
Commensal and Opportunistic Bacteria Present in the Microbiota in Atlantic Cod (Gadus morhua) Larvae Differentially Alter the Hosts’ Innate Immune Responses

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cod Larval Rearing
2.2. Sampling
2.3. Characterisation of Microbial Communities
2.4. Microarray Design, Hybridisation and Annotation
2.5. Electron Microscopy Procedures
2.6. Intestinal Morphometry and Statistical Methods
2.7. Statistical Analyses
3. Results
3.1. Larval Survival and Growth
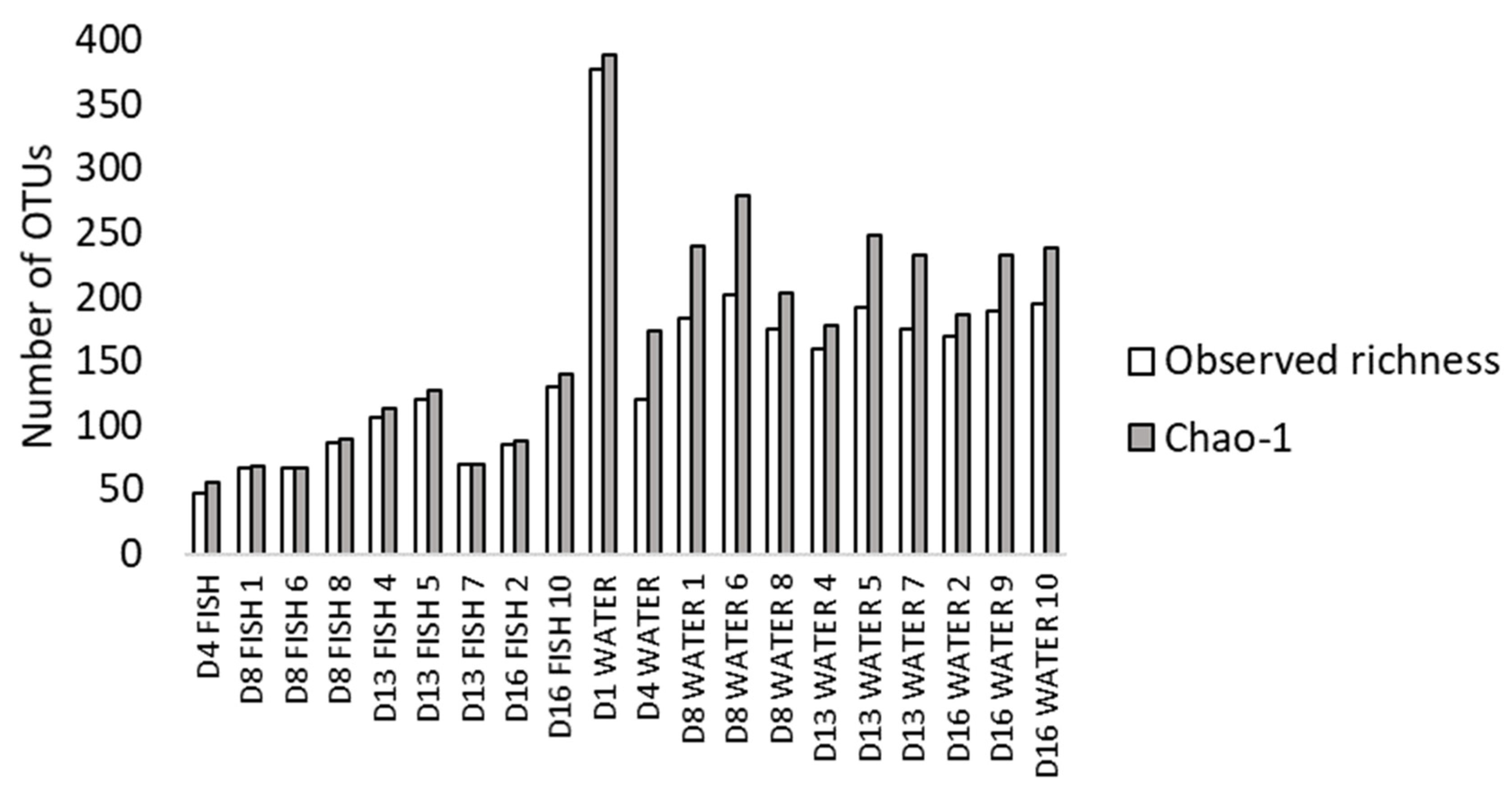
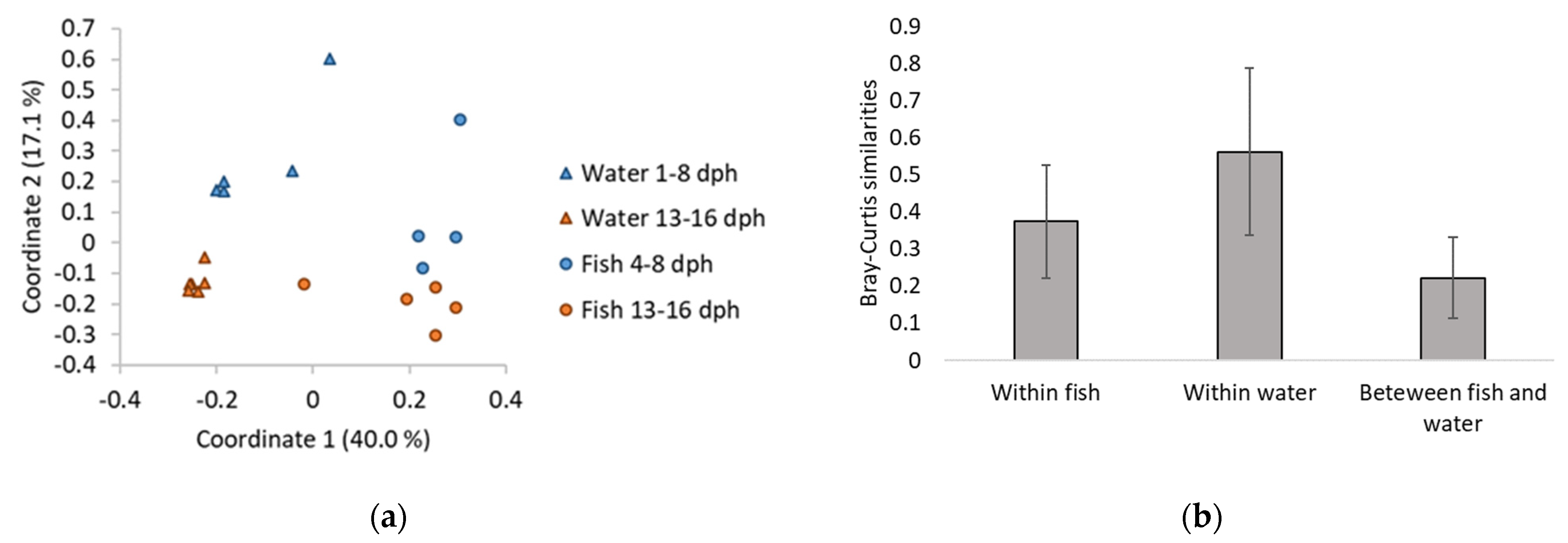
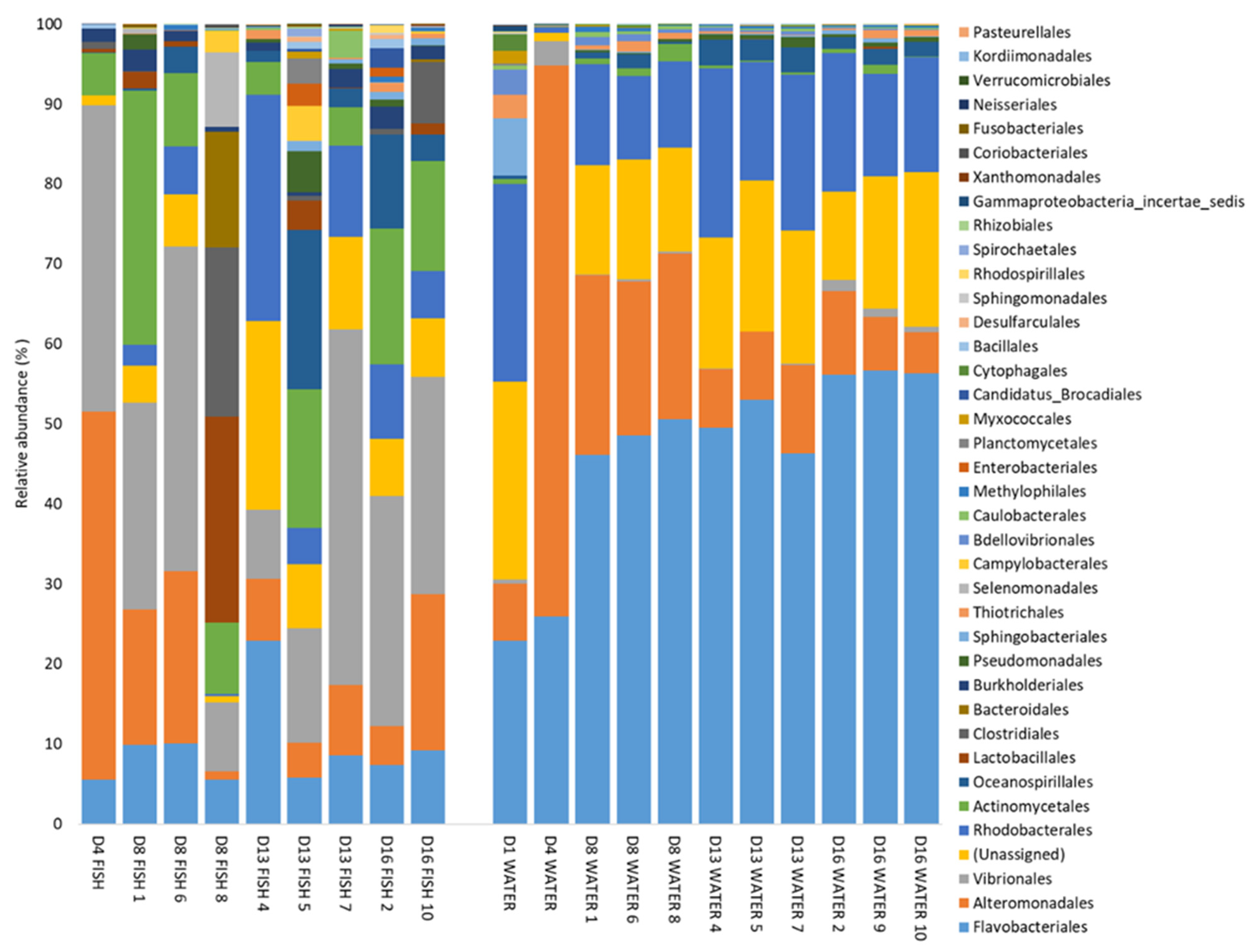
3.2. Composition of Fish and Water Microbiota
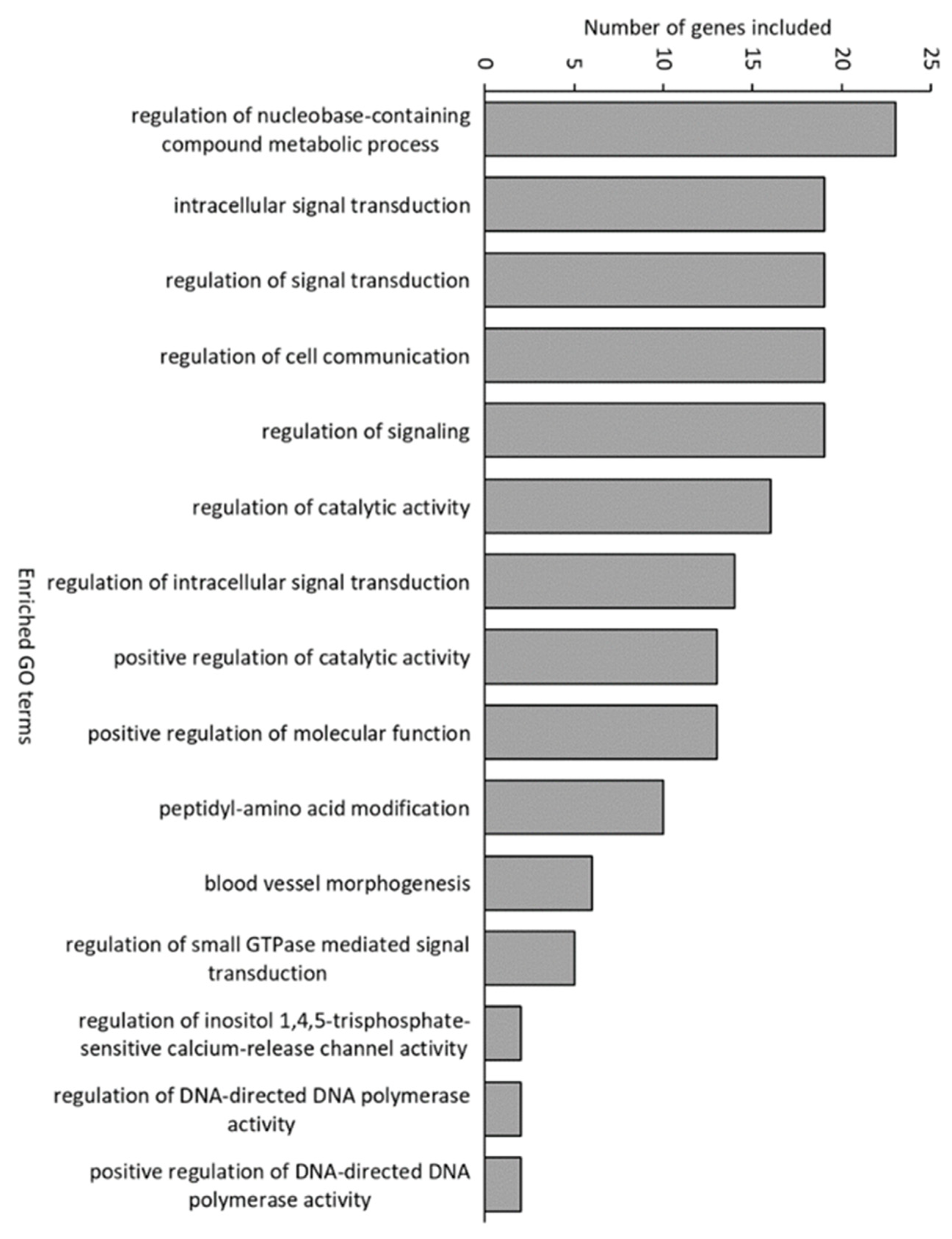
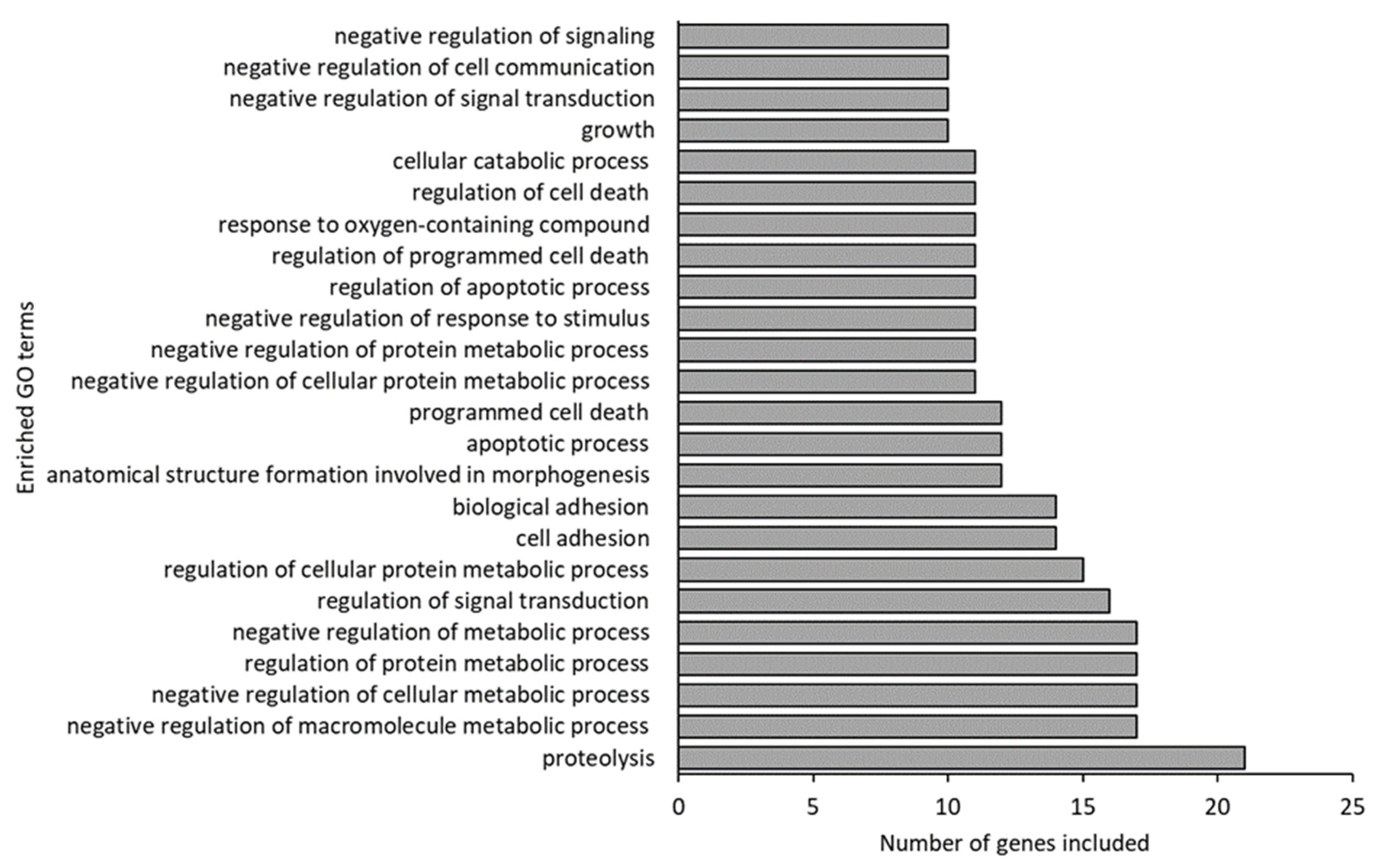
3.3. Gene Expression in Atlantic Cod Larvae
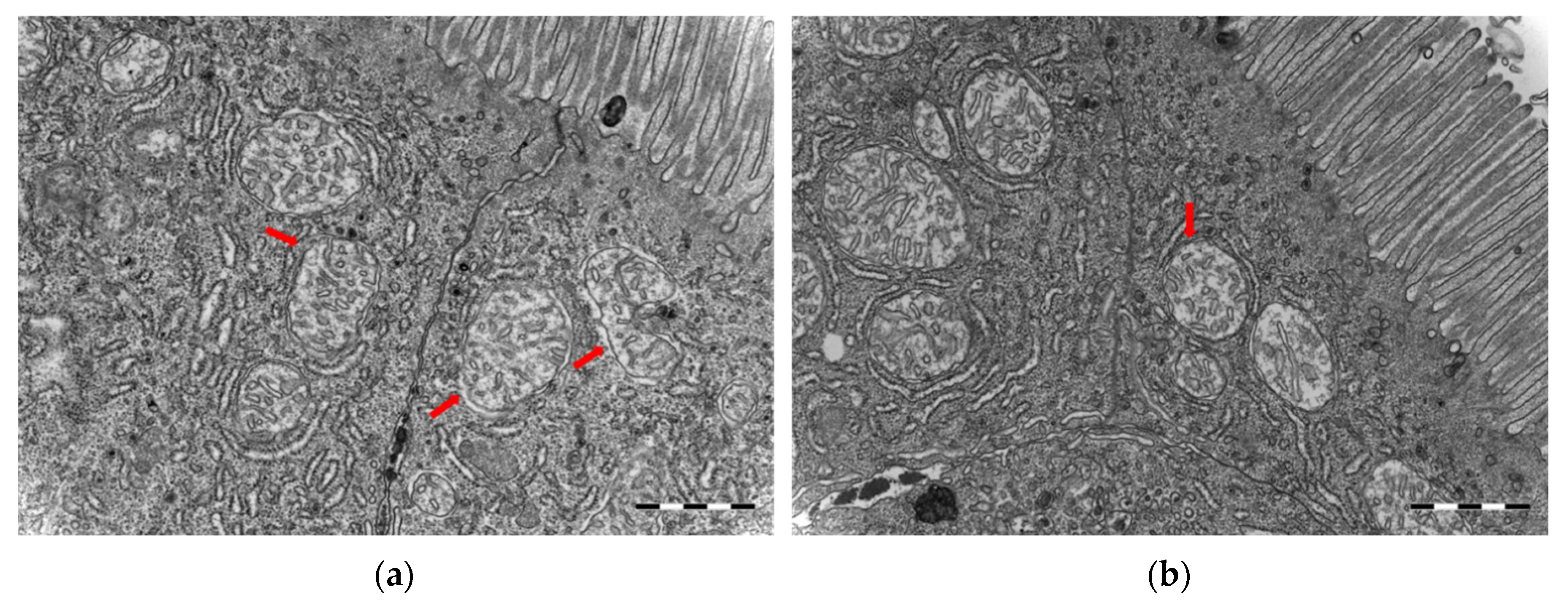
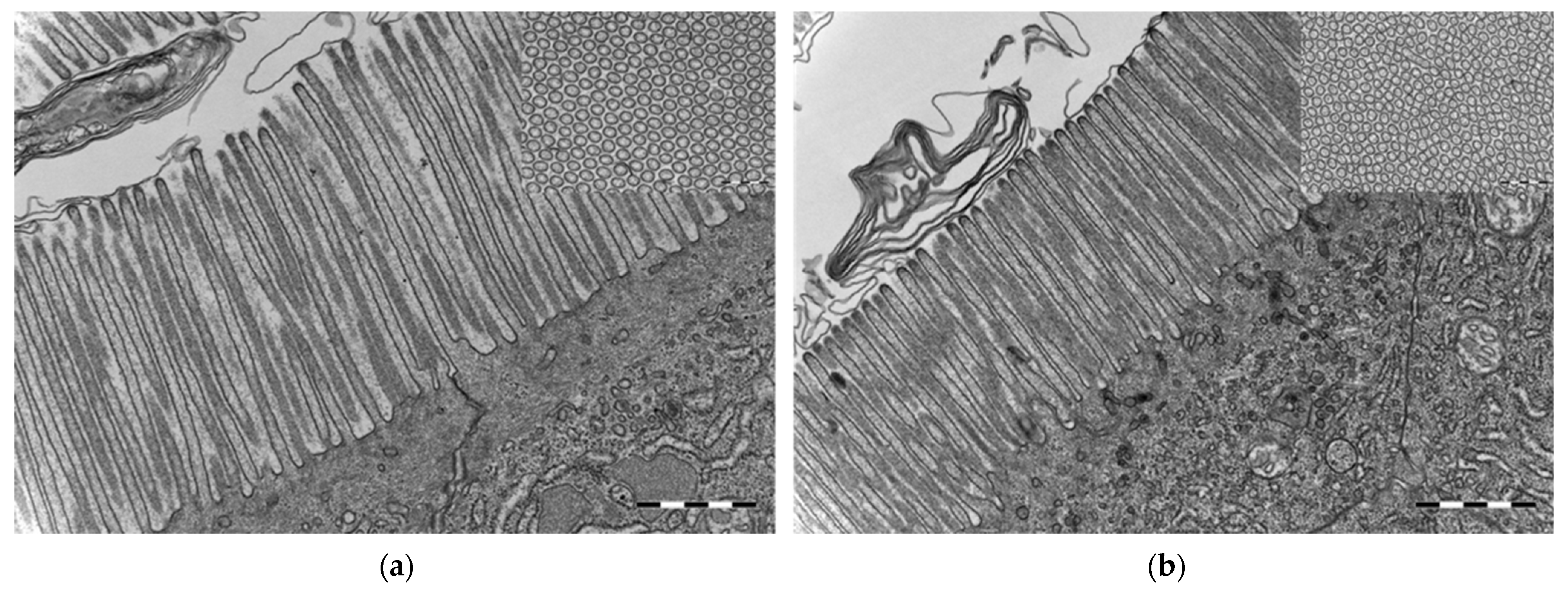
3.4. Ultrastructure and Morphometric Analysis of the Intestinal Tissue
4. Discussion
4.1. Microbial Environments
4.2. Presence of Bacteria Downregulates Host Responses Related to Nutrient Utilisation and Innate Immune Responses
4.3. Presence of Bateria Induces Host Responses Related to Inflammatory Responses and Signalling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hiippala, K.; Jouhten, H.; Ronkainen, A.; Hartikainen, A.; Kainulainen, V.; Jalanka, J.; Satokari, R. The potential of gut commensals in reinforcing intestinal barrier function and alleviating inflammation. Nutrients 2018, 10, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazado, C.C.; Caipang, C.M.A. Probiotics–pathogen interactions elicit differential regulation of cutaneous immune responses in epidermal cells of Atlantic cod Gadus morhua. Fish Shellfish Immunol. 2014, 36, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Lazado, C.C.; Caipang, C.M.A.; Brinchmann, M.F.; Kiron, V. In vitro adherence of two candidate probiotics from Atlantic cod and their interference with the adhesion of two pathogenic bacteria. Vet. Microbiol. 2011, 148, 252–259. [Google Scholar] [CrossRef]
- Kanther, M.; Rawls, J.F. Host–microbe interactions in the developing zebrafish. Curr. Opin. Immunol. 2010, 22, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Naito, T.; Mulet, C.; De Castro, C.; Molinaro, A.; Saffarian, A.; Nigro, G.; Bérard, M.; Clerc, M.; Pedersen, A.B.; Sansonetti, P.J.; et al. Lipopolysaccharide from crypt-specific core microbiota modulates the colonic epithelial proliferation-to-differentiation balance. mBio 2017, 8, e01680-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semova, I.; Carten, J.D.; Stombaugh, J.; Mackey, L.C.; Knight, R.; Farber, S.A.; Rawls, J.F. Microbiota Regulate Intestinal Absorption and Metabolism of Fatty Acids in the Zebrafish. Cell Host Microbe 2012, 12, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Birkbeck, T. Intestinal microflora of fish larvae and fry. Aquac. Res. 1999, 30, 73–93. [Google Scholar] [CrossRef]
- Vadstein, O.; Bergh, Ø.; Gatesoupe, F.-J.; Galindo-Villegas, J.; Mulero, V.; Picchietti, S.; Scapigliati, G.; Makridis, P.; Olsen, Y.; Dierckens, K.; et al. Microbiology and immunology of fish larvae. Rev. Aquac. 2013, 5, S1–S25. [Google Scholar] [CrossRef] [Green Version]
- Olafsen, J.A. Interactions between fish larvae and bacteria in marine aquaculture. Aquaculture 2001, 200, 223–247. [Google Scholar] [CrossRef]
- Vadstein, O.; Mo, T.A.; Bergh, Ø. Microbial Interactions, Prophylaxis and Diseases. In Culture of Cold-Water Marine Fish; Moksness, E., Kjørsvik, E., Olsen, Y., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2003; pp. 28–72. [Google Scholar] [CrossRef]
- Toranzo, A.E.; Magariños, B.; Romalde, J.L. A review of the main bacterial fish diseases in mariculture systems. Aquaculture 2005, 246, 37–61. [Google Scholar] [CrossRef]
- Forberg, T.; Milligan-Myhre, K. Chapter 6—Gnotobiotic Fish as Models to Study Host–Microbe Interactions. In Gnotobiotics; Schoeb, T.R., Eaton, K.A., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 369–383. [Google Scholar] [CrossRef]
- Smith, K.; McCoy, K.D.; Macpherson, A.J. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin. Immunol. 2007, 19, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Pham, L.N.; Kanther, M.; Semova, I.; Rawls, J.F. Methods for generating and colonizing gnotobiotic zebrafish. Nat. Protoc. 2008, 3, 1862. [Google Scholar] [CrossRef] [PubMed]
- Rawls, J.F.; Samuel, B.S.; Gordon, J.I. Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. USA 2004, 101, 4596–4601. [Google Scholar] [CrossRef] [Green Version]
- Uribe, A.; Alam, M.; Midtvedt, T.; Smedfors, B.; Theodorsson, E. Endogenous Prostaglandins and Microflora Modulate DNA Synthesis and Neuroendocrine Peptides in the Rat Gastrointestinal Tract. Scand. J. Gastroenterol. 1997, 32, 691–699. [Google Scholar] [CrossRef]
- Forberg, T.; Arukwe, A.; Vadstein, O. A protocol and cultivation system for gnotobiotic Atlantic cod larvae (Gadus morhua L.) as a tool to study host microbe interactions. Aquaculture 2011, 315, 222–227. [Google Scholar] [CrossRef]
- Forberg, T.; Vestrum, R.I.; Arukwe, A.; Vadstein, O. Bacterial composition and activity determines host gene-expression responses in gnotobiotic Atlantic cod (Gadus morhua) larvae. Vet. Microbiol. 2012, 157, 420–427. [Google Scholar] [CrossRef]
- Forberg, T.; Arukwe, A.; Vadstein, O. Two strategies to unravel gene expression responses of host–microbe interactions in cod (Gadus morhua) larvae. Aquac. Res. 2011, 42, 664–676. [Google Scholar] [CrossRef]
- Star, B.; Nederbragt, A.J.; Jentoft, S.; Grimholt, U.; Malmstrom, M.; Gregers, T.F.; Rounge, T.B.; Paulsen, J.; Solbakken, M.H.; Sharma, A.; et al. The genome sequence of Atlantic cod reveals a unique immune system. Nature 2011, 477, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Solbakken, M.H.; Jentoft, S.; Reitan, T.; Mikkelsen, H.; Gregers, T.F.; Bakke, O.; Jakobsen, K.S.; Seppola, M. Disentangling the immune response and host-pathogen interactions in Francisella noatunensis infected Atlantic cod. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 30, 333–346. [Google Scholar] [CrossRef]
- Tørresen, O.K.; Rise, M.L.; Jin, X.; Star, B.; MacKenzie, S.; Jakobsen, K.S.; Nederbragt, A.J.; Jentoft, S. An improved version of the Atlantic cod genome and advancements in functional genomics: Implications for the future of cod farming. In Genomics in Aquaculture; Academic Press: Cambridge, MA, USA, 2016; pp. 45–72. [Google Scholar] [CrossRef]
- Star, B.; Jentoft, S. Why does the immune system of Atlantic cod lack MHC II? BioEssays 2012, 34, 648–651. [Google Scholar] [CrossRef] [Green Version]
- Fjellheim, A.J.; Klinkenberg, G.; Skjermo, J.; Aasen, I.M.; Vadstein, O. Selection of candidate probionts by two different screening strategies from Atlantic cod (Gadus morhua L.) larvae. Vet. Microbiol. 2010, 144, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Bakke, I.; De Schryver, P.; Boon, N.; Vadstein, O. PCR-based community structure studies of Bacteria associated with eukaryotic organisms: A simple PCR strategy to avoid co-amplification of eukaryotic DNA. J. Microbiol. Methods 2011, 84, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Bakke, I.; Skjermo, J.; Vo, T.A.; Vadstein, O. Live feed is not a major determinant of the microbiota associated with cod larvae (Gadus morhua). Environ. Microbiol. Rep. 2013, 5, 537–548. [Google Scholar] [CrossRef]
- Vestrum, R.I.; Attramadal, K.J.K.; Vadstein, O.; Gundersen, M.S.; Bakke, I. Bacterial community assembly in Atlantic cod larvae (Gadus morhua): Contributions of ecological processes and metacommunity structure. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Mollerup, S.; Friis-Nielsen, J.; Vinner, L.; Hansen, T.A.; Richter, S.R.; Fridholm, H.; Herrera, J.A.R.; Lund, O.; Brunak, S.; Izarzugaza, J.M.G.; et al. Propionibacterium acnes: Disease-Causing Agent or Common Contaminant? Detection in Diverse Patient Samples by Next-Generation Sequencing. J. Clin. Microbiol. 2016, 54, 980. [Google Scholar] [CrossRef] [Green Version]
- Kleppe, L.; Edvardsen, R.B.; Furmanek, T.; Taranger, G.L.; Wargelius, A. Global transcriptome analysis identifies regulated transcripts and pathways activated during oogenesis and early embryogenesis in atlantic cod. Mol. Reprod. Dev. 2014, 81, 619–635. [Google Scholar] [CrossRef] [Green Version]
- Vestrum, R.I.; Attramadal, K.J.K.; Winge, P.; Li, K.; Olsen, Y.; Bones, A.M.; Vadstein, O.; Bakke, I. Rearing water treatment induces microbial selection influencing the microbiota and pathogen associated transcripts of cod (Gadus morhua) larvae. Front. Microbiol. 2018, 9, 851. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2008, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Zheng, X.; Yang, J.; Imamichi, T.; Stephens, R.; Lempicki, R.A. Extracting biological meaning from large gene lists with DAVID. Curr. Protoc. Bioinform. 2009, 27, 13.11.1–13.11.13. [Google Scholar] [CrossRef] [PubMed]
- Galloway, T.F.; Kjørsvik, E.; Kryvi, H. Effect of temperature on viability and axial muscle development in embryos and yolk sac larvae of the Northeast Arctic cod (Gadus morhua). Mar. Biol. 1998, 132, 559–567. [Google Scholar] [CrossRef]
- Brown, A.L., Jr. Microvilli of the human jejunal epithelial cell. J. Cell Biol. 1962, 12, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [Green Version]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and Processes of Microbial Community Assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342. [Google Scholar] [CrossRef] [Green Version]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Heilig, H.; Benvenuti, G.; Verreth, J.; Verdegem, M. The impact of rearing environment on the development of gut microbiota in tilapia larvae. Sci. Rep. 2015, 5, 18206. [Google Scholar] [CrossRef] [Green Version]
- Califano, G.; Castanho, S.; Soares, F.; Ribeiro, L.; Cox, C.J.; Mata, L.; Costa, R. Molecular Taxonomic Profiling of Bacterial Communities in a Gilthead Seabream (Sparus aurata) Hatchery. Front. Microbiol. 2017, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Giatsis, C.; Sipkema, D.; Smidt, H.; Verreth, J.; Verdegem, M. The colonization dynamics of the gut microbiota in tilapia larvae. PLoS ONE 2014, 9, e103641. [Google Scholar] [CrossRef] [Green Version]
- Vadstein, O.; Attramadal, K.J.K.; Bakke, I.; Olsen, Y. K-Selection as Microbial Community Management Strategy: A Method for Improved Viability of Larvae in Aquaculture. Front. Microbiol. 2018, 9, 2730. [Google Scholar] [CrossRef] [Green Version]
- Ina-Salwany, M.Y.; Al-saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Sonnenburg, J.L. Starving our Microbial Self: The Deleterious Consequences of a Diet Deficient in Microbiota-Accessible Carbohydrates. Cell Metab. 2014, 20, 779–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazado, C.C.; Caipang, C.M.A.; Kiron, V. Enzymes from the gut bacteria of Atlantic cod, Gadus morhua and their influence on intestinal enzyme activity. Aquac. Nutr. 2012, 18, 423–431. [Google Scholar] [CrossRef]
- Kjørsvik, E.; Pittman, K.; Pavlov, D. From fertilisation to the end of metamorphosis—Functional development. In Culture of Cold-Water Marine Fish; Moksness, E., Kjørsvik, E., Olsen, Y., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2004. [Google Scholar] [CrossRef]
- Kjørsvik, E.; van der Meeren, T.; Kryvi, H.; Arnfinnson, J.; Kvenseth, P.G. Early development of the digestive tract of cod larvae, Gadus morhua L., during start-feeding and starvation. J. Fish Biol. 1991, 38, 1–15. [Google Scholar] [CrossRef]
- Reddy, B.S.; Pleasants, J.R.; Wostmann, B.S. Pancreatic Enzymes in Germfree and Conventional Rats Fed Chemically Defined, Water-soluble Diet Free from Natural Substrates. J. Nutr. 1969, 97, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, C.L.; Dong, J.; Galipeau, H.J.; Jury, J.; McCarville, J.; Huang, X.; Wang, X.Y.; Naidoo, A.; Anbazhagan, A.N.; Libertucci, J.; et al. Commensal microbiota induces colonic barrier structure and functions that contribute to homeostasis. Sci. Rep. 2018, 8, 14184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meslin, J.C.; Sacquet, E.; Delpal, S. Effects of microflora on the dimensions of enterocyte microvilli in the rat. Reprod. Nutr. Développement 1984, 24, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Willing, B.P.; Van Kessel, A.G. Enterocyte proliferation and apoptosis in the caudal small intestine is influenced by the composition of colonizing commensal bacteria in the neonatal gnotobiotic pig. J. Anim. Sci. 2007, 85, 3256–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hailey, D.W.; Rambold, A.S.; Satpute-Krishnan, P.; Mitra, K.; Sougrat, R.; Kim, P.K.; Lippincott-Schwartz, J. Mitochondria Supply Membranes for Autophagosome Biogenesis during Starvation. Cell 2010, 141, 656–667. [Google Scholar] [CrossRef] [Green Version]
- Kohchi, C.; Inagawa, H.; Nishizawa, T.; Soma, G.-I. ROS and innate immunity. Anticancer Res. 2009, 29, 817–821. [Google Scholar] [PubMed]
- Hall, C.J.; Boyle, R.H.; Astin, J.W.; Flores, M.V.; Oehlers, S.H.; Sanderson, L.E.; Ellett, F.; Lieschke, G.J.; Crosier, K.E.; Crosier, P.S. Immunoresponsive Gene 1 Augments Bactericidal Activity of Macrophage-Lineage Cells by Regulating β-Oxidation-Dependent Mitochondrial ROS Production. Cell Metab. 2013, 18, 265–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, S.; Qi, J.; Echtenkamp, S.F.; Chatterjee, R.; Wang, M.; Boons, G.J.; Dziarski, R.; Gupta, D. Zebrafish peptidoglycan recognition proteins are bactericidal amidases essential for defense against bacterial infections. Immunity 2007, 27, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Royet, J.; Gupta, D.; Dziarski, R. Peptidoglycan recognition proteins: Modulators of the microbiome and inflammation. Nat. Rev. Immunol. 2011, 11, 837–851. [Google Scholar] [CrossRef] [PubMed]
- Thongda, W.; Li, C.; Luo, Y.; Beck, B.H.; Peatman, E. l-Rhamnose-binding lectins (RBLs) in channel catfish, Ictalurus punctatus: Characterization and expression profiling in mucosal tissues. Dev. Comp. Immunol. 2014, 44, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Vasta, G.R.; Nita-Lazar, M.; Giomarelli, B.; Ahmed, H.; Du, S.; Cammarata, M.; Parrinello, N.; Bianchet, M.A.; Amzel, L.M. Structural and functional diversity of the lectin repertoire in teleost fish: Relevance to innate and adaptive immunity. Dev. Comp. Immunol. 2011, 35, 1388–1399. [Google Scholar] [CrossRef]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal Alkaline Phosphatase Detoxifies Lipopolysaccharide and Prevents Inflammation in Zebrafish in Response to the Gut Microbiota. Cell Host Microbe 2007, 2, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Milligan-Myhre, K.; Small, C.M.; Mittge, E.K.; Agarwal, M.; Currey, M.; Cresko, W.A.; Guillemin, K. Innate immune responses to gut microbiota differ between oceanic and freshwater threespine stickleback populations. Dis. Models Mech. 2016, 9, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Solbakken, M.H.; Tørresen, O.K.; Nederbragt, A.J.; Seppola, M.; Gregers, T.F.; Jakobsen, K.S.; Jentoft, S. Evolutionary redesign of the Atlantic cod (Gadus morhua L.) Toll-like receptor repertoire by gene losses and expansions. Sci. Rep. 2016, 6, 25211. [Google Scholar] [CrossRef]
- Seppola, M.; Mikkelsen, H.; Johansen, A.; Steiro, K.; Myrnes, B.; Nilsen, I.W. Ultrapure LPS induces inflammatory and antibacterial responses attenuated in vitro by exogenous sera in Atlantic cod and Atlantic salmon. Fish Shellfish Immunol. 2015, 44, 66–78. [Google Scholar] [CrossRef]
- Bakkemo, K.R.; Mikkelsen, H.; Bordevik, M.; Torgersen, J.; Winther-Larsen, H.C.; Vanberg, C.; Olsen, R.; Johansen, L.H.; Seppola, M. Intracellular localisation and innate immune responses following Francisella noatunensis infection of Atlantic cod (Gadus morhua) macrophages. Fish Shellfish Immunol. 2011, 31, 993–1004. [Google Scholar] [CrossRef]
- Holen, E.; Lie, K.K.; Araujo, P.; Olsvik, P.A. Pathogen recognition and mechanisms in Atlantic cod (Gadus morhua) head kidney cells: Bacteria (LPS) and virus (poly I:C) signals through different pathways and affect distinct genes. Fish Shellfish Immunol. 2012, 33, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Brugman, S.; Witte, M.; Scholman, R.C.; Klein, M.R.; Boes, M.; Nieuwenhuis, E.E.S. T Lymphocyte–Dependent and–Independent Regulation of Cxcl8 Expression in Zebrafish Intestines. J. Immunol. 2014, 192, 484–491. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Cong, B.; Yu, S.; Liu, Y.; Man, X.; Wang, L.; Zhu, Q. Effect of a LECT2 on the immune response of peritoneal lecukocytes against Vibrio anguillarum in roughskin sculpin. Fish Shellfish Immunol. 2018, 74, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Oehlers, S.H.B.; Flores, M.V.; Hall, C.J.; O’Toole, R.; Swift, S.; Crosier, K.E.; Crosier, P.S. Expression of zebrafish cxcl8 (interleukin-8) and its receptors during development and in response to immune stimulation. Dev. Comp. Immunol. 2010, 34, 352–359. [Google Scholar] [CrossRef]
- Van Der Aa, L.M.; Chadzinska, M.; Tijhaar, E.; Boudinot, P.; Verburg-van Kemenade, B.M.L. CXCL8 chemokines in teleost fish: Two lineages with distinct expression profiles during early phases of inflammation. PLoS ONE 2010, 5, e12384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.; Chen, S.; Cao, Z.; Lin, Y.; Mo, D.; Zhang, H.; Gu, J.; Dong, M.; Liu, Z.; Xu, A. Acute phase response in zebrafish upon Aeromonas salmonicida and Staphylococcus aureus infection: Striking similarities and obvious differences with mammals. Mol. Immunol. 2007, 44, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Zhang, Z.; Qu, M.; Ding, S.; Zheng, L. Genomic organization, promoter characterization and expression analysis of the leukocyte cell-derived chemotaxin-2 gene in Epinephelus akaraa. Fish Shellfish Immunol. 2012, 32, 1041–1050. [Google Scholar] [CrossRef]
- Shan, S.J.; Liu, D.Z.; Wang, L.; Zhu, Y.Y.; Zhang, F.M.; Li, T.; An, L.G.; Yang, G.W. Identification and expression analysis of irak1 gene in common carp Cyprinus carpio L.: Indications for a role of antibacterial and antiviral immunity. J. Fish Biol. 2015, 87, 241–255. [Google Scholar] [CrossRef]
- Scheepers, A.; Schmidt, S.; Manolescu, A.; Cheeseman, C.I.; Bell, A.; Zahn, C.; Joost, H.G.; Schürmann, A. Characterization of the human SLC2A11 (GLUT11) gene: Alternative promoter usage, function, expression, and subcellular distribution of three isoforms, and lack of mouse orthologue. Mol. Membr. Biol. 2005, 22, 339–351. [Google Scholar] [CrossRef]
- Tseng, Y.C.; Hwang, P.P. Some insights into energy metabolism for osmoregulation in fish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2008, 148, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Kamalam, B.S.; Medale, F.; Panserat, S. Utilisation of dietary carbohydrates in farmed fishes: New insights on influencing factors, biological limitations and future strategies. Aquaculture 2017, 467, 3–27. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Microvillus Length (nm) | Microvillus Diameter (nm) | Abundance of Microvilli (µm−2) | |||||||
| Mean ± SE | Min | Max | Mean ± SE | Min | Max | Mean ± SE | Min | Max | |
| Germ-free | 2021.40 ± 31.62 | 1038.08 | 3183.59 | 107.01 ± 0.89 | 81.77 | 148.49 | 42.76 ± 1.03 | 25.39 | 68.43 |
| Conventional | 1703.04 ± 11.18 | 1211.07 | 2360.96 | 99.10 ± 0.53 | 77.51 | 123.92 | 54.14 ± 1.05 | 35.56 | 69.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vestrum, R.I.; Forberg, T.; Luef, B.; Bakke, I.; Winge, P.; Olsen, Y.; Vadstein, O. Commensal and Opportunistic Bacteria Present in the Microbiota in Atlantic Cod (Gadus morhua) Larvae Differentially Alter the Hosts’ Innate Immune Responses. Microorganisms 2022, 10, 24. https://doi.org/10.3390/microorganisms10010024
Vestrum RI, Forberg T, Luef B, Bakke I, Winge P, Olsen Y, Vadstein O. Commensal and Opportunistic Bacteria Present in the Microbiota in Atlantic Cod (Gadus morhua) Larvae Differentially Alter the Hosts’ Innate Immune Responses. Microorganisms. 2022; 10(1):24. https://doi.org/10.3390/microorganisms10010024
Chicago/Turabian StyleVestrum, Ragnhild Inderberg, Torunn Forberg, Birgit Luef, Ingrid Bakke, Per Winge, Yngvar Olsen, and Olav Vadstein. 2022. "Commensal and Opportunistic Bacteria Present in the Microbiota in Atlantic Cod (Gadus morhua) Larvae Differentially Alter the Hosts’ Innate Immune Responses" Microorganisms 10, no. 1: 24. https://doi.org/10.3390/microorganisms10010024
APA StyleVestrum, R. I., Forberg, T., Luef, B., Bakke, I., Winge, P., Olsen, Y., & Vadstein, O. (2022). Commensal and Opportunistic Bacteria Present in the Microbiota in Atlantic Cod (Gadus morhua) Larvae Differentially Alter the Hosts’ Innate Immune Responses. Microorganisms, 10(1), 24. https://doi.org/10.3390/microorganisms10010024