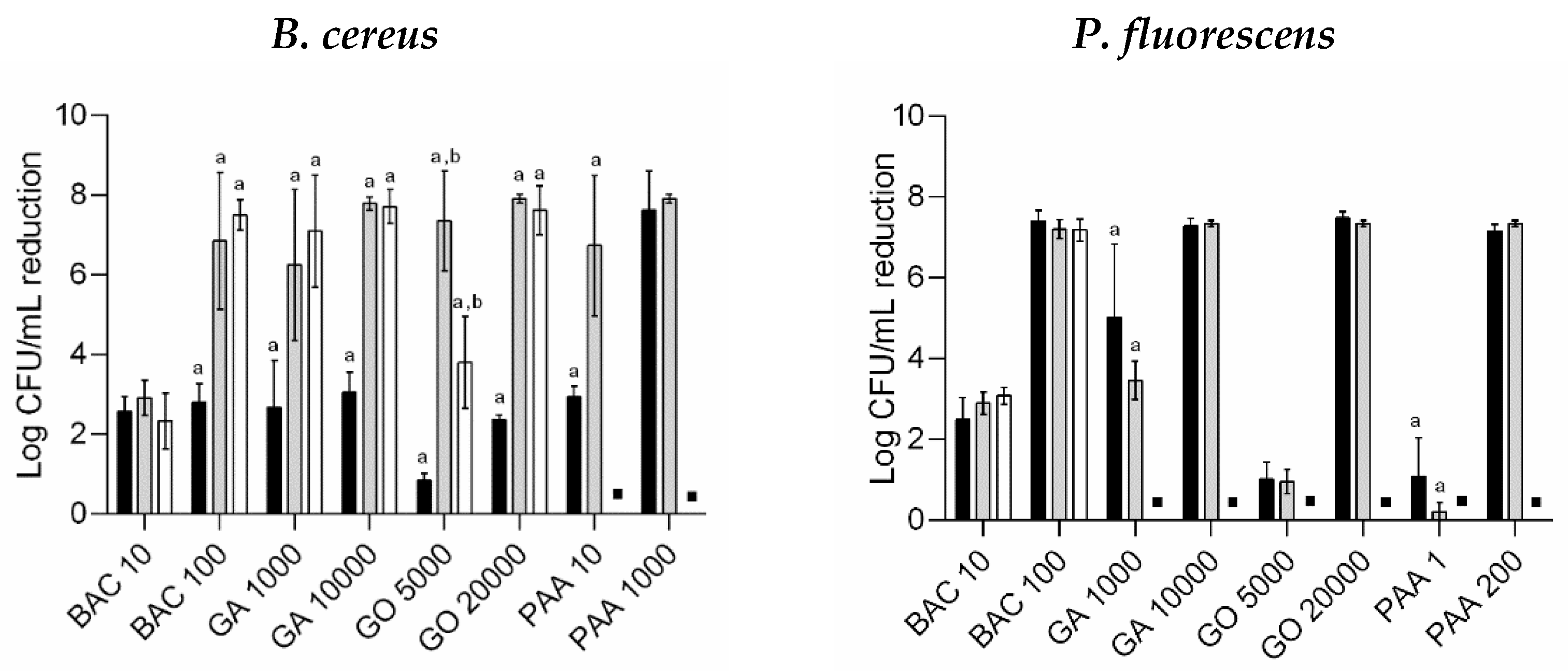
3.2. Antimicrobial Susceptibility of Regrown Cells in Suspension
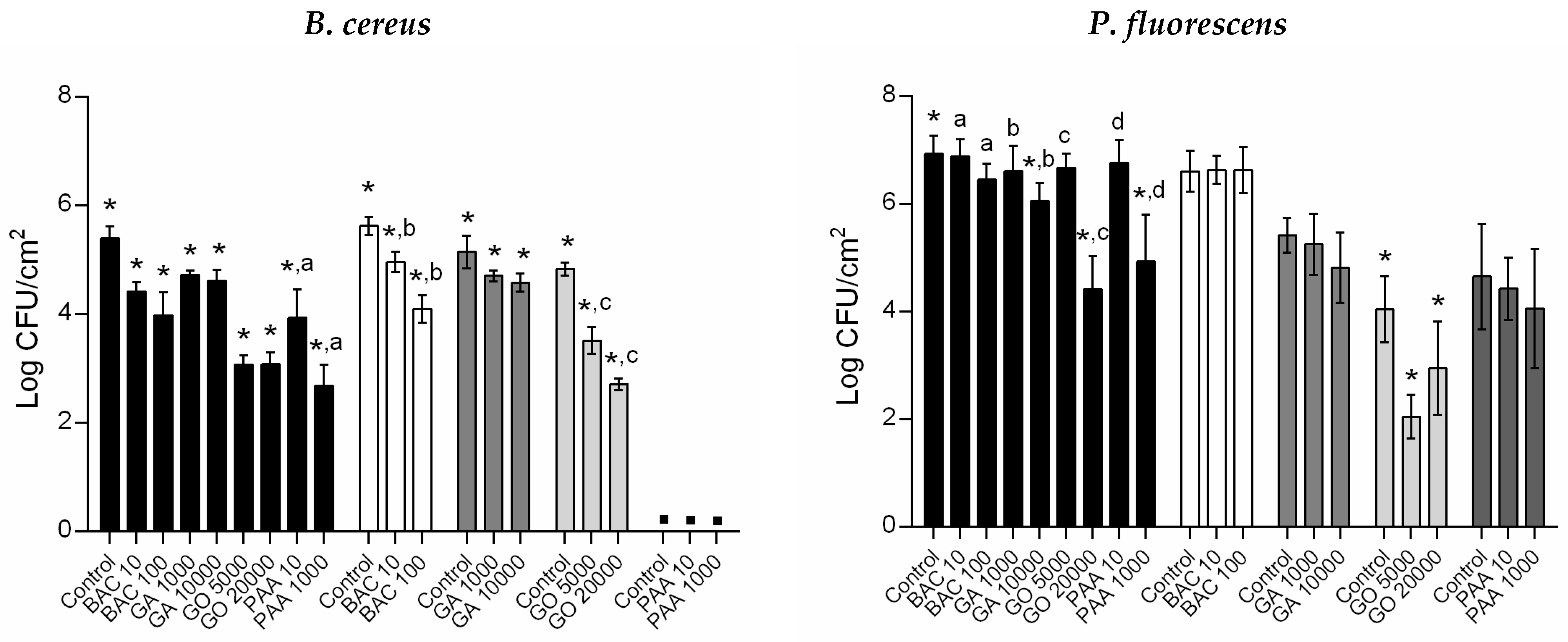
3.3. Antimicrobial Susceptibility of Regrown Biofilms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomes, I.B.; Simões, M.; Simões, L.C. An overview on the reactors to study drinking water biofilms. Water Res. 2014, 62, 63–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Tsai, W.; Chou, C. Inactivation and removal of Bacillus cereus by sanitizer and detergent. Int. J. Food. Microbiol. 2002, 77, 11–18. [Google Scholar] [CrossRef]
- Møretrø, T.; Langsrud, S.; Heir, E. Bacteria on meat abattoir process surfaces after sanitation: Characterisation of survival properties of Listeria monocytogenes and the commensal bacterial flora. Adv. Microbiol. 2013, 3, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Marouani-Gadri, N.; Augier, G.; Carpentier, B. Characterization of bacterial strains isolated from a beef-processing plant following cleaning and disinfection—Influence of isolated strains on biofilm formation by Sakai and EDL 933 E. coli O157:H7. Int. J. Food Microbiol. 2009, 133, 62–67. [Google Scholar] [CrossRef]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Davison, W.M.; Pitts, B.; Stewart, P.S. Spatial and temporal patterns of biocide action against Staphylococcus epidermidis biofilms. Antimicrob. Agents Chemother. 2010, 54, 2920–2927. [Google Scholar] [CrossRef] [Green Version]
- Podlesek, Z.; Zgur Bertok, D. The DNA Damage inducible SOS response is a key player in the generation of bacterial persister cells and population wide tolerance. Front. Microbiol. 2020, 11, 1785. [Google Scholar] [CrossRef]
- Stewart, P.S. Antimicrobial tolerance in biofilms. Microbiol. Spectrum. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simões, L.C.; Lemos, M.; Pereira, A.M.; Abreu, A.C.; Saavedra, M.J.; Simões, M. Persister cells in a biofilm treated with a biocide. Biofouling 2011, 27, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S.; White, B.; Boegli, L.; Hamerly, T.; Williamson, K.S.; Franklin, M.J.; Bothner, B.; James, G.A.; Fisher, S.; Vital-Lopez, F.G.; et al. Conceptual model of biofilm antibiotic tolerance that integrates phenomena of diffusion, metabolism, gene expression, and physiology. J. Bacteriol. 2019, 201, e00307-19. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef]
- Kaldalu, N.; Hauryliuk, V.; Turnbull, K.J.; La Mensa, A.; Putrinš, M.; Tenson, T. In vitro studies of persister cells. Microbiol. Mol. Biol. Rev. 2020, 84, e00070-20. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.; Galdino, A.C.M.; Mello, T.P.; Ramos, L.S.; Branquinha, M.H.; Bolognese, A.M.; Columbano Neto, J.; Roudbary, M. What are the advantages of living in a community? A microbial biofilm perspective! Mem. Inst. Oswaldo Cruz 2018, 113, e180212–e180217. [Google Scholar] [CrossRef] [Green Version]
- Dombach, J.L.; Quintana, J.L.J.; Detweiler, C.S. Staphylococcal bacterial persister cells, biofilms, and intracellular infection are disrupted by JD1, a membrane-damaging small molecule. mBio 2021, 12, e0180121. [Google Scholar] [CrossRef] [PubMed]
- Ortega Morente, E.; Fernandez-Fuentes, M.A.; Grande Burgos, M.J.; Abriouel, H.; Perez Pulido, R.; Galvez, A. Biocide tolerance in bacteria. Int. J. Food Microbiol. 2013, 162, 13–25. [Google Scholar] [CrossRef]
- Jiang, T.; Li, M. Quorum sensing inhibitors: A patent review (2014–2018). Expert. Opin. Ther. Pat. 2013, 23, 867–894. [Google Scholar] [CrossRef]
- Behnke, S.; Parker, A.E.; Woodall, D.; Camper, A.K. Comparing the chlorine disinfection of detached biofilm clusters with those of sessile biofilms and planktonic cells in single- and dual-species cultures. Appl. Environ. Microbiol. 2011, 77, 7176–7184. [Google Scholar] [CrossRef] [Green Version]
- Chebotar, I.V.; Emelyanova, M.A.; Bocharova, J.A.; Mayansky, N.A.; Kopantseva, E.E.; Mikhailovich, V.M. The classification of bacterial survival strategies in the presence of antimicrobials. Microb. Pathog. 2021, 155, 104901. [Google Scholar] [CrossRef]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef]
- Ng, W.J.; Tan, C.T.; Bae, S. Effects of monochloramine on culturability, viability and persistence of Pseudomonas putida and tap water mixed bacterial community. Appl. Microbiol. Biotechnol. 2021, 105, 3799–3810. [Google Scholar] [CrossRef] [PubMed]
- Cogan, N.G.; Szomolay, B.; Dindos, M. Effect of periodic disinfection on persisters in a one-dimensional biofilm model. Bull. Math. Biol. 2013, 75, 94–123. [Google Scholar] [CrossRef] [Green Version]
- Simões, M.; Cleto, S.; Pereira, M.O.; Vieira, M.J. Influence of biofilm composition on the resistance to detachment. Water Sci. Technol. 2007, 55, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Simões, L.C.; Pereira, M.O.; Vieira, M.J. Antagonism between Bacillus cereus and Pseudomonas fluorescens in planktonic systems and in biofilms. Biofouling 2008, 24, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Pan, J.; Bahar, A.A.; Syed, H.; Ren, S. Reverting antibiotic tolerance of Pseudomonas aeruginosa PAO1 persister cells by (Z)-4-bromo-5-(bromomethylene)-3-methylfuran-2(5H)-one. PLoS ONE 2012, 7, e45778. [Google Scholar] [CrossRef] [Green Version]
- European Standard—EN 1276; Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Bactericidal Activity of Chemical Disinfectants and Antiseptics Used in Food, Industrial, Domestic, and Institutional Areas—Test Method and Requirements (Phase 2, Step 1). iTeh, Inc.: Newark, DE, USA, 2009.
- Simões, M.; Simões, L.C.; Vieira, M.J. Species association increases biofilm resistance to chemical and mechanical treatments. Water Res. 2009, 43, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Simões, L.C.; Lemos, M.; Araújo, P.; Pereira, A.M.; Simões, M. The effects of glutaraldehyde on the control of single and dual biofilms of Bacillus cereus and Pseudomonas fluorescens. Biofouling 2011, 27, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Gomes, I.B.; Simões, L.C.; Simões, M. The effects of emerging environmental contaminants on Stenotrophomonas maltophilia isolated from drinking water in planktonic and sessile states. Sci. Total Environ. 2018, 643, 1348–1356. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, F.; Rezaee, M.A.; Ebrahimzadeh, S.; Yousefi, L.; Nouri, R.; Kafil, H.S.; Gholizadeh, P. Novel strategies to combat bacterial biofilms. Mol. Biotechnol. 2021, 63, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Fernandes, S.; Gomes, I.B.; Simões, M. Antibiofilm activity of glycolic acid and glyoxal and their diffusion-reaction interactions with biofilm components. Food Res. Int. 2022, 152, 110921. [Google Scholar] [CrossRef]
- Fernandes, S.; Gomes, I.B.; Simões, M. Antimicrobial activity of glycolic acid and glyoxal against Bacillus cereus and Pseudomonas fluorescens. Food Res. Int. 2020, 136, 109346. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Pan, H.; Yang, D.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. Induction, detection, formation, and resuscitation of viable but non-culturable state microorganisms. Compr. Rev. Food Sci. Food Saf. 2020, 19, 149–183. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhao, X. Effects of quorum sensing on the biofilm formation and viable but non-culturable state. Food Res. Int. 2020, 137, 109742. [Google Scholar] [CrossRef]
- Simões, M.; Simões, L.C.; Cleto, S.; Machado, I.; Pereira, M.O.; Vieira, M.J. Antimicrobial mechanisms of ortho-phthalaldehyde action. J. Basic Microbiol. 2007, 47, 230–242. [Google Scholar] [CrossRef] [Green Version]
- Tehri, N.; Kumar, N.; Raghu, H.V.; Vashishth, A. Biomarkers of bacterial spore germination. Ann. Microbiol. 2018, 68, 513–523. [Google Scholar] [CrossRef]
- Rollet, C.; Gal, L.; Guzzo, J. Biofilm-detached cells, a transition from a sessile to a planktonic phenotype: A comparative study of adhesion and physiological characteristics in Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2009, 290, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, F.; Wang, J.; Zhong, N. Biofilm formation and control strategies of foodborne pathogens: Food safety perspectives. RSC Adv. 2017, 7, 36670–36683. [Google Scholar] [CrossRef] [Green Version]
- Gomes, I.B.; Lemos, M.; Fernandes, S.; Borges, A.; Simões, L.C.; Simões, M. The effects of chemical and mechanical stresses on Bacillus cereus and Pseudomonas fluorescens single- and dual-species biofilm removal. Microorganisms 2021, 9, 1174. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G. Biocidal agents used for disinfection can enhance antibiotic resistance in Gram-negative species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordholt, N.; Kanaris, O.; Schmidt, S.B.I.; Schreiber, F. Persistence against benzalkonium chloride promotes rapid evolution of tolerance during periodic disinfection. Nat. Commun. 2021, 12, 6792. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.M.P.; Wang, X.; Tagkopoulos, I. Biocide-induced emergence of antibiotic resistance in Escherichia coli. Front. Microbiol. 2021, 12, 640923. [Google Scholar] [CrossRef] [PubMed]



 ), GO treatment (
), GO treatment (  ), and PAA treatment (
), and PAA treatment (  ). ▪—no detectable/significant regrown biofilms. *—Surviving cells from biocide exposure were significantly lower compared to unexposed regrown biofilms (control) for each population (Dunnett’s multiple comparisons test, p < 0.05); a, b, c, and d—Surviving cells were significantly different from distinct biocide concentration exposure (unpaired t-test with Welch’s correction, p < 0.05).
), GO treatment ( ), and PAA treatment ( ). ▪—no detectable/significant regrown biofilms. *—Surviving cells from biocide exposure were significantly lower compared to unexposed regrown biofilms (control) for each population (Dunnett’s multiple comparisons test, p < 0.05); a, b, c, and d—Surviving cells were significantly different from distinct biocide concentration exposure (unpaired t-test with Welch’s correction, p < 0.05).
). ▪—no detectable/significant regrown biofilms. *—Surviving cells from biocide exposure were significantly lower compared to unexposed regrown biofilms (control) for each population (Dunnett’s multiple comparisons test, p < 0.05); a, b, c, and d—Surviving cells were significantly different from distinct biocide concentration exposure (unpaired t-test with Welch’s correction, p < 0.05).
), GO treatment ( ), and PAA treatment ( ). ▪—no detectable/significant regrown biofilms. *—Surviving cells from biocide exposure were significantly lower compared to unexposed regrown biofilms (control) for each population (Dunnett’s multiple comparisons test, p < 0.05); a, b, c, and d—Surviving cells were significantly different from distinct biocide concentration exposure (unpaired t-test with Welch’s correction, p < 0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. cereus | P. fluorescens | |||
|---|---|---|---|---|
| Concentration (µg/mL) | Exposure Time (h) | Concentration (µg/mL) | Exposure Time (h) | |
| BAC | 500 | 0.5 | 1000 | 4 |
| GA | 1000 | 0.5 | 40,000 | 0.5 |
| GO | 20,000 | 0.5 | 20,000 | 0.5 |
| PAA | 10,000 | 0.5 | 20,000 | 0.5 |
| B. cereus | P. fluorescens | |||
|---|---|---|---|---|
| Suspension | Biofilm | Suspension | Biofilm | |
| BAC | 10, 100 | 10, 100 | ||
| GA | 1000, 10,000 | 1000, 10,000 | ||
| GO | 5000, 20,000 | 5000, 20,000 | ||
| PAA | 10, 1000 | 1, 200 | 10, 1000 | |
| B. cereus | P. fluorescens | ||||
|---|---|---|---|---|---|
| Untreated | Persister | Untreated | Persister | ||
| BAC | 10 | 12 ± 7 | 28 ± 10 | 72 ± 23 | 78 ± 25 |
| 100 | 4 ± 3 | 4 ± 3 | 28 ± 10 * | 77 ± 22 * | |
| GA | 1000 | 20 ± 5 | 37 ± 23 | 52 ± 33 | 64 ± 29 |
| 10,000 | 16 ± 4 | 34 ± 26 | 25 ± 21 | 40 ± 27 | |
| GO | 5000 | 0.5 ± 0.3 * | 6 ± 3 * | 44 ± 24 * | 2 ± 3 * |
| 20,000 | 0.6 ± 0.3 | 0.8 ± 0.3 | 0.5 ± 0.7 | 9 ± 9 | |
| PAA | 10 | 5 ± 4 | -- | 58 ± 30 | 35 ± 21 |
| 1000 | 0.2 ± 0.1 | -- | 2 ± 4 | 23 ± 24 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, S.; Gomes, I.B.; Sousa, S.F.; Simões, M. Antimicrobial Susceptibility of Persister Biofilm Cells of Bacillus cereus and Pseudomonas fluorescens. Microorganisms 2022, 10, 160. https://doi.org/10.3390/microorganisms10010160
Fernandes S, Gomes IB, Sousa SF, Simões M. Antimicrobial Susceptibility of Persister Biofilm Cells of Bacillus cereus and Pseudomonas fluorescens. Microorganisms. 2022; 10(1):160. https://doi.org/10.3390/microorganisms10010160
Chicago/Turabian StyleFernandes, Susana, Inês B. Gomes, Sérgio F. Sousa, and Manuel Simões. 2022. "Antimicrobial Susceptibility of Persister Biofilm Cells of Bacillus cereus and Pseudomonas fluorescens" Microorganisms 10, no. 1: 160. https://doi.org/10.3390/microorganisms10010160
APA StyleFernandes, S., Gomes, I. B., Sousa, S. F., & Simões, M. (2022). Antimicrobial Susceptibility of Persister Biofilm Cells of Bacillus cereus and Pseudomonas fluorescens. Microorganisms, 10(1), 160. https://doi.org/10.3390/microorganisms10010160